
Pan-Cancer Analysis Identifies Tumor Cell Surface Targets for CAR-T Cell Therapies and Antibody Drug Conjugates
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
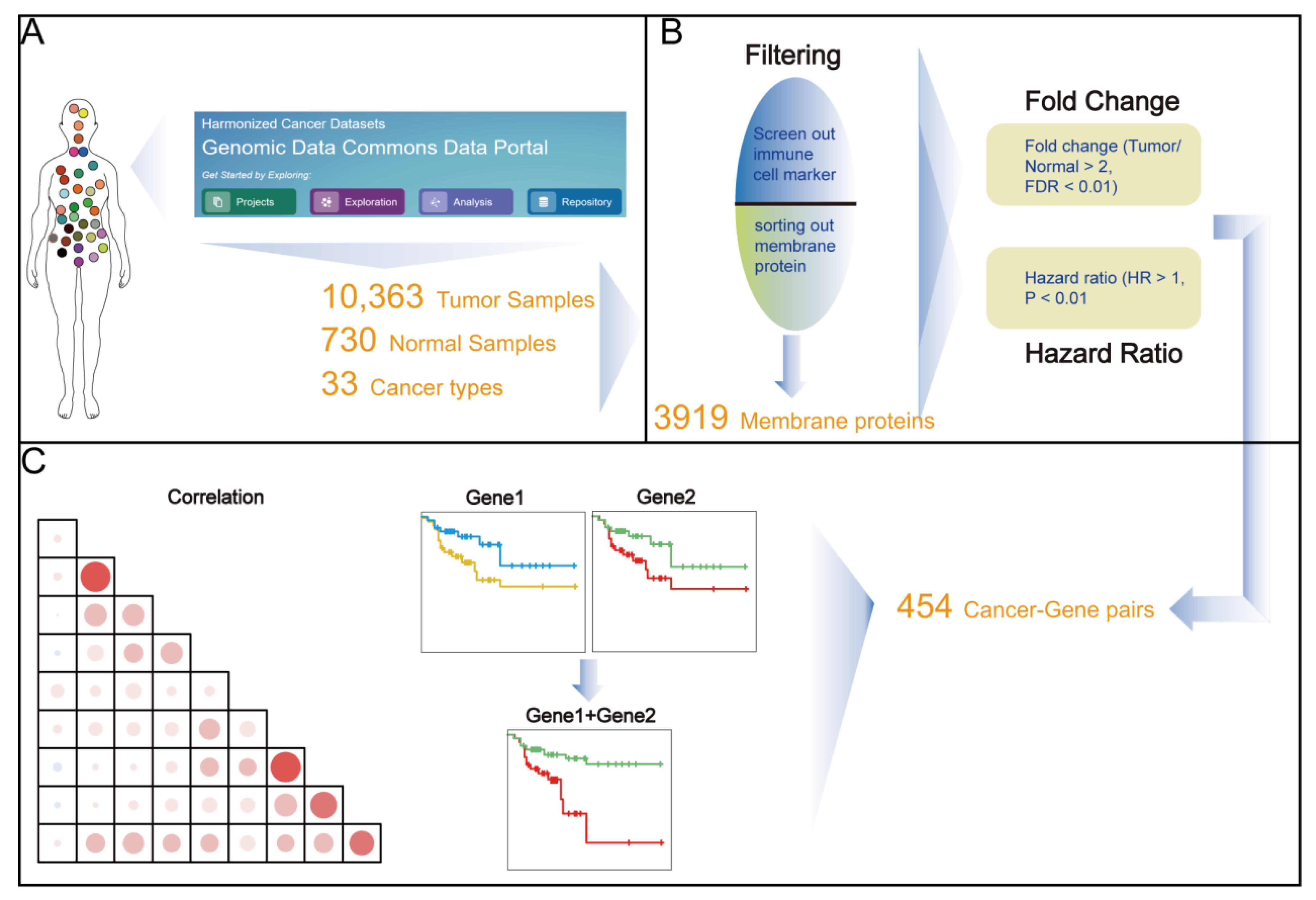
2. Materials and Methods
2.1. Data Source
2.1.1. TCGA
2.1.2. CPTAC (Clinical Proteomic Tumor Analysis Consortium)
2.2. Differential Expression Analysis
2.2.1. TCGA
2.2.2. CPTAC
2.3. Enrichment Analysis
2.4. Survival Analysis
2.4.1. Log-Rank Test
2.4.2. Multivariate/Univariate Cox Regression
2.5. Risk-Score System Establishment
2.6. Correlation Analysis
3. Results
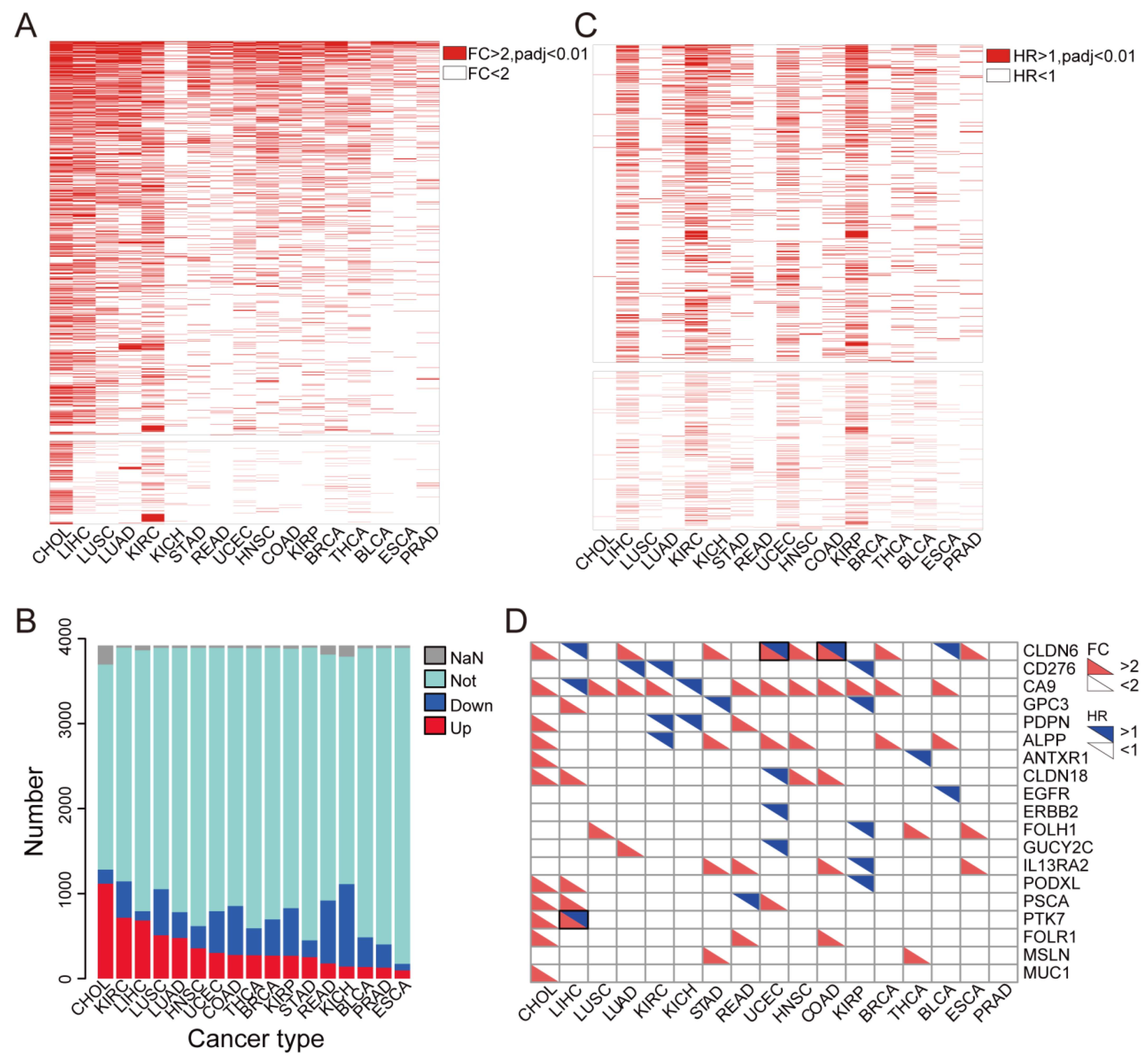
3.1. Identification of Up-Regulated Tumor Cell Membrane Proteins
3.2. Most Highly Expressed Membrane Protein-Coding Genes Could Serve as Risk Factors for Cancer Patients
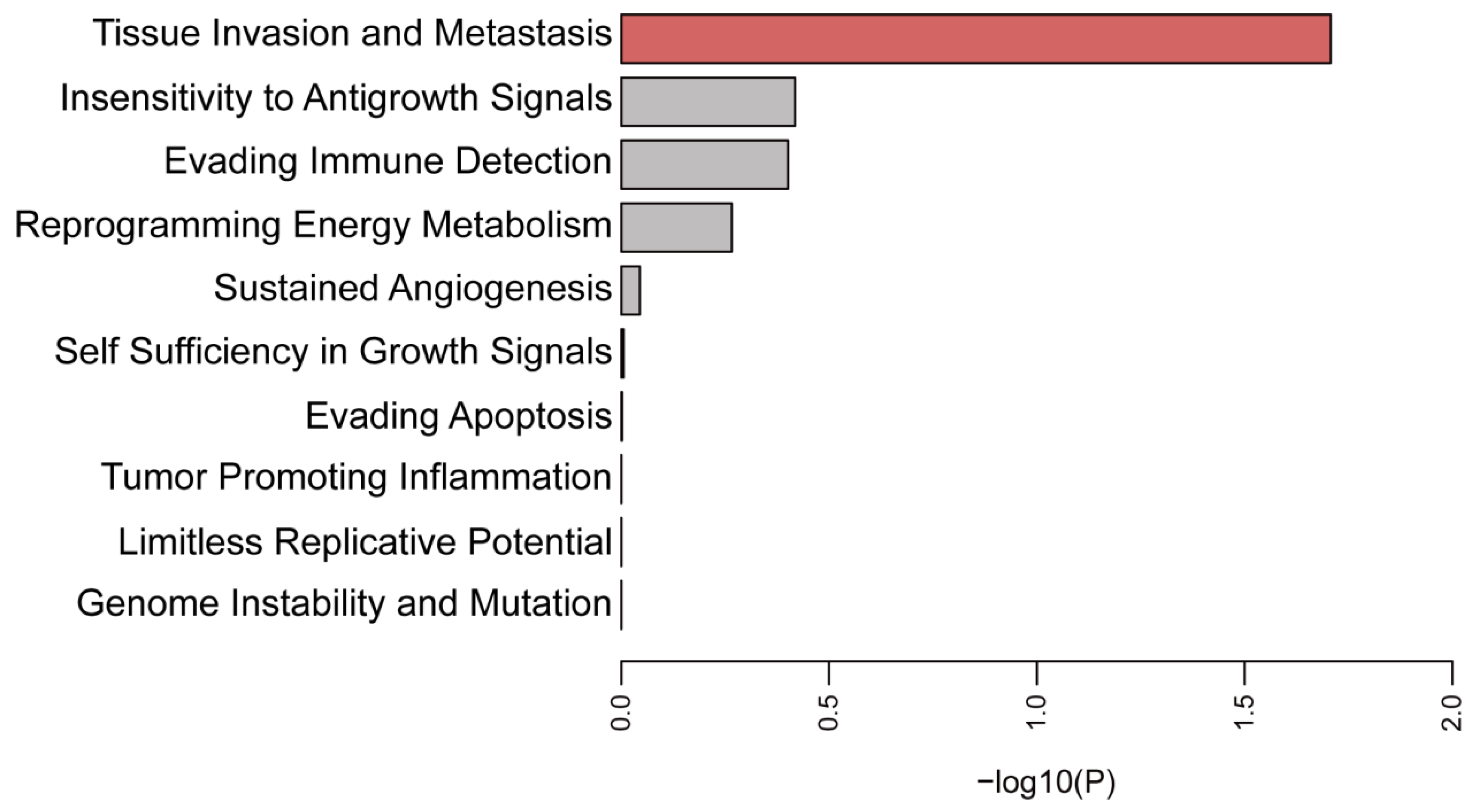
3.3. Function Enrichment Analysis of the Selected Membrane Proteins
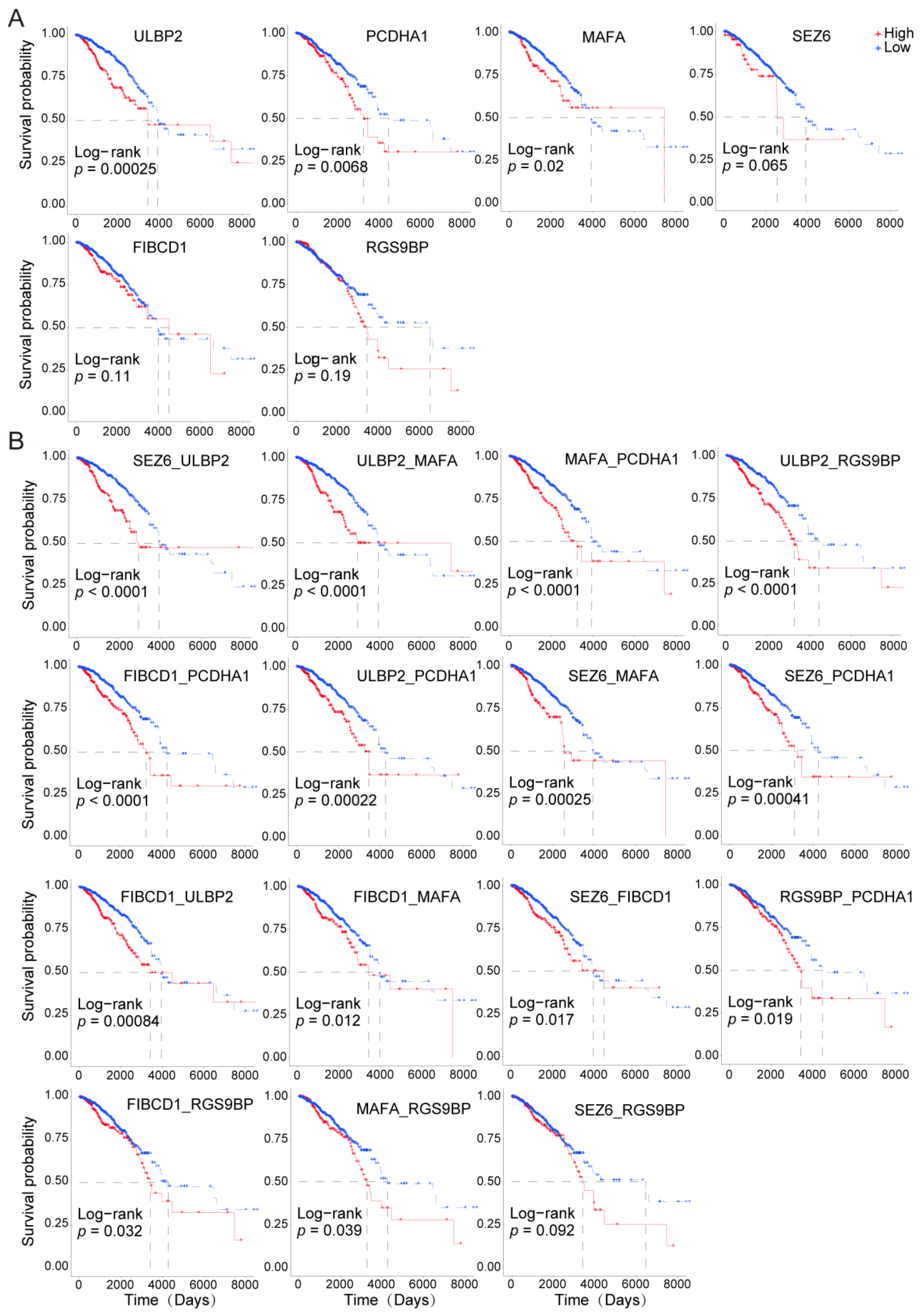
3.4. Paired Membrane Proteins Displayed More Precise Prognostic Value
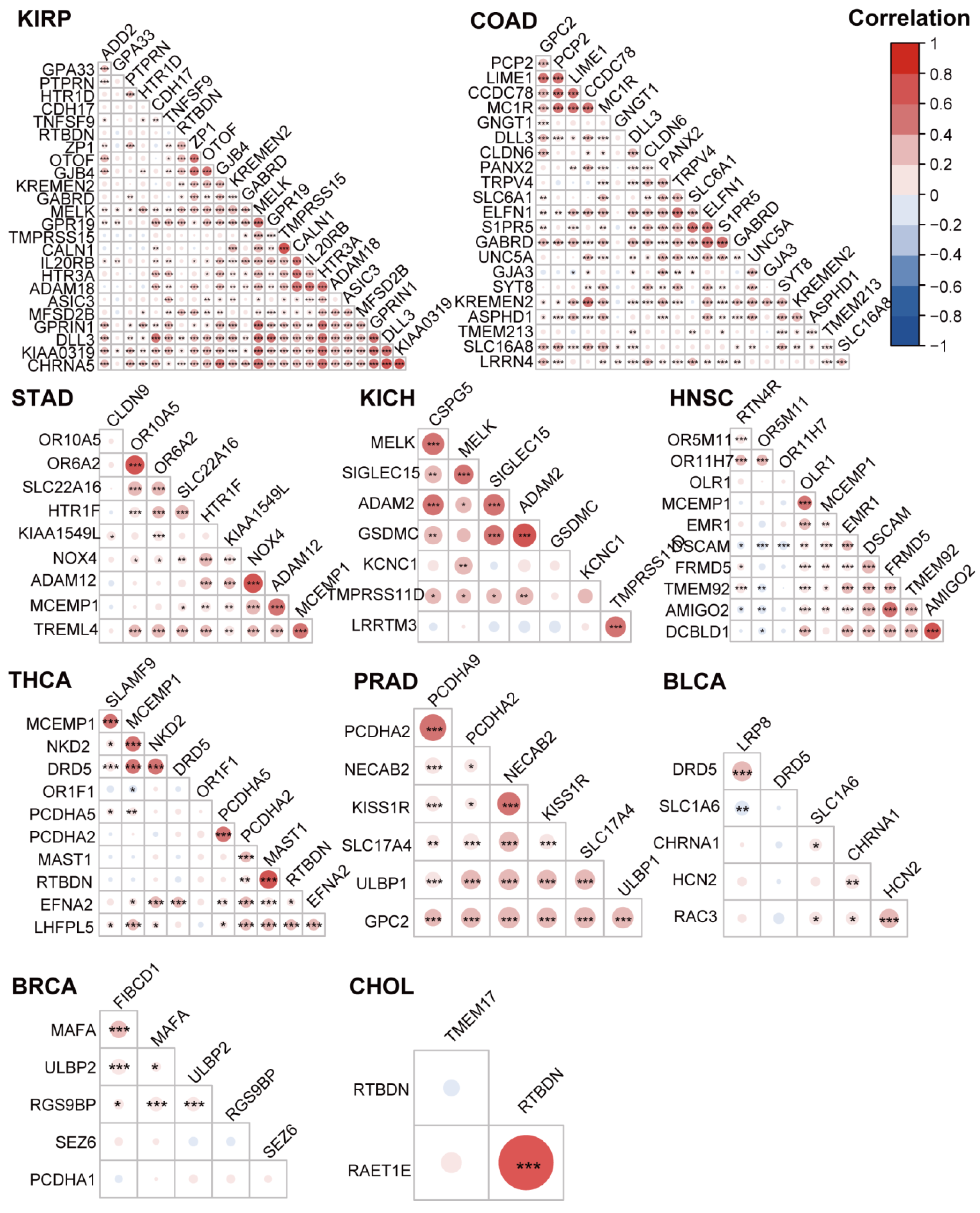
3.5. The Highly Expressed Cell Surface Proteins of the Tumor Tissues were Highly Correlated
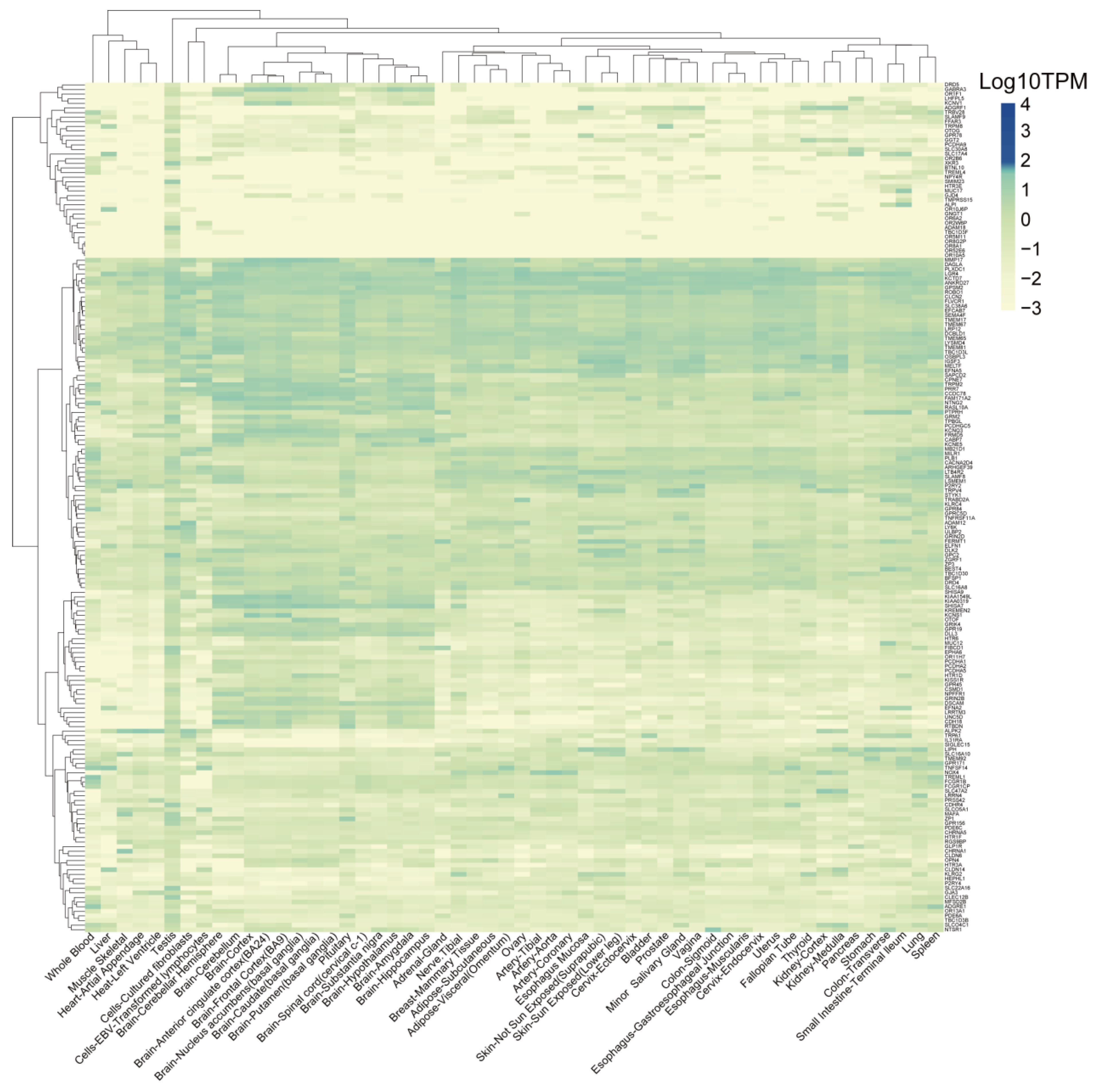
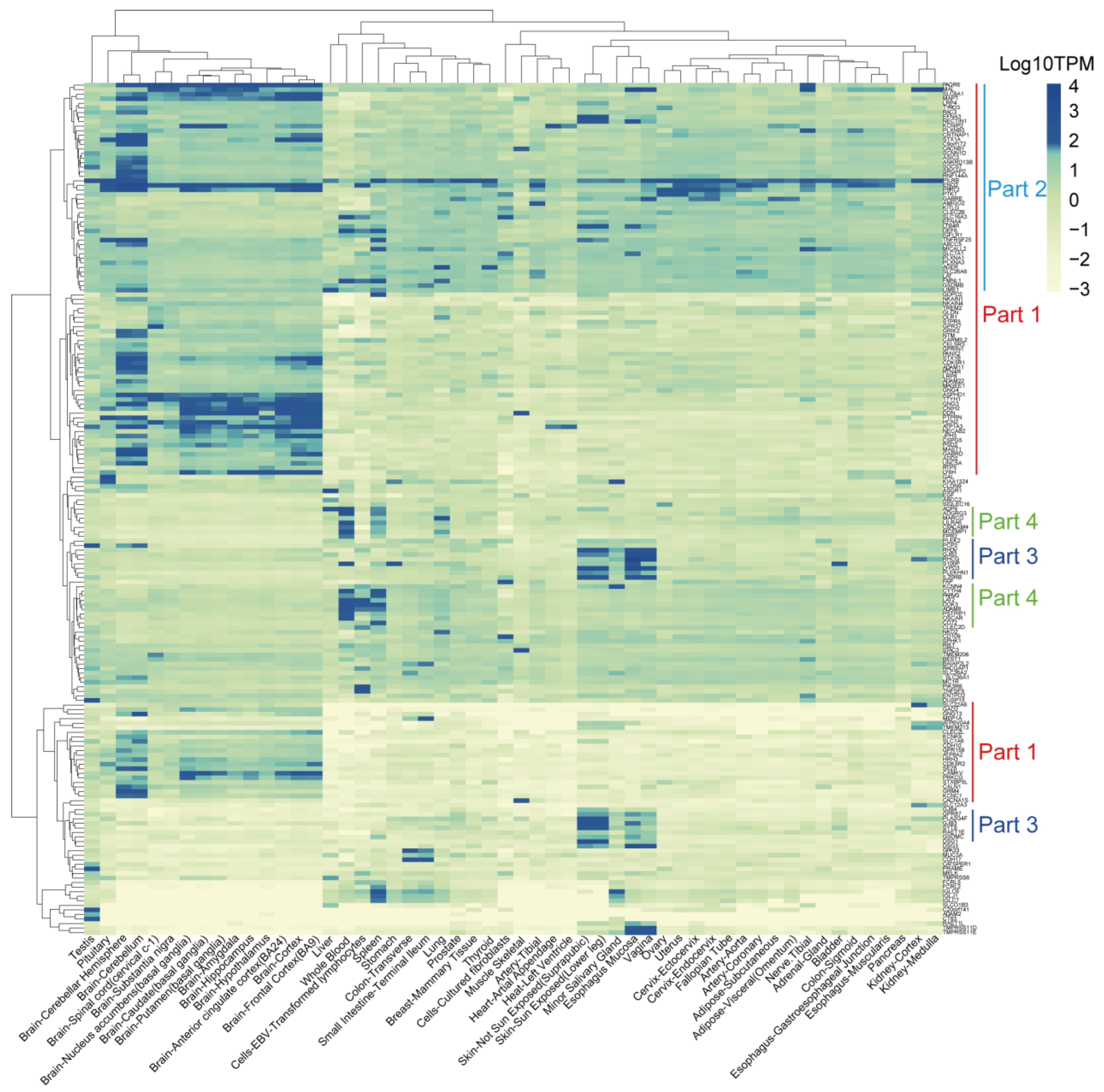
3.6. Identification of TSAs That Are Expressed Less in Normal Tissues
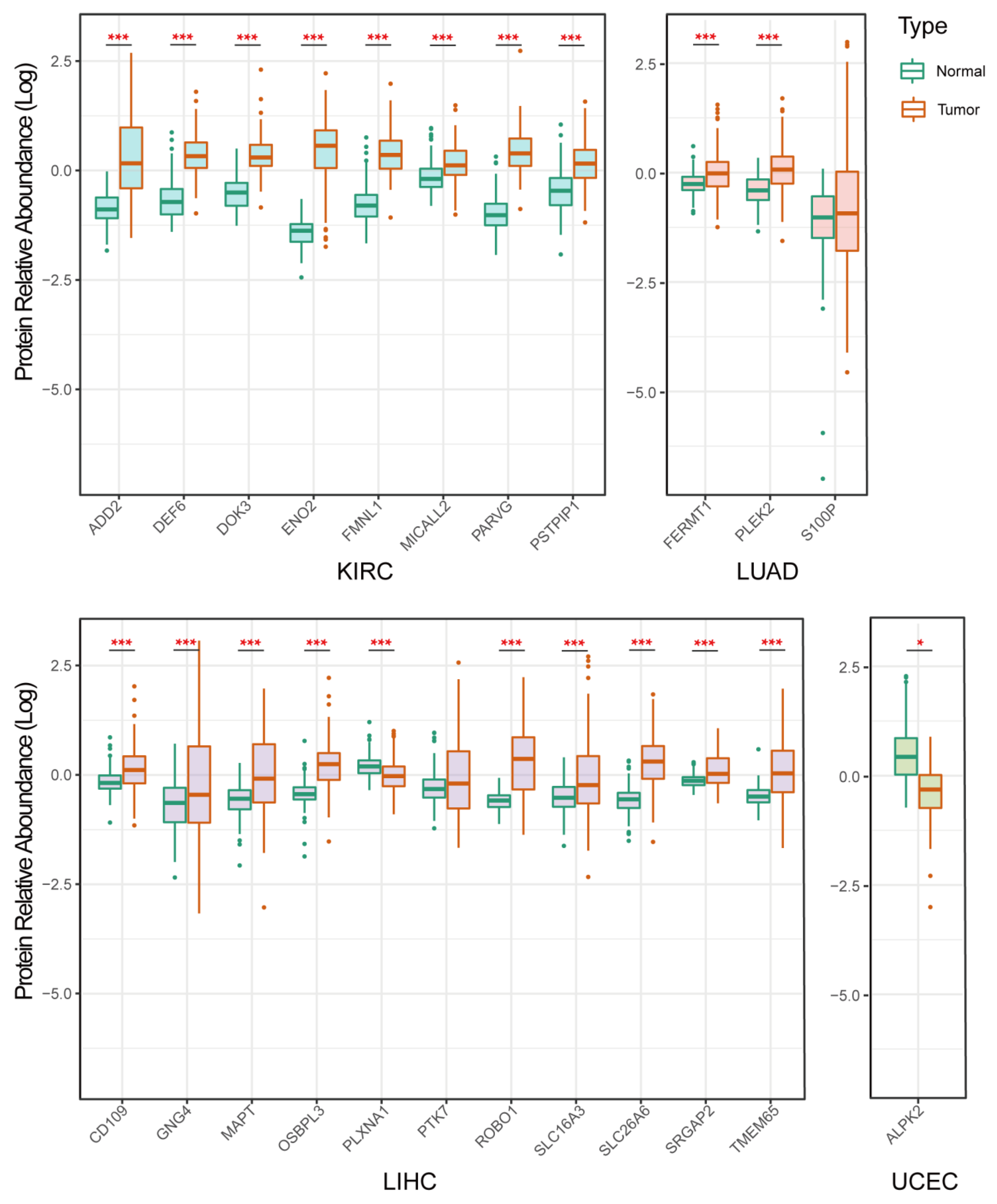
3.7. Validation of Protein Expression of the Selected Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The three Es of cancer immunoediting. Annu. Rev. Immunol. 2004, 22, 329–360. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; van der Leun, A.M.; Yofe, I.; Lubling, Y.; Gelbard-Solodkin, D.; van Akkooi, A.C.J.; van den Braber, M.; Rozeman, E.A.; Haanen, J.; Blank, C.U.; et al. Dysfunctional CD8 T Cells Form a Proliferative, Dynamically Regulated Compartment within Human Melanoma. Cell 2019, 176, 775–789.e18. [Google Scholar] [CrossRef]
- Feng, M.; Jiang, W.; Kim, B.Y.S.; Zhang, C.C.; Fu, Y.X.; Weissman, I.L. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 568–586. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.P.; Weissman, I.L.; Majeti, R. The CD47-SIRPα pathway in cancer immune evasion and potential therapeutic implications. Curr. Opin. Immunol. 2012, 24, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Urabe, F.; Kosaka, N.; Ito, K.; Kimura, T.; Egawa, S.; Ochiya, T. Extracellular vesicles as biomarkers and therapeutic targets for cancer. Am. J. Physiol. Cell Physiol. 2020, 318, C29–C39. [Google Scholar] [CrossRef]
- Li, H.; Ye, C.; Ji, G.; Han, J. Determinants of public T cell responses. Cell Res. 2012, 22, 33–42. [Google Scholar] [CrossRef]
- Paijens, S.T.; Vledder, A.; de Bruyn, M.; Nijman, H.W. Tumor-infiltrating lymphocytes in the immunotherapy era. Cell. Mol. Immunol. 2021, 18, 842–859. [Google Scholar] [CrossRef]
- Thomas, A.; Teicher, B.A.; Hassan, R. Antibody-drug conjugates for cancer therapy. Lancet Oncol. 2016, 17, e254–e262. [Google Scholar] [CrossRef]
- Nguyen, A.; Johanning, G.; Shi, Y. Emerging Novel Combined CAR-T Cell Therapies. Cancers 2022, 14, 1403. [Google Scholar] [CrossRef]
- Ellis, G.I.; Sheppard, N.C.; Riley, J.L. Genetic engineering of T cells for immunotherapy. Nat. Rev. Genet. 2021, 22, 427–447. [Google Scholar] [CrossRef]
- Turtle, C.J.; Hanafi, L.A.; Berger, C.; Gooley, T.A.; Cherian, S.; Hudecek, M.; Sommermeyer, D.; Melville, K.; Pender, B.; Budiarto, T.M.; et al. CD19 CAR-T cells of defined CD4+:CD8+ composition in adult B cell ALL patients. J. Clin. Investig. 2016, 126, 2123–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.Y.; Wang, Y.; Guo, Y.L.; Dai, H.R.; Yang, Q.M.; Zhang, Y.J.; Zhang, Y.; Chen, M.X.; Wang, C.M.; Feng, K.C.; et al. Treatment of CD20-directed Chimeric Antigen Receptor-modified T cells in patients with relapsed or refractory B-cell non-Hodgkin lymphoma: An early phase IIa trial report. Signal Transduct. Target. Ther. 2016, 1, 16002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Wang, Q.; Xu, H.; Gu, C.; Jiang, L.; Wang, J.; Wang, D.; Xu, B.; Mao, X.; Wang, J.; et al. Anti-BCMA CAR-T cells for treatment of plasma cell dyscrasia: Case report on POEMS syndrome and multiple myeloma. J. Hematol. Oncol. 2018, 11, 128. [Google Scholar] [CrossRef] [Green Version]
- Wilkie, S.; van Schalkwyk, M.C.; Hobbs, S.; Davies, D.M.; van der Stegen, S.J.; Pereira, A.C.; Burbridge, S.E.; Box, C.; Eccles, S.A.; Maher, J. Dual targeting of ErbB2 and MUC1 in breast cancer using chimeric antigen receptors engineered to provide complementary signaling. J. Clin. Immunol. 2012, 32, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Mount, C.W.; Majzner, R.G.; Sundaresh, S.; Arnold, E.P.; Kadapakkam, M.; Haile, S.; Labanieh, L.; Hulleman, E.; Woo, P.J.; Rietberg, S.P.; et al. Potent antitumor efficacy of anti-GD2 CAR T cells in H3-K27M(+) diffuse midline gliomas. Nat. Med. 2018, 24, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Harrer, D.C.; Dörrie, J.; Schaft, N. CSPG4 as Target for CAR-T-Cell Therapy of Various Tumor Entities-Merits and Challenges. Int. J. Mol. Sci. 2019, 20, 5942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, R.A.; Yang, J.C.; Kitano, M.; Dudley, M.E.; Laurencot, C.M.; Rosenberg, S.A. Case report of a serious adverse event following the administration of T cells transduced with a chimeric antigen receptor recognizing ERBB2. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 843–851. [Google Scholar] [CrossRef]
- Feng, K.; Guo, Y.; Dai, H.; Wang, Y.; Li, X.; Jia, H.; Han, W. Chimeric antigen receptor-modified T cells for the immunotherapy of patients with EGFR-expressing advanced relapsed/refractory non-small cell lung cancer. Sci. China Life Sci. 2016, 59, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Lo, A.; Wang, L.S.; Scholler, J.; Monslow, J.; Avery, D.; Newick, K.; O’Brien, S.; Evans, R.A.; Bajor, D.J.; Clendenin, C.; et al. Tumor-Promoting Desmoplasia Is Disrupted by Depleting FAP-Expressing Stromal Cells. Cancer Res. 2015, 75, 2800–2810. [Google Scholar] [CrossRef]
- Schubert, M.L.; Schmitt, M.; Wang, L.; Ramos, C.A.; Jordan, K.; Müller-Tidow, C.; Dreger, P. Side-effect management of chimeric antigen receptor (CAR) T-cell therapy. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2021, 32, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kaur, G.; Sankin, A.I.; Chen, F.; Guan, F.; Zang, X. Immune checkpoint blockade and CAR-T cell therapy in hematologic malignancies. J. Hematol. Oncol. 2019, 12, 59. [Google Scholar] [CrossRef]
- Boyiadzis, M.M.; Dhodapkar, M.V.; Brentjens, R.J.; Kochenderfer, J.N.; Neelapu, S.S.; Maus, M.V.; Porter, D.L.; Maloney, D.G.; Grupp, S.A.; Mackall, C.L.; et al. Chimeric antigen receptor (CAR) T therapies for the treatment of hematologic malignancies: Clinical perspective and significance. J. Immunother. Cancer 2018, 6, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miliotou, A.N.; Papadopoulou, L.C. CAR T-cell Therapy: A New Era in Cancer Immunotherapy. Curr. Pharm. Biotechnol. 2018, 19, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.; Goetsch, L.; Dumontet, C.; Corvaïa, N. Strategies and challenges for the next generation of antibody-drug conjugates. Nat. Rev. Drug Discov. 2017, 16, 315–337. [Google Scholar] [CrossRef]
- Autio, K.A.; Boni, V.; Humphrey, R.W.; Naing, A. Probody Therapeutics: An Emerging Class of Therapies Designed to Enhance On-Target Effects with Reduced Off-Tumor Toxicity for Use in Immuno-Oncology. Clin. Cancer Res. 2020, 26, 984–989. [Google Scholar] [CrossRef] [Green Version]
- Lomize, A.L.; Lomize, M.A.; Krolicki, S.R.; Pogozheva, I.D. Membranome: A database for proteome-wide analysis of single-pass membrane proteins. Nucleic Acids Res. 2017, 45, D250–D255. [Google Scholar] [CrossRef] [Green Version]
- Lomize, A.L.; Hage, J.M.; Pogozheva, I.D. Membranome 2.0: Database for proteome-wide profiling of bitopic proteins and their dimers. Bioinformatics 2018, 34, 1061–1062. [Google Scholar] [CrossRef] [Green Version]
- UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [CrossRef]
- Zhang, X.; Lan, Y.; Xu, J.; Quan, F.; Zhao, E.; Deng, C.; Luo, T.; Xu, L.; Liao, G.; Yan, M.; et al. CellMarker: A manually curated resource of cell markers in human and mouse. Nucleic Acids Res. 2019, 47, D721–D728. [Google Scholar] [CrossRef]
- GTEx Consortium. Human genomics. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakur, A.H.; Huang, S.; Qian, X.; Chang, X. SURVFIT: Doubly sparse rule learning for survival data. J. Biomed. Inform. 2021, 117, 103691. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Li, Z.; Zhu, X.; Yan, P.; Song, D.; Yin, H.; Hu, P.; Lin, R.; Wu, S.; Meng, T.; et al. Collagen Type III Alpha 1 chain regulated by GATA-Binding Protein 6 affects Type II IFN response and propanoate metabolism in the recurrence of lower grade glioma. J. Cell. Mol. Med. 2020, 24, 10803–10815. [Google Scholar] [CrossRef]
- Ouyang, Z.; Li, G.; Zhu, H.; Wang, J.; Qi, T.; Qu, Q.; Tu, C.; Qu, J.; Lu, Q. Construction of a Five-Super-Enhancer-Associated-Genes Prognostic Model for Osteosarcoma Patients. Front. Cell Dev. Biol. 2020, 8, 598660. [Google Scholar] [CrossRef]
- Li, J.; Xiang, L.; Wang, Q.; Ma, X.; Chen, X.; Zhu, Y.; Yang, Y.; Huang, L.; He, H.; Xu, L.; et al. Highly Potent Immunotoxins Targeting the Membrane-distal N-lobe of GPC3 for Immunotherapy of Hepatocellular Carcinoma. J. Cancer 2022, 13, 1370–1384. [Google Scholar] [CrossRef]
- Reinhard, K.; Rengstl, B.; Oehm, P.; Michel, K.; Billmeier, A.; Hayduk, N.; Klein, O.; Kuna, K.; Ouchan, Y.; Wöll, S.; et al. An RNA vaccine drives expansion and efficacy of claudin-CAR-T cells against solid tumors. Science 2020, 367, 446–453. [Google Scholar] [CrossRef]
- Tang, X.; Liu, F.; Liu, Z.; Cao, Y.; Zhang, Z.; Wang, Y.; Huang, J.; Fan, S.; Zhao, S.; Chen, Y.; et al. Bioactivity and safety of B7-H3-targeted chimeric antigen receptor T cells against anaplastic meningioma. Clin. Transl. Immunol. 2020, 9, e1137. [Google Scholar] [CrossRef]
- Lamers, C.H.; Sleijfer, S.; van Steenbergen, S.; van Elzakker, P.; van Krimpen, B.; Groot, C.; Vulto, A.; den Bakker, M.; Oosterwijk, E.; Debets, R.; et al. Treatment of metastatic renal cell carcinoma with CAIX CAR-engineered T cells: Clinical evaluation and management of on-target toxicity. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21, 904–912. [Google Scholar] [CrossRef]
- Wu, X.; Luo, H.; Shi, B.; Di, S.; Sun, R.; Su, J.; Liu, Y.; Li, H.; Jiang, H.; Li, Z. Combined Antitumor Effects of Sorafenib and GPC3-CAR T Cells in Mouse Models of Hepatocellular Carcinoma. Mol. Ther. J. Am. Soc. Gene Ther. 2019, 27, 1483–1494. [Google Scholar] [CrossRef]
- Shi, D.; Shi, Y.; Kaseb, A.O.; Qi, X.; Zhang, Y.; Chi, J.; Lu, Q.; Gao, H.; Jiang, H.; Wang, H.; et al. Chimeric Antigen Receptor-Glypican-3 T-Cell Therapy for Advanced Hepatocellular Carcinoma: Results of Phase I Trials. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 3979–3989. [Google Scholar] [CrossRef] [PubMed]
- Shiina, S.; Ohno, M.; Ohka, F.; Kuramitsu, S.; Yamamichi, A.; Kato, A.; Motomura, K.; Tanahashi, K.; Yamamoto, T.; Watanabe, R.; et al. CAR T Cells Targeting Podoplanin Reduce Orthotopic Glioblastomas in Mouse Brains. Cancer Immunol. Res. 2016, 4, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Berahovich, R.; Zhou, H.; Liu, X.; Li, F.; Xu, S.; Wei, Y.; Ouaret, D.; Bodmer, W.; Wu, L.; et al. PLAP -CAR T cells mediate high specific cytotoxicity against colon cancer cells. Front. Biosci. 2020, 25, 1765–1786. [Google Scholar] [CrossRef]
- Byrd, T.T.; Fousek, K.; Pignata, A.; Szot, C.; Samaha, H.; Seaman, S.; Dobrolecki, L.; Salsman, V.S.; Oo, H.Z.; Bielamowicz, K.; et al. TEM8/ANTXR1-Specific CAR T Cells as a Targeted Therapy for Triple-Negative Breast Cancer. Cancer Res. 2018, 78, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Shi, Z.; Wang, P.; Wang, C.; Yang, L.; Du, G.; Zhang, H.; Shi, B.; Jia, J.; Li, Q.; et al. Claudin18.2-Specific Chimeric Antigen Receptor Engineered T Cells for the Treatment of Gastric Cancer. J. Natl. Cancer Inst. 2019, 111, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.L.; Morgan, R.A.; Yang, J.C.; Sherry, R.M.; Robbins, P.F.; Restifo, N.P.; Feldman, S.A.; Lu, Y.C.; Lu, L.; Zheng, Z.; et al. Pilot Trial of Adoptive Transfer of Chimeric Antigen Receptor-transduced T Cells Targeting EGFRvIII in Patients With Glioblastoma. J. Immunother. 2019, 42, 126–135. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, Y.; Wu, Z.; Feng, K.; Tong, C.; Wang, Y.; Dai, H.; Shi, F.; Yang, Q.; Han, W. Anti-EGFR chimeric antigen receptor-modified T cells in metastatic pancreatic carcinoma: A phase I clinical trial. Cytotherapy 2020, 22, 573–580. [Google Scholar] [CrossRef]
- Ahmed, N.; Brawley, V.; Hegde, M.; Bielamowicz, K.; Kalra, M.; Landi, D.; Robertson, C.; Gray, T.L.; Diouf, O.; Wakefield, A.; et al. HER2-Specific Chimeric Antigen Receptor-Modified Virus-Specific T Cells for Progressive Glioblastoma: A Phase 1 Dose-Escalation Trial. JAMA Oncol. 2017, 3, 1094–1101. [Google Scholar] [CrossRef]
- Kloss, C.C.; Lee, J.; Zhang, A.; Chen, F.; Melenhorst, J.J.; Lacey, S.F.; Maus, M.V.; Fraietta, J.A.; Zhao, Y.; June, C.H. Dominant-Negative TGF-β Receptor Enhances PSMA-Targeted Human CAR T Cell Proliferation And Augments Prostate Cancer Eradication. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 1855–1866. [Google Scholar] [CrossRef] [Green Version]
- Magee, M.S.; Kraft, C.L.; Abraham, T.S.; Baybutt, T.R.; Marszalowicz, G.P.; Li, P.; Waldman, S.A.; Snook, A.E. GUCY2C-directed CAR-T cells oppose colorectal cancer metastases without autoimmunity. Oncoimmunology 2016, 5, e1227897. [Google Scholar] [CrossRef]
- Sharma, P.; Debinski, W. Receptor-Targeted Glial Brain Tumor Therapies. Int. J. Mol. Sci. 2018, 19, 3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, M.K.; Ohishi, T.; Kawada, M.; Kato, Y. A cancer-specific anti-podocalyxin monoclonal antibody (60-mG(2a)-f) exerts antitumor effects in mouse xenograft models of pancreatic carcinoma. Biochem. Biophys. Rep. 2020, 24, 100826. [Google Scholar] [CrossRef] [PubMed]
- Priceman, S.J.; Gerdts, E.A.; Tilakawardane, D.; Kennewick, K.T.; Murad, J.P.; Park, A.K.; Jeang, B.; Yamaguchi, Y.; Yang, X.; Urak, R.; et al. Co-stimulatory signaling determines tumor antigen sensitivity and persistence of CAR T cells targeting PSCA+ metastatic prostate cancer. Oncoimmunology 2018, 7, e1380764. [Google Scholar] [CrossRef]
- Jie, Y.; Liu, G.; Feng, L.; Li, Y.; E, M.; Wu, L.; Li, Y.; Rong, G.; Li, Y.; Wei, H.; et al. PTK7-Targeting CAR T-Cells for the Treatment of Lung Cancer and Other Malignancies. Front. Immunol. 2021, 12, 665970. [Google Scholar] [CrossRef] [PubMed]
- Ao, X.; Yang, Y.; Li, W.; Tan, Y.; Guo, W.; Ao, L.; He, X.; Wu, X.; Xia, J.; Xu, X.; et al. Anti-αFR CAR-engineered NK-92 Cells Display Potent Cytotoxicity Against αFR-positive Ovarian Cancer. J. Immunother. 2019, 42, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zhao, R.; Wu, D.; Zheng, D.; Wu, Z.; Shi, J.; Wei, X.; Wu, Q.; Long, Y.; Lin, S.; et al. Mesothelin is a target of chimeric antigen receptor T cells for treating gastric cancer. J. Hematol. Oncol. 2019, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatty, G.L.; O’Hara, M.H.; Lacey, S.F.; Torigian, D.A.; Nazimuddin, F.; Chen, F.; Kulikovskaya, I.M.; Soulen, M.C.; McGarvey, M.; Nelson, A.M.; et al. Activity of Mesothelin-Specific Chimeric Antigen Receptor T Cells Against Pancreatic Carcinoma Metastases in a Phase 1 Trial. Gastroenterology 2018, 155, 29–32. [Google Scholar] [CrossRef]
- Rajabzadeh, A.; Rahbarizadeh, F.; Ahmadvand, D.; Kabir Salmani, M.; Hamidieh, A.A. A VHH-Based Anti-MUC1 Chimeric Antigen Receptor for Specific Retargeting of Human Primary T Cells to MUC1-Positive Cancer Cells. Cell J. 2021, 22, 502–513. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.; Zhang, M.; Gong, J.; Li, Q.; Shan, B.; Wang, T.; Zhang, L.; Zheng, T.; Li, X. The aberrant expression of rhythm genes affects the genome instability and regulates the cancer immunity in pan-cancer. Cancer Med. 2020, 9, 1818–1829. [Google Scholar] [CrossRef]
- Schettini, F.; Barbao, P.; Brasó-Maristany, F.; Galván, P.; Martínez, D.; Paré, L.; De Placido, S.; Prat, A.; Guedan, S. Identification of cell surface targets for CAR-T cell therapies and antibody-drug conjugates in breast cancer. ESMO Open 2021, 6, 100102. [Google Scholar] [CrossRef]
- Fraietta, J.A.; Lacey, S.F.; Orlando, E.J.; Pruteanu-Malinici, I.; Gohil, M.; Lundh, S.; Boesteanu, A.C.; Wang, Y.; O’Connor, R.S.; Hwang, W.T.; et al. Determinants of response and resistance to CD19 chimeric antigen receptor (CAR) T cell therapy of chronic lymphocytic leukemia. Nat. Med. 2018, 24, 563–571. [Google Scholar] [CrossRef]
- Vairy, S.; Garcia, J.L.; Teira, P.; Bittencourt, H. CTL019 (tisagenlecleucel): CAR-T therapy for relapsed and refractory B-cell acute lymphoblastic leukemia. Drug Des. Dev. Ther. 2018, 12, 3885–3898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, D.; Tomiuk, S.; Küster, L.N.; Rawashdeh, W.A.; Henze, J.; Tischler-Höhle, G.; Agorku, D.J.; Brauner, J.; Linnartz, C.; Lock, D.; et al. Identification of CD318, TSPAN8 and CD66c as target candidates for CAR T cell based immunotherapy of pancreatic adenocarcinoma. Nat. Commun. 2021, 12, 1453. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, P.; Velasquez, M.P.; Gottschalk, S.; Zhang, J.; Fan, Y. Unifying heterogeneous expression data to predict targets for CAR-T cell therapy. Oncoimmunology 2021, 10, 2000109. [Google Scholar] [CrossRef] [PubMed]
- Theruvath, J.; Sotillo, E.; Mount, C.W.; Graef, C.M.; Delaidelli, A.; Heitzeneder, S.; Labanieh, L.; Dhingra, S.; Leruste, A.; Majzner, R.G.; et al. Locoregionally administered B7-H3-targeted CAR T cells for treatment of atypical teratoid/rhabdoid tumors. Nat. Med. 2020, 26, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Dong, Q.; Xu, Y.; Yu, G.; Zhang, X. B7-H3: Another molecule marker for Mo-DCs? Cell. Mol. Immunol. 2005, 2, 307–311. [Google Scholar]
- Nair, K.S.; Zingde, S.M. Adhesion of neutrophils to fibronectin: Role of the cd66 antigens. Cell. Immunol. 2001, 208, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Ge, H.; Long, Y.; Yang, C.; Chang, Y.E.; Mu, L.; Sayour, E.J.; De Leon, G.; Wang, Q.J.; Yang, J.C.; et al. CD70, a novel target of CAR T-cell therapy for gliomas. Neuro-Oncology 2018, 20, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Lantto, R.; Nasi, A.; Sammicheli, S.; Amu, S.; Fievez, V.; Moutschen, M.; Pensieroso, S.; Hejdeman, B.; Chiodi, F.; Rethi, B. Increased extrafollicular expression of the B-cell stimulatory molecule CD70 in HIV-1-infected individuals. AIDS 2015, 29, 1757–1766. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Chang, K.; Xu, C.; Li, Q.; Du, Z. High expression of DLL3 is associated with a poor prognosis and immune infiltration in invasive breast cancer patients. Transl. Oncol. 2021, 14, 101080. [Google Scholar] [CrossRef]
- Ding, Z.C.; Habtetsion, T.; Cao, Y.; Li, T.; Liu, C.; Kuczma, M.; Chen, T.; Hao, Z.; Bryan, L.; Munn, D.H.; et al. Adjuvant IL-7 potentiates adoptive T cell therapy by amplifying and sustaining polyfunctional antitumor CD4+ T cells. Sci. Rep. 2017, 7, 12168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cheng, C.; Sun, W.; Wang, H. Engineering T Cells Using CRISPR/Cas9 for Cancer Therapy. Methods Mol. Biol. 2020, 2115, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Sau, S.; Alsaab, H.O.; Bhise, K.; Alzhrani, R.; Nabil, G.; Iyer, A.K. Multifunctional nanoparticles for cancer immunotherapy: A groundbreaking approach for reprogramming malfunctioned tumor environment. J. Control. Release Off. J. Control. Release Soc. 2018, 274, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, J. Therapeutic effect of dual CAR-T targeting PDL1 and MUC16 antigens on ovarian cancer cells in mice. BMC Cancer 2020, 20, 678. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Yang, P.; Gu, J.; Wu, H.; Chi, X.; Liu, C.; Wang, Y.; Xue, J.; Qi, W.; Sun, Q.; et al. Recombination of a dual-CAR-modified T lymphocyte to accurately eliminate pancreatic malignancy. J. Hematol. Oncol. 2018, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- DeLucia, D.C.; Lee, J.K. Identification of Cell Surface Targets for CAR T Cell Immunotherapy. Methods Mol. Biol. 2020, 2097, 45–54. [Google Scholar] [CrossRef]
- Kuhlmann, L.; Cummins, E.; Samudio, I.; Kislinger, T. Cell-surface proteomics for the identification of novel therapeutic targets in cancer. Expert Rev. Proteom. 2018, 15, 259–275. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Abbreviation | TCGA | CPTAC | |||
|---|---|---|---|---|---|---|
| Tumor Sample | Normal Sample | Tumor Sample | Normal Sample | PDC Study ID | ||
| Bladder urothelial carcinoma | BLCA | 411 | 19 | |||
| Breast invasive carcinoma | BRCA | 1104 | 113 | 133 | 18 | PDC000120 |
| Colon adenocarcinoma | COAD | 471 | 41 | 95 | 100 | PDC000116 |
| Cholangiocarcinoma | CHOL | 36 | 9 | |||
| Esophageal carcinoma | ESCA | 162 | 11 | |||
| Head and neck squamous cell carcinoma | HNSC | 502 | 44 | |||
| Kidney chromophobe | KICH | 65 | 24 | |||
| Kidney renal clear cell carcinoma | KIRC | 535 | 72 | 110 | 84 | PDC000127 |
| Kidney renal papillary cell carcinoma | KIRP | 289 | 32 | |||
| Liver hepatocellular carcinoma | LIHC | 374 | 50 | 165 | 165 | PDC000198 |
| Lung adenocarcinoma | LUAD | 526 | 59 | 113 | 102 | PDC000153 |
| Lung squamous cell carcinoma | LUSC | 501 | 49 | 110 | 104 | PDC000224 |
| Prostate adenocarcinoma | PRAD | 499 | 52 | |||
| Rectum adenocarcinoma | READ | 167 | 10 | |||
| Stomach adenocarcinoma | STAD | 375 | 32 | |||
| Thyroid carcinoma | THCA | 510 | 58 | |||
| Uterine corpus endometrial carcinoma | UCEC | 548 | 35 | 100 | 49 | PDC000125 |
| Cancer | High-Expressed and High-Risk Membrane Proteins in Pan-Cancer | Total |
|---|---|---|
| BLCA | CHRNA1, DRD5, HCN2, LRP8, RAC3, SLC1A6 | 6 |
| BRCA | FIBCD1, MAFA, PCDHA1, RGS9BP, SEZ6, ULBP2 | 6 |
| CHOL | RAET1E, RTBDN, TMEM17 | 3 |
| COAD | ASPHD1, CCDC78, CLDN6, DLL3, ELFN1, GABRD, GJA3, GNGT1, GPC2, KREMEN2, LIME1, LRRN4, MC1R, PANX2, PCP2, S1PR5, SLC16A8, SLC6A1, SYT8, TMEM213, TRPV4, UNC5A | 22 |
| ESCA | OR2W6P | 1 |
| HNSC | AMIGO2, DCBLD1, DSCAM, EMR1, FRMD5, MCEMP1, OLR1, OR11H7, OR5M11, RTN4R, TMEM92 | 11 |
| KICH | ADAM2, CSPG5, GSDMC, KCNC1, LRRTM3, MELK, SIGLEC15, TMPRSS11D | 8 |
| KIRC | ADAM11, ADAM12, ADAM8, ADD2, AGER, AQP9, ARHGEF39, ASGR1, ASIC3, ATP1A3, BEST1, BEST4, BTNL10, C20orf141, C9orf172, CABP7, CACNA2D4, CACNB1, CATSPER1, CCDC78, CD72, CDHR4, CDK5R2, CEACAM4, CELSR3, CLEC12B, CLEC2B, CLEC2D, CNIH2, CNTNAP1, CPNE7, CYTH4, DEF6, DLK2, DOK3, DRD4, EFNA3, EMR1, ENO2, FAP, FCGR1B, FCGR1C, FCRL2, FCRL5, FFAR3, FMNL1, FPR2,GABRE, GAD2, GGT2, GNG13, GPC2, GPR171, GPR19, GPR45, GPR84, GPR97, GPRIN1, GRIK4, GRIN2D, GRM2, GRM4, GSDMB, IGFLR1, IGLC6, IGLC7, IGLJ1, IL20RB, IL31RA, JPH3, KCNIP2, KCNN4, KIAA1324, KLRC4, KREMEN2, LAT, LAT2, LILRA6, LSMEM1, LTB4R, LTB4R2, LY6H, MARCO, MB21D1, MC1R, MCEMP1, MELK, MICALL2, MILR1, MMP17, MUC12, MUC3A, NPFFR1, NPY4R, NTM, NTNG2, OPN4, OR11H7, OR13A1, OR2B6, OSCAR, OTOF, PAQR6, PARVG, PCDHGC5, PDE6C, PIK3R6, PILRB, PLB1, PLEKHN1, PLXDC1, PLXNB3, PRAME, PRKCG, PRR7, PRRT2, PSD2, PSTPIP1, PTPRN, RASL10A, RELT, RLTPR, RTP5, SCNN1D, SIGLEC16, SLAMF8, SLAMF9, SLC16A8, SLCO5A1, SMIM23, SPHK1, STAC3, STX1B, TBC1D3B, TBC1D3F, TBC1D3L, TMPRSS6, TNFRSF25, TNFSF14, TPBGL, TRABD2A, TRBV28, TREML1, TRPM2, TRPM8, TTYH1, ZGRF1, ZP1 | 148 |
| KIRP | ADAM18, ADD2, ASIC3, CALN1, CDH17, CHRNA5, DLL3, GABRD, GJB4, GPA33, GPR19, GPRIN1, HTR1D, HTR3A, IL20RB, KIAA0319, KREMEN2, MELK, MFSD2B, OTOF, PTPRN, RTBDN, TMPRSS15, TNFSF9, ZP1 | 25 |
| LIHC | ABCC5, ADAM22, ANKRD13B, ANKRD27, ARHGEF39, ATP8A2, BAIAP2L2, BFSP1, CACNA1S, CD109, CDH10, CDK5R1, CHRNA5, CLCN2, CLEC2L, CNTNAP1, CSMD1, CSPG5, CT83, DAGLA, DDN, DUSP15, EFCAB7, EFNA3, EFNA4, EFNA5EGF, ENTPD2, EPHA6, FAM171A2, FIBCD1, FLVCR1, GABRA3, GDPD2, GJD4, GLDN, GLP1R, GNG4, GNGT1, GPR156, GPRC5D, GPRIN1, GPSM2, GRIN2D, GRM4, GSDMC, IGSF3, KCNE5, KCNQ3, KCTD7, KISS1R, KITLG, LRP12, LRP4, LRP8, LYSMD4, MAGEE1, MAPT, MELK, MEP1A, OR52E6, OR8A1, OR8G3P, OSBPL3, P2RY4, PDE6A, PIK3R6, PLXNA1, PLXNA3, PRR7, PRSS42, PTK7, PTPRN, PVRL1, RACGAP1, RAET1E, RNF144A, ROBO1, SAPCD2, SEMA4F, SHISA7, SLC16A3, SLC22A6, SLC26A2, SLC26A6, SLC30A8, SLC36A1, SLC38A6, SLC47A2, SLC7A1, SOCS7, SRGAP2, STX1A, STXBP5L, TBC1D30, TMEM206, TMEM213, TMEM65, TMEM67, TMEM81, TNFRSF11A, TREM2, TYRO3, XKR3, ZP3 | 105 |
| LUAD | ABCC2, ALPI, BAIAP2L2, C20orf141, CDH17, CDK5R1, CLDN14, DSG3, ENTPD2, FERMT1, GDPD2, GJA3, GJB2, GJB3, GJB4, GPR37, GPR78, GPR87, GPRIN1, GRIK2, HCN2, HTR1D, IL20RB, KCNV1, KIAA1549L, LGR4, LY6K, LYPD3, MELK, MFI2, OR10J6P, OR1F1, PLEK2, PTPRH, RAET1L, RHCG, RHOV, S100P, SAPCD2, SLCO1B3, STYK1, TMPRSS11E, TRPA1, UNC5D | 44 |
| PRAD | GPC2, KISS1R, NECAB2, PCDHA2, PCDHA9, SLC17A4 | 6 |
| READ | ATP6V0A4 | 1 |
| STAD | ADAM12, CLDN9, HTR1F, KIAA1549L, MCEMP1, NOX4, OR10A5, OR6A2, SLC22A16, TREML4 | 10 |
| THCA | DRD5, EFNA2, LHFPL5, MAST1, MCEMP1, NKD2, OR1F1, PCDHA2, PCDHA5, RTBDN, SLAMF9 | 11 |
| UCEC | ADAM18, ALPK2, ATP1A3, CABP7, CAMKV, CDH18, CDK5R2, CELSR3, CLDN6, CLDN9, DLL3, DSG1, GAL, GNG3, GPR110, GPR158, GPR19, GRIN2B, HEPHL1, HRH3, HTR3A, HTR3E, HTR6, KCNK9, KCNQ3, KCNS1, KIAA1549L, KLRG2, LIPH, MAFA, MAL, MAST1, MEP1A, MUC17, MUC3A, NKAIN1, NKAIN4, NTSR1, OTOG, P2RY2, PLA2G4F, PTPRN, RAC3, SHISA9, SLC12A3, SLC16A10, SLCO4C1 | 47 |
| Protein Name | Cancer Type | Reference |
|---|---|---|
| CLDN6 | Testicular, ovarian, uterine and lung adenocarcinoma | [37] |
| CD276 | Anaplastic meningioma | [38] |
| CA9 | Metastatic Renal Cell Carcinoma | [39] |
| GPC3 | Hepatocellular Carcinoma | [40,41] |
| PDPN | Glioblastomas | [42] |
| ALPP | Colorectal cancer | [43] |
| ANTXR1 | Triple-negative breast cancer | [44] |
| CLDN18 | Gastric cancer | [45] |
| EGFR | Glioblastoma, Pancreatic carcinoma | [46,47] |
| HER2 | Glioblastoma | [48] |
| FOLH1 | Prostate cancer | [49] |
| GUCY2C | Colorectal cancer | [50] |
| IL13RA2 | Glioblastoma | [51] |
| PODXL | Pancreatic carcinoma | [52] |
| PSCA | Prostate cancer | [53] |
| PTK7 | Lung cancer | [54] |
| FOLR1 | Ovarian Cancer | [55] |
| MSLN | Gastric cancer, Pancreatic Carcinoma | [56,57] |
| MUC1 | MUC1-positive cancer cells | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhou, J.; Zhang, W.; You, W.; Wang, J.; Zhou, L.; Liu, L.; Chen, W.-W.; Li, H. Pan-Cancer Analysis Identifies Tumor Cell Surface Targets for CAR-T Cell Therapies and Antibody Drug Conjugates. Cancers 2022, 14, 5674. https://doi.org/10.3390/cancers14225674
Li X, Zhou J, Zhang W, You W, Wang J, Zhou L, Liu L, Chen W-W, Li H. Pan-Cancer Analysis Identifies Tumor Cell Surface Targets for CAR-T Cell Therapies and Antibody Drug Conjugates. Cancers. 2022; 14(22):5674. https://doi.org/10.3390/cancers14225674
Chicago/Turabian StyleLi, Xinhui, Jian Zhou, Weiwen Zhang, Wenhua You, Jun Wang, Linlin Zhou, Lei Liu, Wei-Wei Chen, and Hanjie Li. 2022. "Pan-Cancer Analysis Identifies Tumor Cell Surface Targets for CAR-T Cell Therapies and Antibody Drug Conjugates" Cancers 14, no. 22: 5674. https://doi.org/10.3390/cancers14225674
APA StyleLi, X., Zhou, J., Zhang, W., You, W., Wang, J., Zhou, L., Liu, L., Chen, W. -W., & Li, H. (2022). Pan-Cancer Analysis Identifies Tumor Cell Surface Targets for CAR-T Cell Therapies and Antibody Drug Conjugates. Cancers, 14(22), 5674. https://doi.org/10.3390/cancers14225674