
CAM-Xenograft Model Provides Preclinical Evidence for the Applicability of [68Ga]Ga-Pentixafor in CRC Imaging
 , , , , ,
, , , , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Radiolabelling
2.2. Cell Culture
2.3. In Vitro Cell Internalisation Experiments
2.4. In Ovo Assay and Tumour Cell Inoculation
2.5. PET/MR Imaging
2.5.1. Image Acquisition
2.5.2. Image Post-Processing and Data Analysis
2.6. Histopathological Evaluation of the In Ovo CAM Xenografts
2.7. Statistical Analyses
3. Results
3.1. Radiolabelling
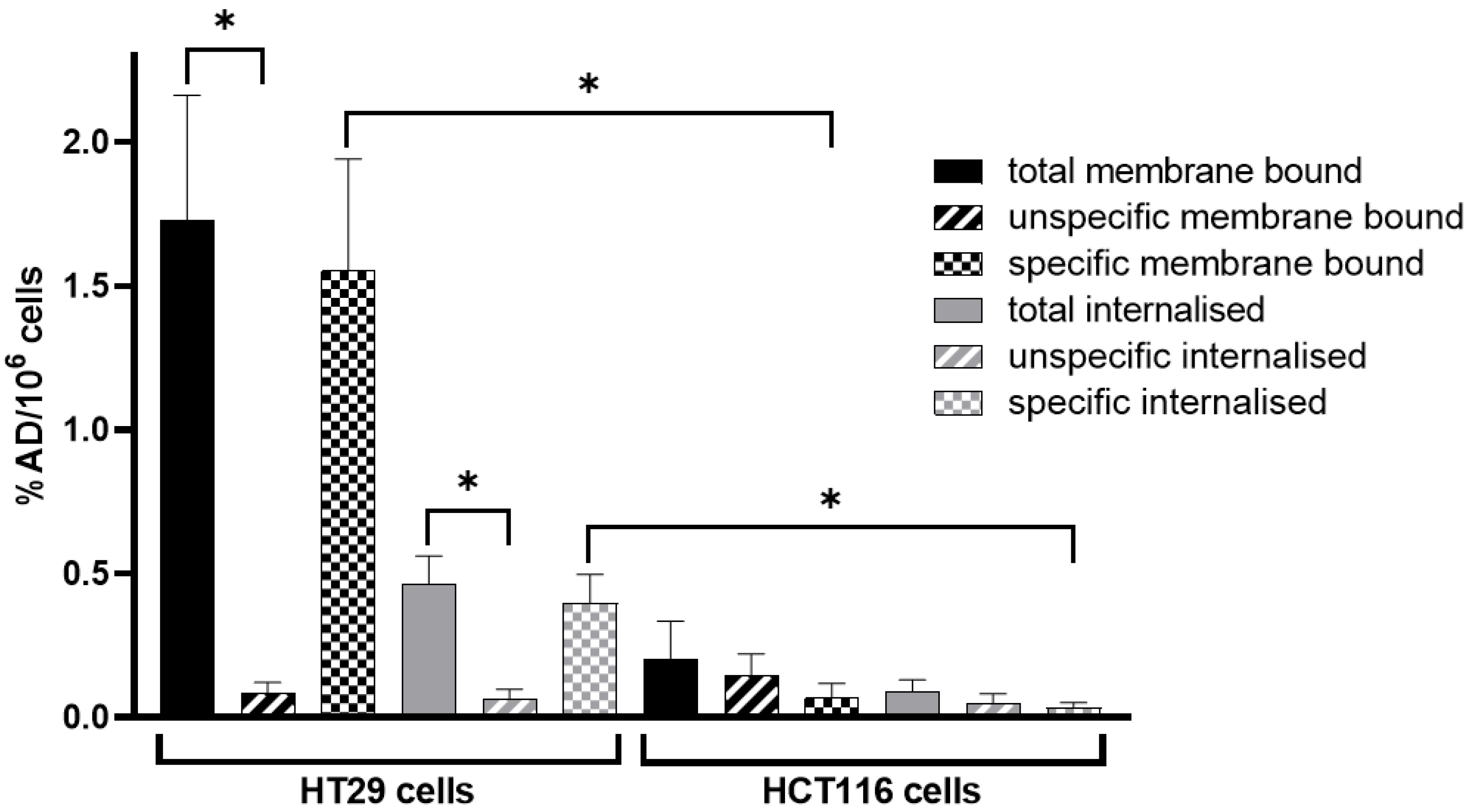
3.2. In Vitro Cell Internalisation Experiments
3.3. Multimodal Imaging
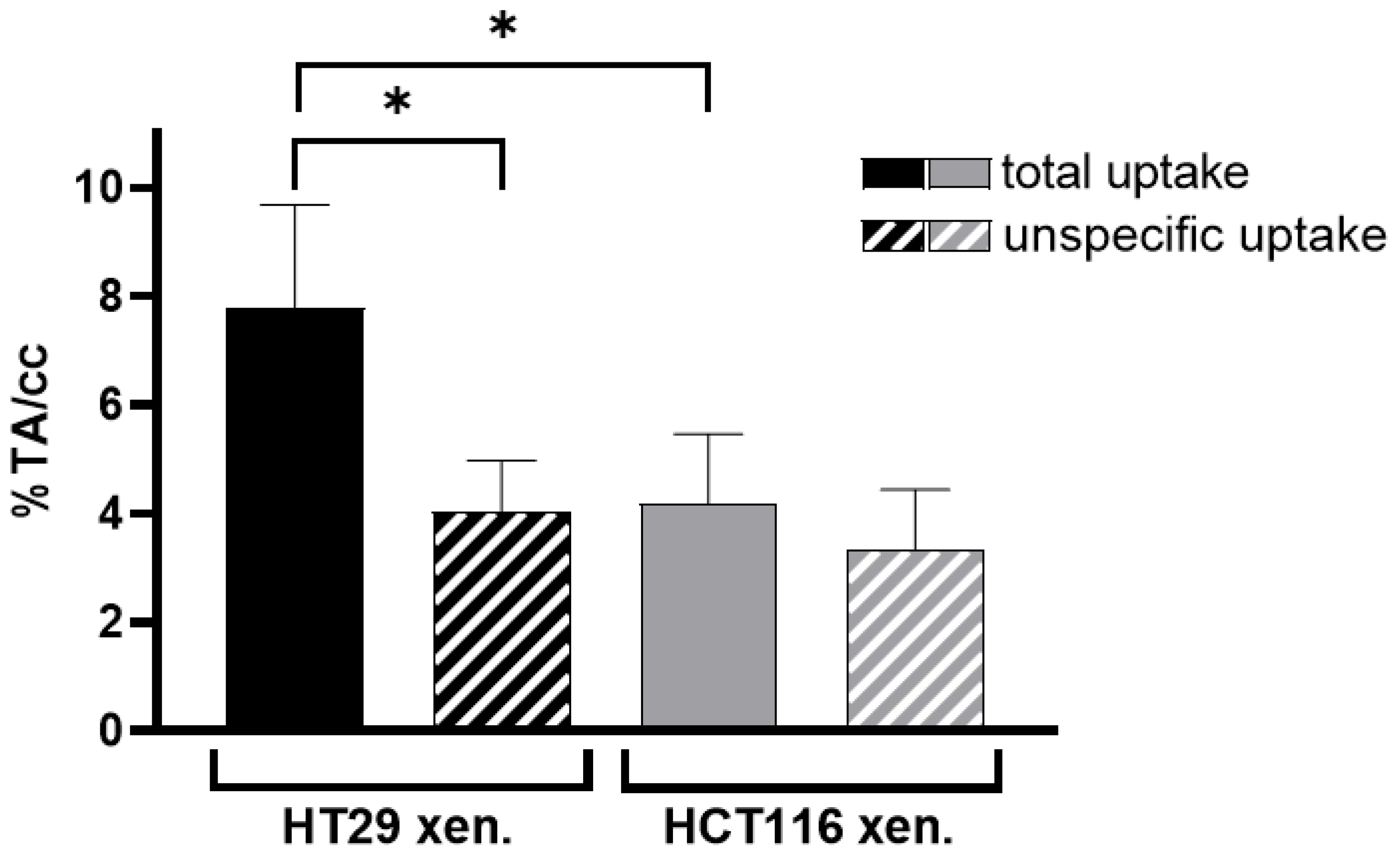
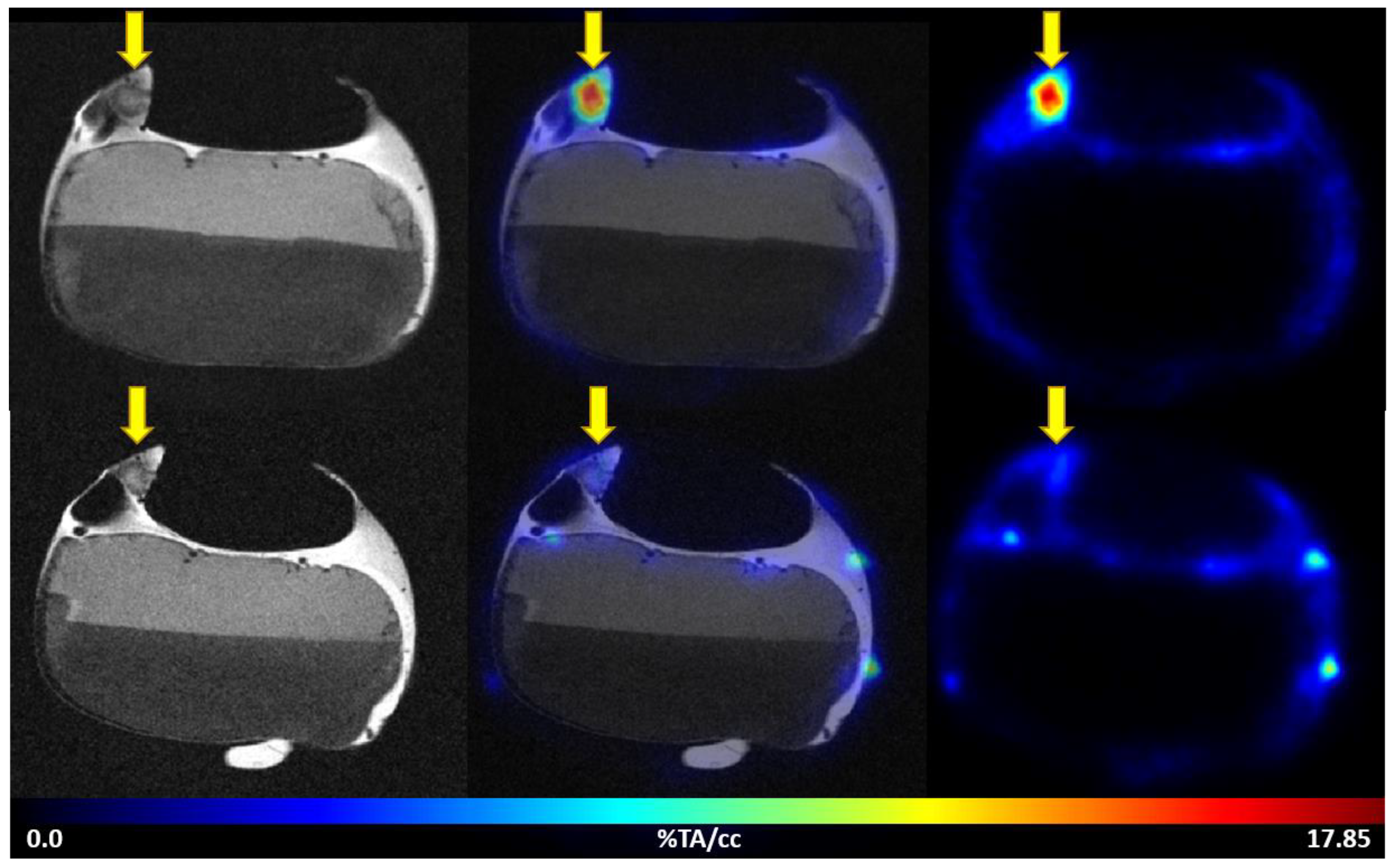
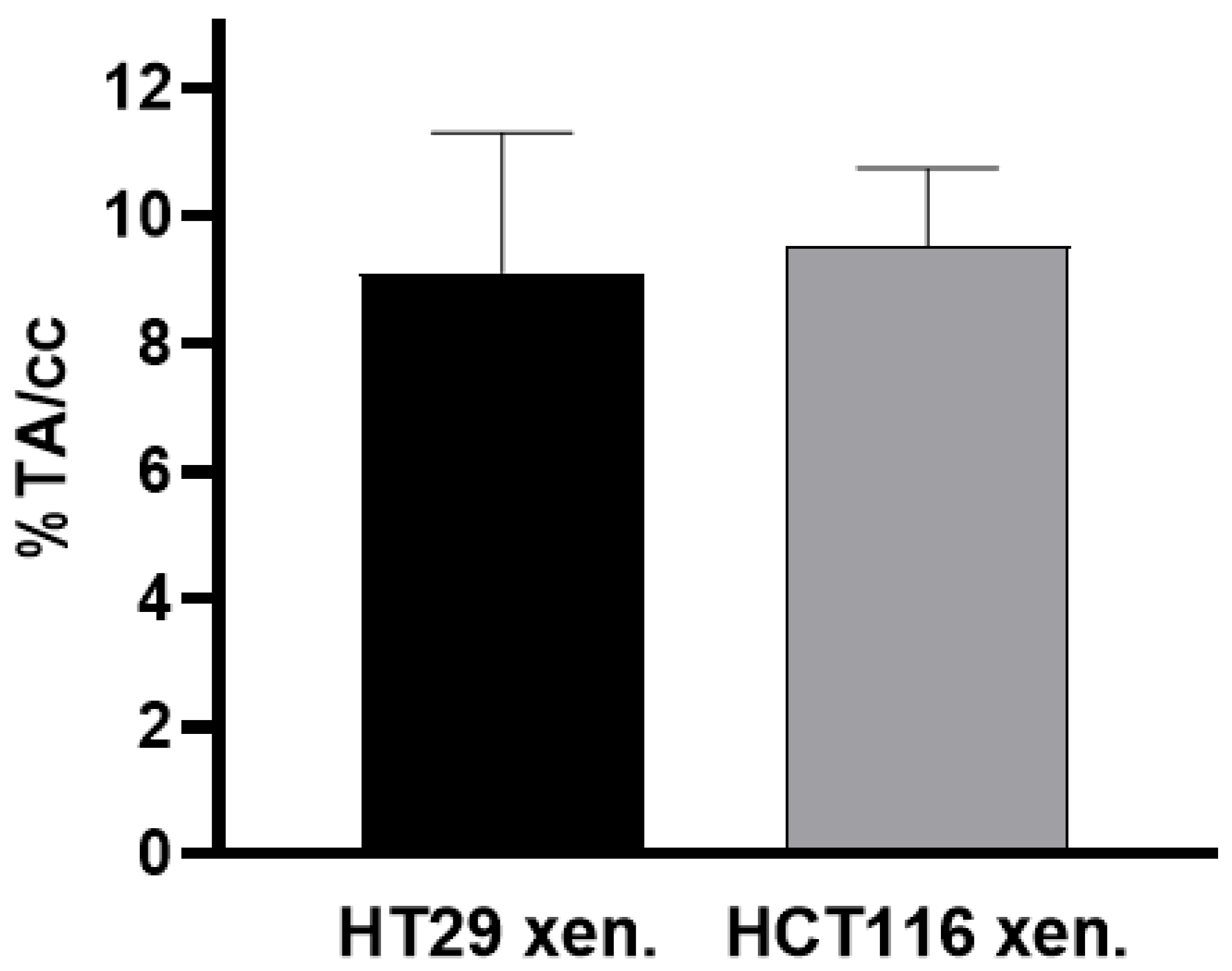
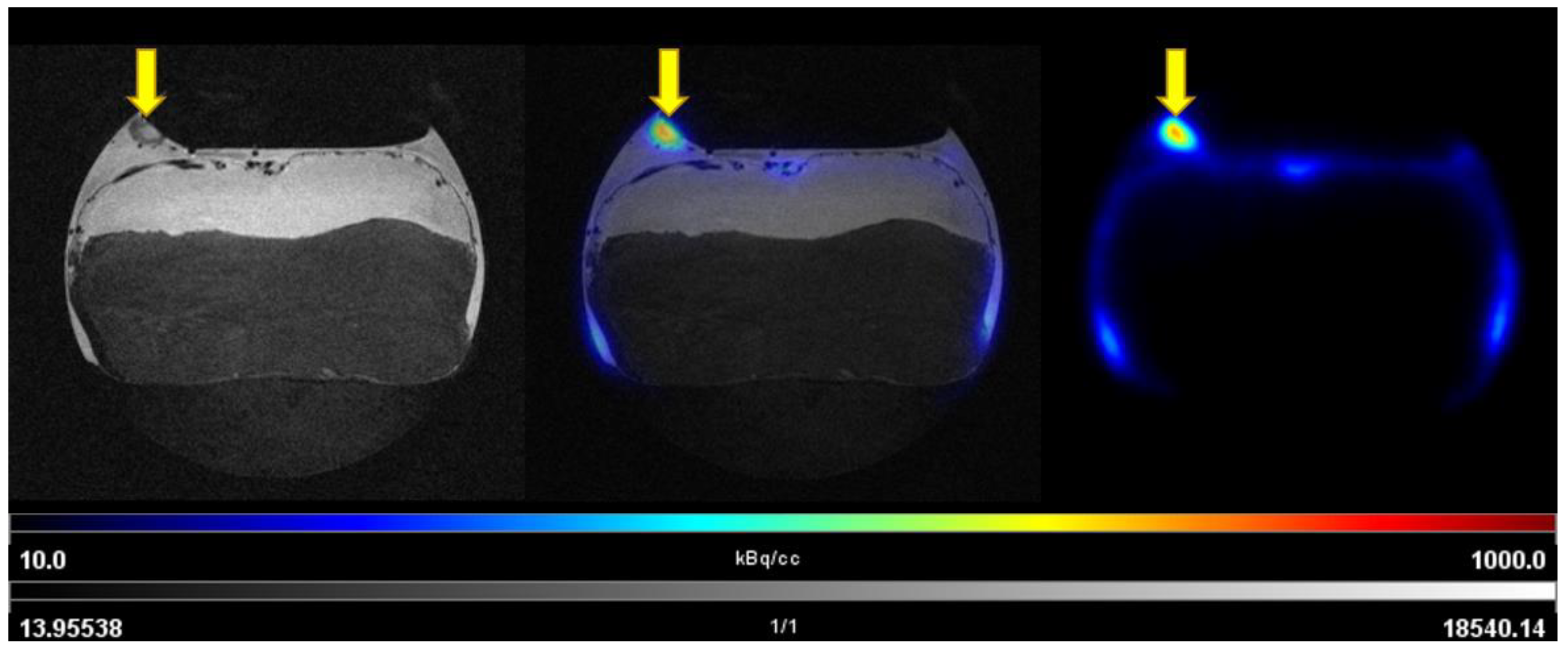
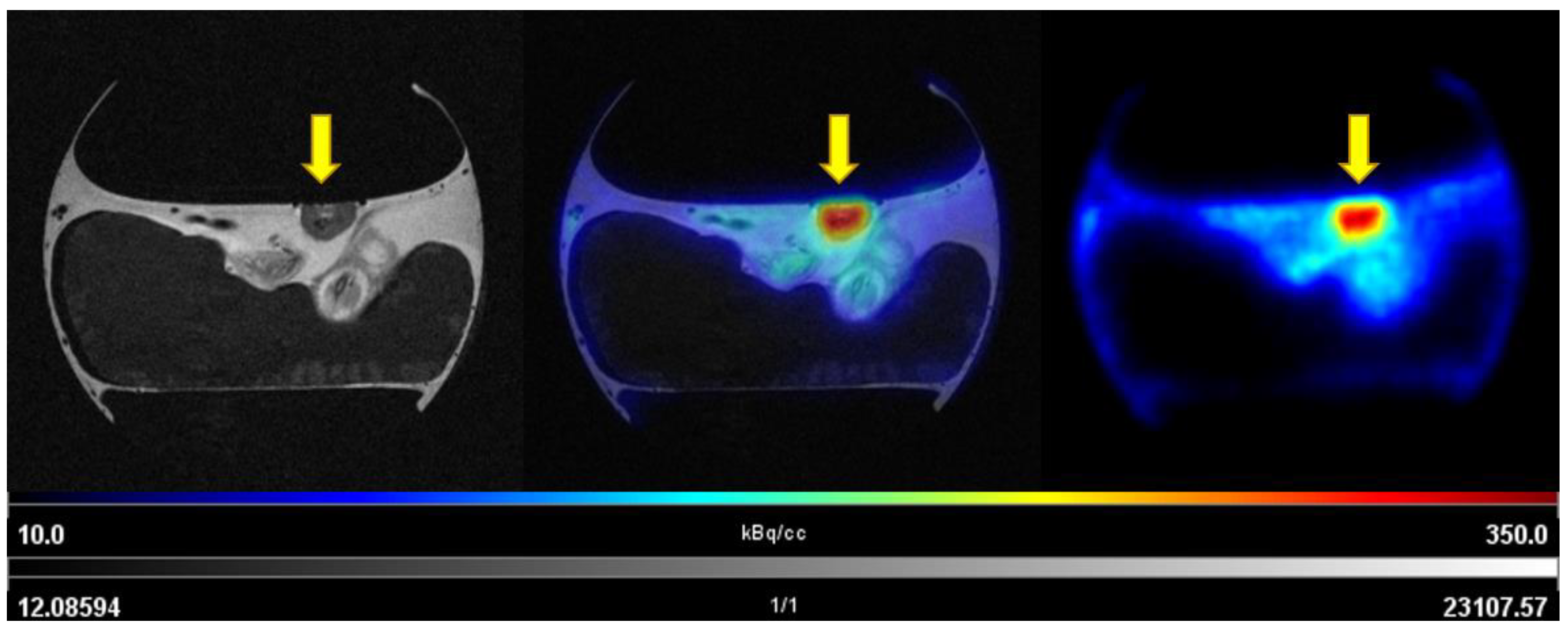
3.3.1. PET/MRI Measurements Using [68Ga]Ga-Pentixafor
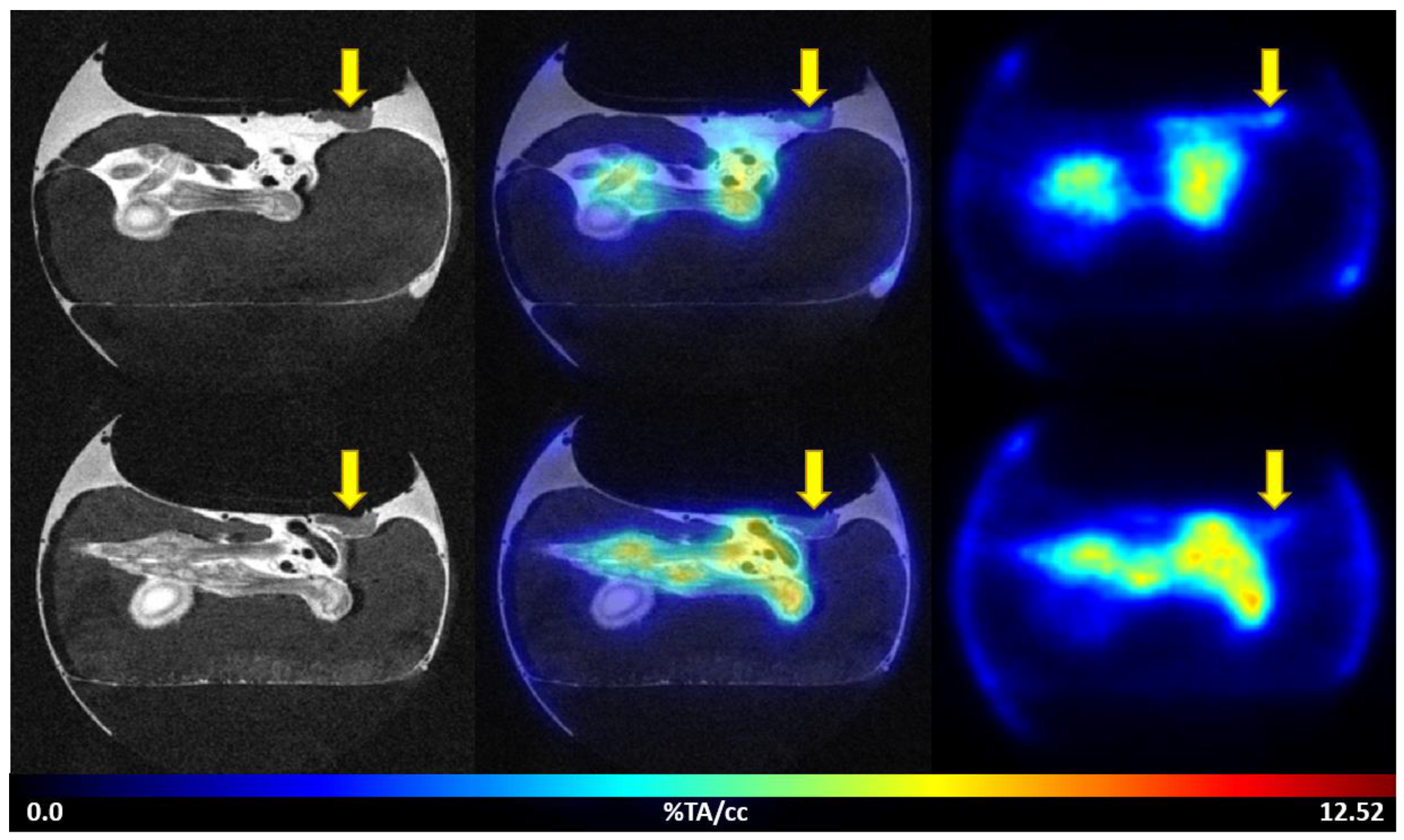
3.3.2. PET/MRI Measurements Using 2-[18F]FDG
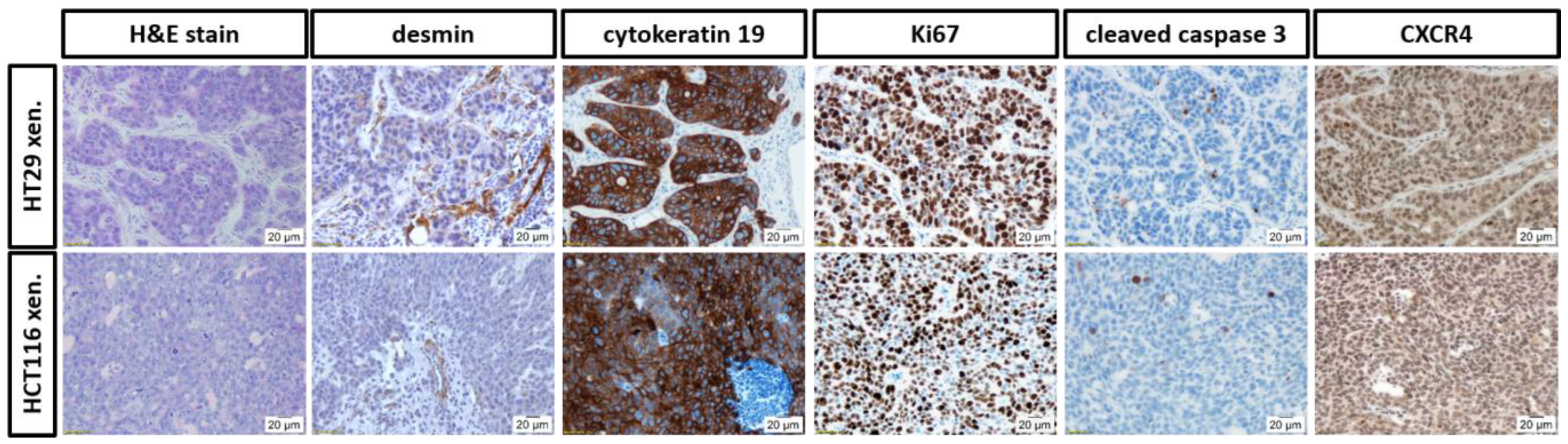
3.4. Histopathological Evaluation of the In Ovo CAM-Xenografts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Cancer Observatory: Cancer Today. Available online: https://gco.iarc.fr/today/online-analysis-pie (accessed on 21 August 2022).
- Surveillance, Epidemiology, and End Results Program. Cancer Statistics Explorer Network. SEER*Explorer. Available online: https://seer.cancer.gov/statistics-network/explorer (accessed on 21 August 2022).
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Przegląd Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Takeuchi, H.; Lam, S.T.; Turner, R.R.; Wang, H.-J.; Kuo, C.; Foshag, L.; Bilchik, A.J.; Hoon, D.S. Chemokine receptor CXCR4 expression in colorectal cancer patients increases the risk for recurrence and for poor survival. J. Clin. Oncol. 2005, 23, 2744–2753. [Google Scholar] [CrossRef] [PubMed]
- Schimanski, C.C.; Schwald, S.; Simiantonaki, N.; Jayasinghe, C.; Gönner, U.; Wilsberg, V.; Junginger, T.; Berger, M.R.; Galle, P.R.; Moehler, M. Effect of chemokine receptors CXCR4 and CCR7 on the metastatic behavior of human colorectal cancer. Clin. Cancer Res. 2005, 11, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.; Homey, B.; Soto, H.; Ge, N.; Catron, D.; Buchanan, M.E.; McClanahan, T.; Murphy, E.; Yuan, W.; Wagner, S.N.; et al. Involvement of chemokine receptors in breast cancer metastasis. Nature 2001, 410, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Zeelenberg, I.S.; Ruuls-Van Stalle, L.; Roos, E. The chemokine receptor CXCR4 is required for outgrowth of colon carcinoma micrometastases. Cancer Res. 2003, 63, 3833–3839. [Google Scholar]
- Xu, C.; Zheng, L.; Li, D.; Chen, G.; Gu, J.; Chen, J.; Yao, Q. CXCR4 overexpression is correlated with poor prognosis in colorectal cancer. Life Sci. 2018, 208, 333–340. [Google Scholar] [CrossRef]
- Ottaiano, A.; Santorsola, M.; Del Prete, P.; Perri, F.; Scala, S.; Caraglia, M.; Nasti, G. Prognostic significance of CXCR4 in colorectal cancer: An updated meta-analysis and critical appraisal. Cancers 2021, 13, 3284. [Google Scholar] [CrossRef]
- Vega, P.; Valentín, F.; Cubiella, J. Colorectal cancer diagnosis: Pitfalls and opportunities. World J. Gastrointest. Oncol. 2015, 7, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A review of colorectal cancer in terms of epidemiology, risk factors, development, symptoms and diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef]
- Lim, M.; Kim, C.-J.; Sunkara, V.; Kim, M.-H.; Cho, Y.-K. Liquid biopsy in lung cancer: Clinical applications of circulating biomarkers (CTCs and ctDNA). Micromachines 2018, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Lucignani, G. Imaging biomarkers: From research to patient care—A shift in view. Eur. J. Nucl. Med. Mol. Imaging 2007, 34, 1693–1697. [Google Scholar] [CrossRef] [PubMed]
- Society of Nuclear Medicine & Molecular Imaging. Fact Sheet: Molecular Imaging and Colorectal Cancer. Available online: https://www.snmmi.org/AboutSNMMI/Content.aspx?ItemNumber=5658# (accessed on 23 August 2022).
- Balber, T.; Tran, L.; Benčurová, K.; Raitanen, J.; Egger, G.; Mitterhauser, M. Experimental nuclear medicine meets tumour biology. Pharmaceuticals 2022, 15, 227. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; He, X.; Jin, C.; He, X.; Wu, S.; Zhou, R.; Zhang, X.; Zhang, K.; Gu, W.; Wang, J.; et al. Transpathology: Molecular imaging-based pathology. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 2338–2350. [Google Scholar] [CrossRef] [PubMed]
- Maffione, A.M.; Rubello, D.; Caroli, P.; Colletti, P.M.; Matteucci, F. Is it time to introduce PET/CT in colon cancer guidelines? Clin. Nucl. Med. 2020, 45, 525–530. [Google Scholar] [CrossRef]
- Ametamey, S.M.; Honer, M.; Schubiger, P.A. Molecular imaging with PET. Chem. Rev. 2008, 108, 1501–1516. [Google Scholar] [CrossRef]
- Ido, T.; Wan, C.N.; Fowler, J.S.; Wolf, A.P. Fluorination with F2. A convenient synthesis of 2-deoxy-2-fluoro-D-glucose. J. Org. Chem. 1977, 42, 2341–2342. [Google Scholar] [CrossRef]
- Wadsak, W.; Mitterhauser, M. Basics and principles of radiopharmaceuticals for PET/CT. Eur. J. Radiol. 2010, 73, 461–469. [Google Scholar] [CrossRef]
- Feng, H.; Wang, X.; Chen, J.; Cui, J.; Gao, T.; Gao, Y.; Zeng, W. Nuclear imaging of glucose metabolism: Beyond 18F-FDG. Contrast Media Mol. Imaging 2019, 2019, 7954854. [Google Scholar] [CrossRef] [Green Version]
- Van Cutsem, E.; Verheul, H.M.; Flamen, P.; Rougier, P.; Beets-Tan, R.; Glynne-Jones, R.; Seufferlein, T. Imaging in colorectal cancer: Progress and challenges for the clinicians. Cancers 2016, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Demmer, O.; Gourni, E.; Schumacher, U.; Kessler, H.; Wester, H.J. PET imaging of CXCR4 receptors in cancer by a new optimized ligand. ChemMedChem 2011, 6, 1789–1791. [Google Scholar] [CrossRef] [Green Version]
- Gourni, E.; Demmer, O.; Schottelius, M.; D’Alessandria, C.; Schulz, S.; Dijkgraaf, I.; Schumacher, U.; Schwaiger, M.; Kessler, H.; Wester, H.J. PET of CXCR4 expression by a 68Ga-labeled highly specific targeted contrast agent. J. Nucl. Med. 2011, 52, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Wester, H.J.; Keller, U.; Schottelius, M.; Beer, A.; Philipp-Abbrederis, K.; Hoffmann, F.; Šimeček, J.; Gerngross, C.; Lassmann, M.; Herrmann, K.; et al. Disclosing the CXCR4 expression in lymphoproliferative diseases by targeted molecular imaging. Theranostics 2015, 5, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Kawada, K.; Iwamoto, M.; Akagami, M.; Hida, K.; Nakanishi, Y.; Kanda, K.; Kawada, M.; Seno, H.; Taketo, M.M.; et al. The role of CXCR3 and CXCR4 in colorectal cancer metastasis. Int. J. Cancer 2013, 132, 276–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogh, J.; Trempe, G. New human tumour cell lines. In Human Tumour Cells In Vitro, 1st ed.; Fogh, J., Ed.; Springer: Boston, MA, USA, 1975; Volume 1, pp. 115–159. [Google Scholar] [CrossRef]
- Brattain, M.G.; Fine, W.D.; Khaled, F.M.; Thompson, J.; Brattain, D.E. Heterogeneity of malignant cells from a human colonic carcinoma. Cancer Res. 1981, 41, 1751–1756. [Google Scholar] [CrossRef]
- Kleibeuker, E.A.; Ten Hooven, M.A.; Castricum, K.C.; Honeywell, R.; Griffioen, A.W.; Verheul, H.M.; Slotman, B.J.; Thijssen, V.L. Optimal treatment scheduling of ionizing radiation and sunitinib improves the antitumour activity and allows dose reduction. Cancer Med. 2015, 4, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Steinmann, S.; Kunze, P.; Hampel, C.; Eckstein, M.; Bertram Bramsen, J.; Muenzner, J.K.; Carlé, B.; Ndreshkjana, B.; Kemenes, S.; Gasparini, P.; et al. DAPK1 loss triggers tumour invasion in colorectal tumour cells. Cell Death Dis. 2019, 10, 895. [Google Scholar] [CrossRef] [Green Version]
- Kunze, P.; Kreiss, L.; Novosadová, V.; Roehe, A.V.; Steinmann, S.; Prochazka, J.; Geppert, C.I.; Hartmann, A.; Schürmann, S.; Friedrich, O.; et al. Multiphoton microscopy reveals DAPK1-dependent extracellular matrix remodeling in a chorioallantoic membrane (CAM) model. Cancers 2022, 14, 2364. [Google Scholar] [CrossRef]
- Lauzier, A.; Normandeau-Guimond, J.; Vaillancourt-Lavigueur, V.; Boivin, V.; Charbonneau, M.; Rivard, N.; Scott, M.S.; Dubois, C.M.; Jean, S. Colorectal cancer cells respond differentially to autophagy inhibition in vivo. Sci. Rep. 2019, 9, 11316. [Google Scholar] [CrossRef] [Green Version]
- Rous, P.; Murphy, J.B. Tumour implantations in the developing embryo. J. Am. Med. Assoc. 1911, LVI, 741–742. [Google Scholar]
- Ribatti, D. The chick embryo chorioallantoic membrane as a model for tumour biology. Exp. Cell Res. 2014, 328, 314–324. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane (CAM). A multifaceted experimental model. Mech. Dev. 2016, 141, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Koch, A.B.F.; Löffler, J.; Jelezko, F.; Lindén, M.; Li, H.; Abaei, A.; Zuo, Z.; Beer, A.J.; Rasche, V. In vivo PET/MRI imaging of the chorioallantoic membrane. Front. Phys. 2020, 8, 151. [Google Scholar] [CrossRef]
- Würbach, L.; Heidrich, A.; Opfermann, T.; Gebhardt, P.; Saluz, H.P. Insights into bone metabolism of avian embryos in ovo via 3D and 4D 18F-fluoride positron emission tomography. Mol. Imaging Biol. MIB Off. Publ. Acad. Mol. Imaging 2012, 14, 688–698. [Google Scholar] [CrossRef]
- Gebhardt, P.; Würbach, L.; Heidrich, A.; Heinrich, L.; Walther, M.; Opfermann, T.; Sørensen, B.; Saluz, P. Dynamic behaviour of selected PET tracers in embryonated chicken eggs. Rev. Española De Med. Nucl. E Imagen Mol. 2013, 32, 371–377. [Google Scholar] [CrossRef]
- Warnock, G.; Turtoi, A.; Blomme, A.; Bretin, F.; Bahri, M.A.; Lemaire, C.; Libert, L.C.; Seret, A.E.; Luxen, A.; Castronovo, V.; et al. In vivo PET/CT in a human glioblastoma chicken chorioallantoic membrane model: A new tool for oncology and radiotracer development. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2013, 54, 1782–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haller, S.; Ametamey, S.M.; Schibli, R.; Müller, C. Investigation of the chick embryo as a potential alternative to the mouse for evaluation of radiopharmaceuticals. Nucl. Med. Biol. 2015, 42, 226–233. [Google Scholar] [CrossRef]
- Zlatopolskiy, B.D.; Zischler, J.; Schäfer, D.; Urusova, E.A.; Guliyev, M.; Bannykh, O.; Endepols, H.; Neumaier, B. Discovery of 7-[18F]fluorotryptophan as a novel positron emission tomography (PET) probe for the visualization of tryptophan metabolism in vivo. J. Med. Chem. 2018, 61, 189–206. [Google Scholar] [CrossRef]
- Steinemann, G.; Dittmer, A.; Schmidt, J.; Josuttis, D.; Fähling, M.; Biersack, B.; Beindorff, N.; Koziolek, E.J.; Schobert, R.; Brenner, W.; et al. Antitumour and antiangiogenic activity of the novel chimeric inhibitor animacroxam in testicular germ cell cancer. Mol. Oncol. 2019, 13, 2679–2696. [Google Scholar] [CrossRef] [Green Version]
- Winter, G.; Koch, A.B.F.; Löffler, J.; Lindén, M.; Solbach, C.; Abaei, A.; Li, H.; Glatting, G.; Beer, A.J.; Rasche, V. Multi-modal PET and MR imaging in the hen’s egg test-chorioallantoic membrane (HET-CAM) model for initial in vivo testing of target-specific radioligands. Cancers 2020, 12, 1248. [Google Scholar] [CrossRef]
- Briest, F.; Koziolek, E.J.; Albrecht, J.; Schmidt, F.; Bernsen, M.R.; Haeck, J.; Kühl, A.A.; Sedding, D.L.; Hartung, T.; Exner, S.; et al. Does the proteasome inhibitor bortezomib sensitize to DNA-damaging therapy in gastroenteropancreatic neuroendocrine neoplasms?—A preclinical assessment in vitro and in vivo. Neoplasia 2021, 23, 80–98. [Google Scholar] [CrossRef]
- Löffler, J.; Hamp, C.; Scheidhauer, E.; Di Carlo, D.; Solbach, C.; Abaei, A.; Hao, L.; Glatting, G.; Beer, A.J.; Rasche, V.; et al. Comparison of quantification of target-specific accumulation of [(18)F]F-siPSMA-14 in the HET-CAM model and in mice using PET/MRI. Cancers 2021, 13, 4007. [Google Scholar] [CrossRef] [PubMed]
- Löffler, J.; Herrmann, H.; Scheidhauer, E.; Wirth, M.; Wasserloos, A.; Solbach, C.; Glatting, G.; Beer, A.J.; Rasche, V.; Winter, G. Blocking studies to evaluate receptor-specific radioligand binding in the CAM model by PET and MR imaging. Cancers 2022, 14, 3870. [Google Scholar] [CrossRef] [PubMed]
- Rosenbruch, M. The sensitivity of chicken embryos in incubated eggs. ALTEX-Altern. Anim. Exp. 1997, 14, 111–113. [Google Scholar]
- Weber, W.T.; Mausner, R. Migration patterns of avian embryonic bone marrow cells and their differentiation to functional T and B cells. In Avian Immunology, 1st ed.; Benedict, A.A., Ed.; Springer: Boston, MA, USA, 1977; Volume 88, pp. 47–59. [Google Scholar] [CrossRef]
- Janse, E.M.; Jeurissen, S.H. Ontogeny and function of two non-lymphoid cell populations in the chicken embryo. Immunobiology 1991, 182, 472–481. [Google Scholar] [CrossRef]
- Wörsdörfer, P.; Dalda, N.; Kern, A.; Krüger, S.; Wagner, N.; Kwok, C.K.; Henke, E.; Ergün, S. Generation of complex human organoid models including vascular networks by incorporation of mesodermal progenitor cells. Sci. Rep. 2019, 9, 15663. [Google Scholar] [CrossRef] [Green Version]
- Garreta, E.; Prado, P.; Tarantino, C.; Oria, R.; Fanlo, L.; Martí, E.; Zalvidea, D.; Trepat, X.; Roca-Cusachs, P.; Gavaldà-Navarro, A.; et al. Fine tuning the extracellular environment accelerates the derivation of kidney organoids from human pluripotent stem cells. Nat. Mater. 2019, 18, 397–405. [Google Scholar] [CrossRef]
- DeBord, L.C.; Pathak, R.R.; Villaneuva, M.; Liu, H.C.; Harrington, D.A.; Yu, W.; Lewis, M.T.; Sikora, A.G. The chick chorioallantoic membrane (CAM) as a versatile patient-derived xenograft (PDX) platform for precision medicine and preclinical research. Am. J. Cancer Res. 2018, 8, 1642–1660. [Google Scholar]
- Bundesgesetz über Versuche an lebenden Tieren (Tierversuchsgesetz 2012—TVG 2012) (§ 1 Abs.1, § 2 Z 1 lit. b) [Austrian Animal Experimentation Law 2012]. Available online: https://www.ris.bka.gv.at/GeltendeFassung/Bundesnormen/20008142/TVG%202012%2c%20Fassung%20vom%2007.11.2022.pdf (accessed on 25 September 2022).
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique, 1st ed.; Methuen & Co.: London, UK, 1959. [Google Scholar] [CrossRef]
- Spreckelmeyer, S.; Schulze, O.; Brenner, W. Fully-automated production of [68Ga]Ga-PentixaFor on the module Modular Lab-PharmTracer. EJNMMI Radiopharm. Chem. 2020, 5, 8. [Google Scholar] [CrossRef]
- Watts, A.; Chutani, S.; Arora, D.; Madivanane, V.; Thakur, S.; Kamboj, M.; Singh, B. Automated radiosynthesis, quality control, and biodistribution of Ga-68 Pentixafor: First Indian experience. Indian J. Nucl. Med. 2021, 36, 237–244. [Google Scholar] [CrossRef]
- Tatsi, A.; Maina, T.; Cescato, R.; Waser, B.; Krenning, E.P.; de Jong, M.; Cordopatis, P.; Reubi, J.C.; Nock, B.A. [111In-DOTA]Somatostatin-14 analogs as potential pansomatostatin-like radiotracers—First results of a preclinical study. EJNMMI Res. 2012, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Z.; Syrovets, T.; Wu, Y.; Hafner, S.; Vernikouskaya, I.; Liu, W.; Ma, G.; Weil, T.; Simmet, T.; Rasche, V. The CAM cancer xenograft as a model for initial evaluation of MR labelled compounds. Sci. Rep. 2017, 7, 46690. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, M.; Becker, J.; Eberth, S.; Kube, D.; Wilting, J. The chick chorioallantoic membrane as an in vivo xenograft model for Burkitt lymphoma. BMC Cancer 2014, 14, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz, P.; Schenker, A.; Sähr, H.; Lehner, B.; Fellenberg, J. Optimization of the chicken chorioallantoic membrane assay as reliable in vivo model for the analysis of osteosarcoma. PLoS ONE 2019, 14, e0215312. [Google Scholar] [CrossRef] [PubMed]
- Burke, W.H.; Sharp, P.J. Sex differences in body weight of chicken embryos. Poult. Sci. 1989, 68, 805–810. [Google Scholar] [CrossRef]
- Heidrich, A.; Würbach, L.; Opfermann, T.; Saluz, H.P. Motion-artifact-free in vivo imaging utilizing narcotized avian embryos in ovo. Mol. Imaging Biol. 2001, 13, 208–214. [Google Scholar] [CrossRef]
- Yusuf, F.; Rehimi, R.; Dai, F.; Brand-Saperi, B. Expression of chemokine receptor CXCR4 during chick embryo development. Anat. Embryol. 2005, 210, 35–41. [Google Scholar] [CrossRef]
- Liang, T.S.; Hartt, J.K.; Lu, S.; Martins-Green, M.; Gao, J.L.; Murphy, P.M. Cloning, mRNA distribution, and functional expression of an avian counterpart of the chemokine receptor/HIV coreceptor CXCR4. J. Leukoc. Biol. 2001, 69, 297–305. [Google Scholar] [CrossRef]
- Osl, T.; Schmidt, A.; Schwaiger, M.; Schottelius, M.; Wester, H.J. A new class of PentixaFor- and PentixaTher-based theranostic agents with enhanced CXCR4-targeting efficiency. Theranostics 2020, 10, 8264–8280. [Google Scholar] [CrossRef]
- Demmer, O.; Dijkgraaf, I.; Schumacher, U.; Marinelli, L.; Cosconati, S.; Gourni, E.; Wester, H.J.; Kessler, H. Design, synthesis, and functionalization of dimeric peptides targeting chemokine receptor CXCR4. J. Med. Chem. 2011, 54, 7648–7662. [Google Scholar] [CrossRef]
- Ensembl. Gene: CXCR4. Available online: https://www.ensembl.org/Homo_sapiens/Gene/Summary?g=ENSG00000121966 (accessed on 29 August 2022).
- COSMIC. Catalogue of Somatic Mutations in Cancer. Sample Information: HT-29. Available online: https://cancer.sanger.ac.uk/cosmic/sample/overview?id=1998447 (accessed on 29 August 2022).
- COSMIC. Catalogue of Somatic Mutations in Cancer. Sample Information: HCT116. Available online: https://cancer.sanger.ac.uk/cosmic/sample/overview?id=2301978 (accessed on 29 August 2022).
- COSMIC. Catalogue of Somatic Mutations in Cancer. Genomic Mutation ID: COSV54011351. Available online: https://cancer.sanger.ac.uk/cosmic/mutation/overview?id=123752043 (accessed on 29 August 2022).
- COSMIC. Catalogue of Somatic Mutations in Cancer. Genomic Mutation ID: COSV54010069. Available online: https://cancer.sanger.ac.uk/cosmic/mutation/overview?id=123751801 (accessed on 29 August 2022).
- Bozkurt, E.; Atay, E.; Bilir, A.; Ertekin, A.; Buğra Koca, H.; Cem Sabaner, M. A novel model of early type 1 diabetes mellitus: The chick embryo air sack model. Saudi J. Biol. Sci. 2021, 28, 5538–5546. [Google Scholar] [CrossRef]
- Christensen, V.L.; Wineland, M.J.; Fasenko, G.M.; Donaldson, W.E. Egg storage effects on plasma glucose and supply and demand tissue glycogen concentrations of broiler embryos. Poult. Sci. 2001, 80, 1729–1735. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benčurová, K.; Friske, J.; Anderla, M.; Mayrhofer, M.; Wanek, T.; Nics, L.; Egger, G.; Helbich, T.H.; Hacker, M.; Haug, A.; et al. CAM-Xenograft Model Provides Preclinical Evidence for the Applicability of [68Ga]Ga-Pentixafor in CRC Imaging. Cancers 2022, 14, 5549. https://doi.org/10.3390/cancers14225549
Benčurová K, Friske J, Anderla M, Mayrhofer M, Wanek T, Nics L, Egger G, Helbich TH, Hacker M, Haug A, et al. CAM-Xenograft Model Provides Preclinical Evidence for the Applicability of [68Ga]Ga-Pentixafor in CRC Imaging. Cancers. 2022; 14(22):5549. https://doi.org/10.3390/cancers14225549
Chicago/Turabian StyleBenčurová, Katarína, Joachim Friske, Maximilian Anderla, Manuela Mayrhofer, Thomas Wanek, Lukas Nics, Gerda Egger, Thomas H. Helbich, Marcus Hacker, Alexander Haug, and et al. 2022. "CAM-Xenograft Model Provides Preclinical Evidence for the Applicability of [68Ga]Ga-Pentixafor in CRC Imaging" Cancers 14, no. 22: 5549. https://doi.org/10.3390/cancers14225549
APA StyleBenčurová, K., Friske, J., Anderla, M., Mayrhofer, M., Wanek, T., Nics, L., Egger, G., Helbich, T. H., Hacker, M., Haug, A., Mitterhauser, M., & Balber, T. (2022). CAM-Xenograft Model Provides Preclinical Evidence for the Applicability of [68Ga]Ga-Pentixafor in CRC Imaging. Cancers, 14(22), 5549. https://doi.org/10.3390/cancers14225549