
Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota

 , , ,
, , , 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Gut Dysbiosis and Carcinogenesis
3. Gut Microbial Alteration, Chemotherapy, and Cancer Prevention
4. Spice-Derived Phytochemicals and CRC Prevention by Modulating Gut Bacteria for In Vivo Studies
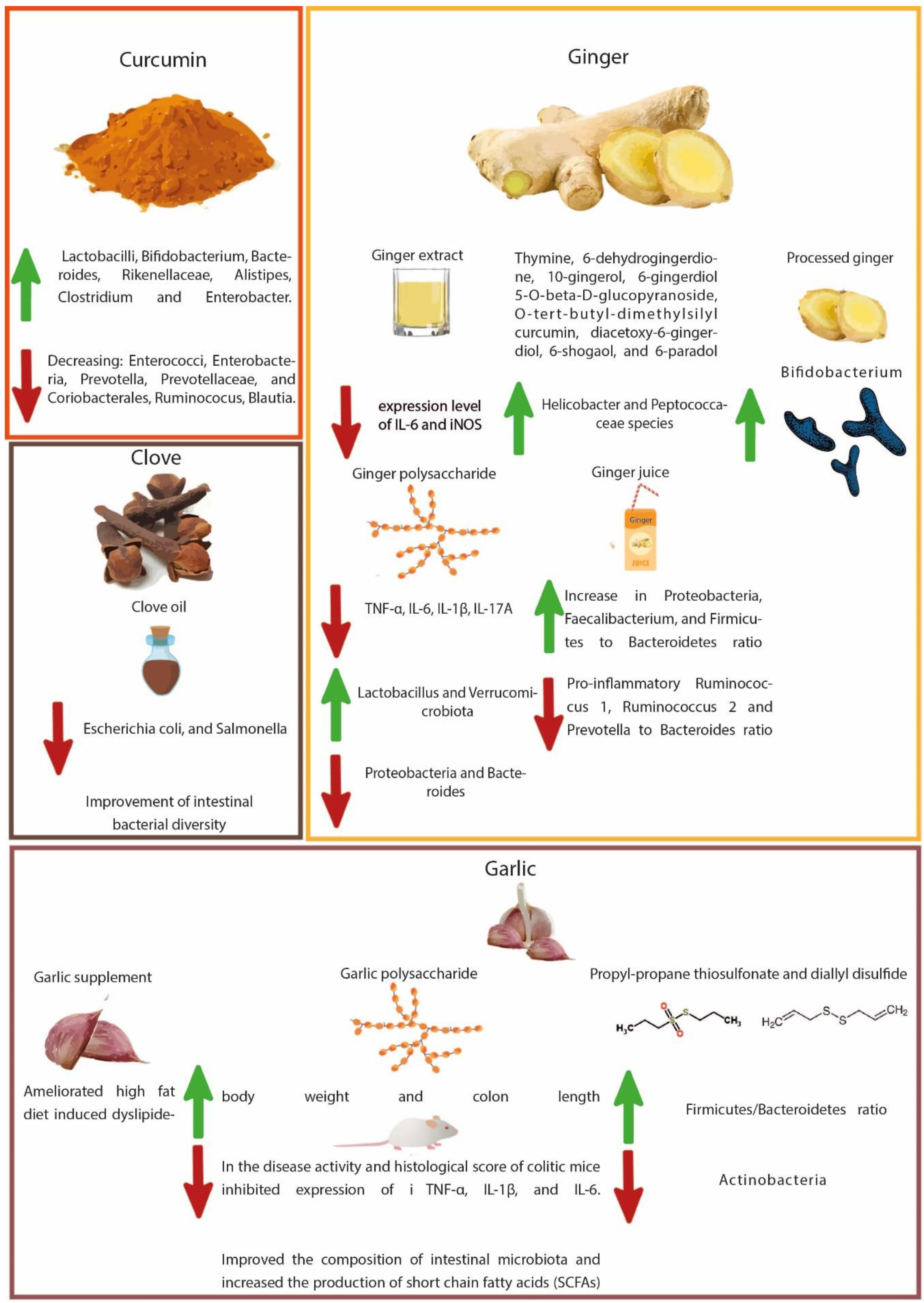
4.1. Turmeric-Derived Compounds
4.2. Ginger-Derived Compounds
4.3. Garlic-Derived Compounds
4.4. Clove-Derived Compounds
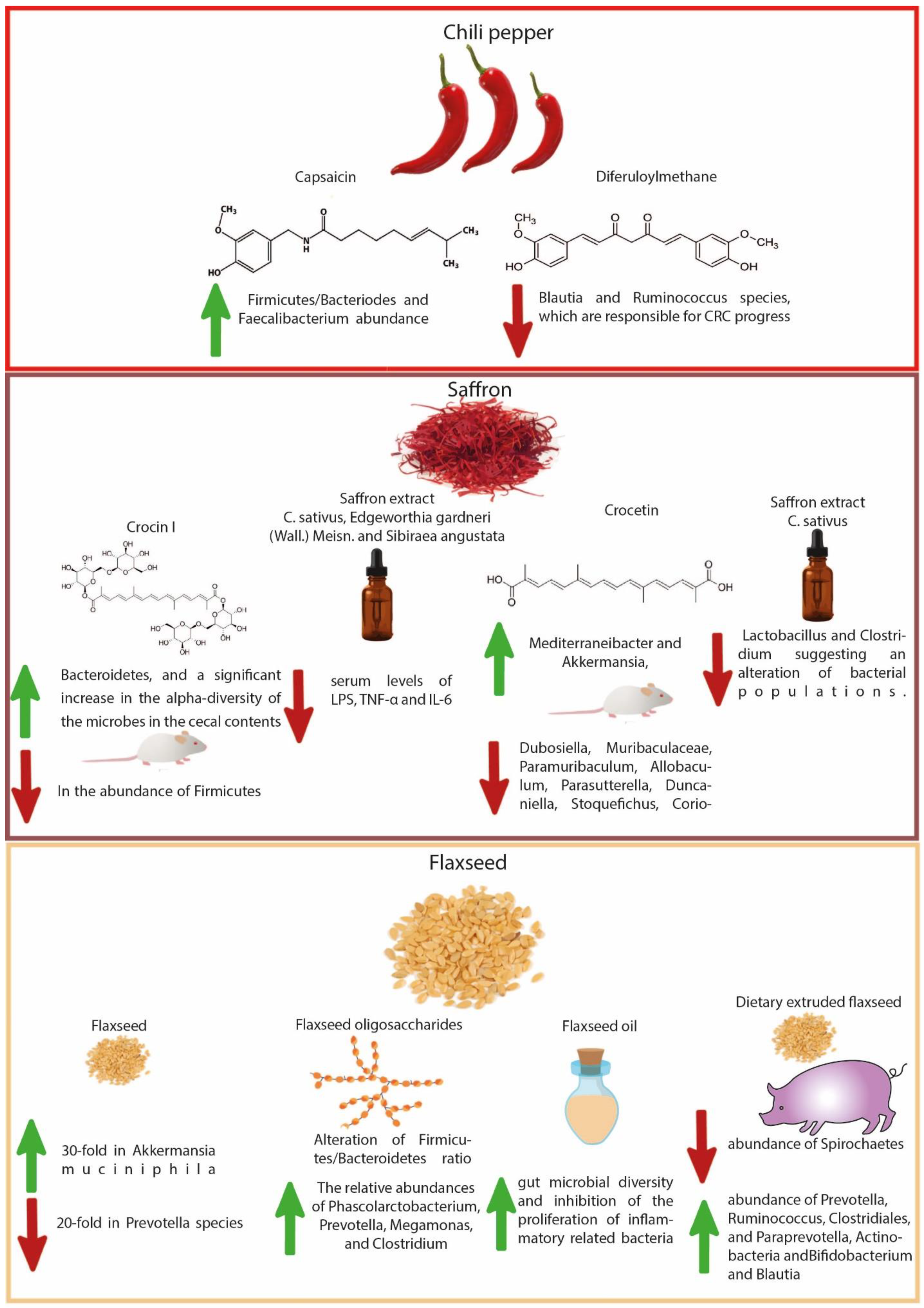
4.5. Chili Pepper-Derived Compounds
4.6. Saffron-Derived Compounds
4.7. Flaxseed-Derived Compounds
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CRC | colorectal cancer |
| CSC | cancer stem cells |
| CTLA-4 | T lymphocytes associated with antigen 4 |
| DSS | dextran sulfate sodium |
| IBD | inflammatory bowel disease |
| IL-1 | interleukin-1 |
| JNK pathway | c-Jun N-terminal kinase pathway |
| LPS | Lipopolysaccharide |
| MAPKs | mitogen activated protein kinases |
| MMPs | matrix metalloproteinases |
| NF-ĸB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK cells | natural killer cells |
| SCFAs | short chain fatty acids |
| TNF-α | tumor necrosis factor alpha |
References
- Vogelaar, I.; van Ballegooijen, M.; Schrag, D.; Boer, R.; Winawer, S.J.; Habbema, J.D.F.; Zauber, A.G. How Much Can Current Interventions Reduce Colorectal Cancer Mortality in the US? Mortality Projections for Scenarios of Risk-factor Modification, Screening, and Treatment. Cancer 2006, 107, 1624–1633. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fedewa, S.A.; Ahnen, D.J.; Meester, R.G.; Barzi, A.; Jemal, A. Colorectal Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 177–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.; Kasi, P.M.; Wallace, M.B. Colorectal Cancer. Lancet 2019, 10207, 1467–1480. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Zhou, J.; Zhu, Y.; Luo, L.; He, T.; Hu, H.; Liu, H.; Zhang, Y.; Luo, D.; Xu, S.; et al. Abdominal Obesity and Colorectal Cancer Risk: Systematic Review and Meta-Analysis of Prospective Studies. Biosci. Rep. 2017, 37, BSR20170945. [Google Scholar] [CrossRef] [Green Version]
- Liang, P.S.; Chen, T.Y.; Giovannucci, E. Cigarette Smoking and Colorectal Cancer Incidence and Mortality: Systematic Review and Meta-analysis. Int. J. Cancer 2009, 124, 2406–2415. [Google Scholar] [CrossRef]
- McNabb, S.; Harrison, T.A.; Albanes, D.; Berndt, S.I.; Brenner, H.; Caan, B.J.; Campbell, P.T.; Cao, Y.; Chang-Claude, J.; Chan, A.; et al. Meta-analysis of 16 Studies of the Association of Alcohol with Colorectal Cancer. Int. J. Cancer 2020, 146, 861–873. [Google Scholar] [CrossRef]
- Zheng, X.; Hur, J.; Nguyen, L.H.; Liu, J.; Song, M.; Wu, K.; Smith-Warner, S.A.; Ogino, S.; Willett, W.C.; Chan, A.T.; et al. Comprehensive Assessment of Diet Quality and Risk of Precursors of Early-Onset Colorectal Cancer. J. Natl. Cancer Inst. 2021, 113, 543–552. [Google Scholar] [CrossRef]
- de Rezende, L.F.M.; de Sá, T.H.; Markozannes, G.; Rey-López, J.P.; Lee, I.M.; Tsilidis, K.K.; Ioannidis, J.P.; Eluf-Neto, J. Physical Activity and Cancer: An Umbrella Review of the Literature Including 22 Major Anatomical Sites and 770,000 Cancer Cases. Br. J. Sports Med. 2018, 52, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Lichtenstein, P.; Holm, N.V.; Verkasalo, P.K.; Iliadou, A.; Kaprio, J.; Koskenvuo, M.; Pukkala, E.; Skytthe, A.; Hemminki, K. Environmental and Heritable Factors in the Causation of Cancer—Analyses of Cohorts of Twins from Sweden, Denmark, and Finland. N. Engl. J. Med. 2000, 343, 78–85. [Google Scholar] [CrossRef]
- Graff, R.E.; Möller, S.; Passarelli, M.N.; Witte, J.S.; Skytthe, A.; Christensen, K.; Tan, Q.; Adami, H.O.; Czene, K.; Harris, J.R.; et al. Familial Risk and Heritability of Colorectal Cancer in the Nordic Twin Study of Cancer. Clin. Gastroenterol. Hepatol. 2017, 15, 1256–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasperson, K.W.; Tuohy, T.M.; Neklason, D.W.; Burt, R.W. Hereditary and Familial Colon Cancer. Gastroenterology 2010, 138, 2044–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.H.; Yu, J. Gut Microbiota in Colorectal Cancer: Mechanisms of Action and Clinical Applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.; de Martel, C.; Vignat, J.; Ferlay, J.; Bray, F.; Franceschi, S. Global Burden of Cancers Attributable to Infections in 2012: A Synthetic Analysis. Lancet Glob. Health 2016, 4, e609–e616. [Google Scholar] [CrossRef] [Green Version]
- Hughes, L.A.; van den Brandt, P.A.; Goldbohm, R.A.; de Goeij, A.F.; de Bruïne, A.P.; van Engeland, M.; Weijenberg, M.P. Childhood and Adolescent Energy Restriction and Subsequent Colorectal Cancer Risk: Results from the Netherlands Cohort Study. Int. J. Epidemiol. 2010, 39, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Nimptsch, K.; Wu, K. Is Timing Important? The Role of Diet and Lifestyle during Early Life on Colorectal Neoplasia. Curr. Colorectal Cancer Rep. 2018, 14, 1–11. [Google Scholar] [CrossRef]
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the Human Microbiome. Nutr. Rev. 2012, 70, S38–S44. [Google Scholar] [CrossRef] [Green Version]
- Bull, M.J.; Plummer, N.T. Part 1: The Human Gut Microbiome in Health and Disease. Integr. Med. 2014, 13, 17–22. [Google Scholar]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Gao, Z.; Huang, L.; Qin, H. Gut Microbiota and Colorectal Cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 757–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Louca, P.; Gibson, R.; Menni, C.; Spector, T.D.; le Roy, C.I. The Complexities of the Diet-Microbiome Relationship: Advances and Perspectives. Genome Med. 2021, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth Cells Directly Sense Gut Commensals and Maintain Homeostasis at the Intestinal Host-Microbial Interface. Proc. Natl. Acad. Sci. USA 2008, 105, 20858–20863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkaid, Y.; Naik, S. Compartmentalized and Systemic Control of Tissue Immunity by Commensals. Nat. Immunol. 2013, 14, 646–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnúsdóttir, S.; Ravcheev, D.; de Crécy-Lagard, V.; Thiele, I. Systematic Genome Assessment of B-Vitamin Biosynthesis Suggests Co-Operation among Gut Microbes. Front. Genet. 2015, 6, 148. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A.; Segre, J.A. The Human Microbiome: Our Second Genome. Annu. Rev. Genomics Hum. Genet. 2012, 13, 151–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geva-Zatorsky, N.; Sefik, E.; Kua, L.; Pasman, L.; Tan, T.G.; Ortiz-Lopez, A.; Yanortsang, T.B.; Yang, L.; Jupp, R.; Mathis, D.; et al. Mining the Human Gut Microbiota for Immunomodulatory Organisms. Cell 2017, 168, 928–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haber, A.L.; Biton, M.; Rogel, N.; Herbst, R.H.; Shekhar, K.; Smillie, C.; Burgin, G.; Delorey, T.M.; Howitt, M.R.; Katz, Y.; et al. A Single-Cell Survey of the Small Intestinal Epithelium. Nature 2017, 551, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Korem, T.; Zeevi, D.; Suez, J.; Weinberger, A.; Avnit-Sagi, T.; Pompan-Lotan, M.; Matot, E.; Jona, G.; Harmelin, A.; Cohen, N.; et al. Growth Dynamics of Gut Microbiota in Health and Disease Inferred from Single Metagenomic Samples. Science (1979) 2015, 349, 1101–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Shindo, Y.; Hazama, S. Novel Biomarkers for Personalized Cancer Immunotherapy. Cancers 2019, 11, 1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picardo, S.L.; Coburn, B.; Hansen, A.R. The Microbiome and Cancer for Clinicians. Crit. Rev. Oncol. Hematol. 2019, 141, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I.D.; Nicholson, J.K. Gut Microbiome Interactions with Drug Metabolism, Efficacy, and Toxicity. Transl. Res. 2017, 179, 204–222. [Google Scholar] [CrossRef] [Green Version]
- Brenchley, J.M.; Douek, D.C. Microbial Translocation across the GI Tract. Annu. Rev. Immunol. 2012, 30, 149. [Google Scholar] [CrossRef] [Green Version]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Núñez, G. Gut Microbiota-Induced Immunoglobulin G Controls Systemic Infection by Symbiotic Bacteria and Pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Knight, R. Dietary Effects on Human Gut Microbiome Diversity. Br. J. Nutr. 2015, 113, S1–S5. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-Induced Extinctions in the Gut Microbiota Compound over Generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, A.P.; Redinbo, M.R.; Bultman, S.J. The Role of the Microbiome in Cancer Development and Therapy. CA Cancer J. Clin. 2017, 67, 326–344. [Google Scholar] [CrossRef] [Green Version]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host Lifestyle Affects Human Microbiota on Daily Timescales. Genome Biol. 2014, 15, R89. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of Diet in Shaping Gut Microbiota Revealed by a Comparative Study in Children from Europe and Rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z. Gut Microbiota: An Important Link between Western Diet and Chronic Diseases. Nutrients 2019, 11, 2287. [Google Scholar] [CrossRef] [Green Version]
- Brennan, C.A.; Garrett, W.S. Gut Microbiota, Inflammation, and Colorectal Cancer. Annu. Rev. Microbiol. 2016, 70, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.H.R.; Bakir, I.A.; Hart, A.L.; Graham, T.A. Clonal Evolution of Colorectal Cancer in IBD. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Lasry, A.; Zinger, A.; Ben-Neriah, Y. Inflammatory Networks Underlying Colorectal Cancer. Nat. Immunol. 2016, 17, 230–240. [Google Scholar] [CrossRef]
- Alam, W.; Ullah, H.; Santarcangelo, C.; di Minno, A.; Khan, H.; Daglia, M.; Arciola, C.R. Micronutrient Food Supplements in Patients with Gastro-Intestinal and Hepatic Cancers. Int. J. Mol. Sci. 2021, 22, 8014. [Google Scholar] [CrossRef]
- Kim, D.H.; Khan, H.; Ullah, H.; Hassan, S.T.S.; Šmejkal, K.; Efferth, T.; Mahomoodally, M.F.; Xu, S.; Habtemariam, S.; Filosa, R.; et al. MicroRNA Targeting by Quercetin in Cancer Treatment and Chemoprotection. Pharmacol. Res. 2019, 147, 104346. [Google Scholar] [CrossRef]
- Shankar, E.; Kanwal, R.; Candamo, M.; Gupta, S. Dietary Phytochemicals as Epigenetic Modifiers in Cancer: Promise and Challenges. Semin. Cancer Biol. 2016, 40, 82–99. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.; Ullah, H.; Martorell, M.; Valdes, S.E.; Belwal, T.; Tejada, S.; Sureda, A.; Kamal, M.A. Flavonoids Nanoparticles in Cancer: Treatment, Prevention and Clinical Prospects. Semin. Cancer Biol. 2021, 69, 200–211. [Google Scholar] [CrossRef]
- Khan, H.; Reale, M.; Ullah, H.; Sureda, A.; Tejada, S.; Wang, Y.; Zhang, Z.J.; Xiao, J. Anti-Cancer Effects of Polyphenols via Targeting P53 Signaling Pathway: Updates and Future Directions. Biotechnol. Adv. 2020, 38, 107385. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Banerjee, S.; Mondal, A.; Chakraborty, U.; Pumarol, J.; Croley, C.R.; Bishayee, A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells 2020, 9, 1451. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural Products Targeting the PI3K-Akt-MTOR Signaling Pathway in Cancer: A Novel Therapeutic Strategy. Semin. Cancer Biol. 2019, 80, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Moloudizargari, M.; Asghari, M.H.; Nabavi, S.F.; Gulei, D.; Berindan-Neagoe, I.; Bishayee, A.; Nabavi, S.M. Targeting Hippo Signaling Pathway by Phytochemicals in Cancer Therapy. Semin. Cancer Biol. 2020, 80, 183–194. [Google Scholar] [CrossRef]
- Tewari, D.; Priya, A.; Bishayee, A.; Bishayee, A. Targeting Transforming Growth Factor-β Signalling for Cancer Prevention and Intervention: Recent Advances in Developing Small Molecules of Natural Origin. Clin. Transl. Med. 2022, 12, e795. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.; Mishra, S.R.; Behera, B.P.; Mahapatra, K.K.; Panigrahi, D.P.; Bhol, C.S.; Praharaj, P.P.; Sethi, G.; Patra, S.K.; Bhutia, S.K. Autophagy-Modulating Phytochemicals in Cancer Therapeutics: Current Evidences and Future Perspectives. Semin. Cancer Biol. 2020, 80, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Ullah, H.; Castilho, P.C.M.F.; Gomila, A.S.; D’Onofrio, G.; Filosa, R.; Wang, F.; Nabavi, S.M.; Daglia, M.; Silva, A.S.; et al. Targeting NF-ΚB Signaling Pathway in Cancer by Dietary Polyphenols. Crit. Rev. Food Sci. Nutr. 2019, 60, 2790–2800. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhou, Y.; Li, Y.; Xu, D.P.; Li, S.; Li, H.B. Spices for Prevention and Treatment of Cancers. Nutrients 2016, 8, 495. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, H.; Fan, D.; Deng, J. The Anticancer Activity and Mechanisms of Ginsenosides: An Updated Review. eFood 2020, 1, 226–241. [Google Scholar] [CrossRef]
- Jing, H. Black Garlic: Processing, Composition Change, and Bioactivity. eFood 2020, 1, 242–246. [Google Scholar] [CrossRef]
- Ekor, M. The Growing Use of Herbal Medicines: Issues Relating to Adverse Reactions and Challenges in Monitoring Safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Urbi, Z. Effect of Naphthalene Acetic Acid on the Adventitious Rooting in Shoot Cuttings of Andrographis paniculata (Burm. f.) Wall. Ex Nees: An Important Therapeutical Herb. Int. J. Agron. 2016, 2016, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.S.; Sharfaraz, A.; Dutta, A.; Ahsan, A.; Masud, M.A.; Ahmed, I.A.; Goh, B.H.; Urbi, Z.; Sarker, M.M.R.; Ming, L.C. A Review of Ethnobotany, Phytochemistry, Antimicrobial Pharmacology and Toxicology of Nigella sativa L. Biomed. Pharmacother. 2021, 143, 112182. [Google Scholar] [CrossRef]
- DeLuca, J.A.; Garcia-Villatoro, E.L.; Allred, C.D. Flaxseed Bioactive Compounds and Colorectal Cancer Prevention. Curr. Oncol. Rep. 2018, 20, 59. [Google Scholar] [CrossRef]
- Jaksevicius, A.; Carew, M.; Mistry, C.; Modjtahedi, H.; Opara, E.I. Inhibitory Effects of Culinary Herbs and Spices on the Growth of HCA-7 Colorectal Cancer Cells and Their COX-2 Expression. Nutrients 2017, 9, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanankutty, A. PI3K/Akt/MTOR Pathway as a Therapeutic Target for Colorectal Cancer: A Review of Preclinical and Clinical Evidence. Curr. Drug Targets 2019, 20, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Xin, L.; Liang, A.; Fu, Y. Cancer Stem Cell Hypothesis: A Brief Summary and Two Proposals. Cytotechnology 2013, 65, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, J.L.; Scott, A.J.; Pouncey, A.L.; Marchesi, J.; Kinross, J.; Teare, J. Colorectal Carcinogenesis: An Archetype of Gut Microbiota–Host Interaction. Ecancermedicalscience 2018, 12, 865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpa, E.S.; Ninfali, P. Phytochemicals as Innovative Therapeutic Tools against Cancer Stem Cells. Int. J. Mol. Sci. 2015, 16, 15727–15742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koury, J.; Zhong, L.; Hao, J. Targeting Signaling Pathways in Cancer Stem Cells for Cancer Treatment. Stem Cells Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.S.; Kader, M.A.; Goh, K.W.; Islam, M.; Khan, M.S.; Harun-Ar, M.R.; Ooi, J.; Melo, H.C.; Al-Worafi, Y.M.; Moshawih, S.; et al. Herb and Spices in Colorectal Cancer Prevention and Treatment: A Narrative Review. Front. Pharmacol. 2022, 13, 865801. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Jayachandran, M.; Xu, B. Diet-Derived Phytochemicals Targeting Colon Cancer Stem Cells and Microbiota in Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 3976. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the Normal Gut Microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wu, L.; Huntington, N.D.; Zhang, X. Crosstalk between Gut Microbiota and Innate Immunity and Its Implication in Autoimmune Diseases. Front. Immunol. 2020, 11, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarner, F.; Malagelada, J.R. Gut Flora in Health and Disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Jeyamogan, S.; Khan, N.A.; Anwar, A.; Shah, M.R.; Siddiqui, R. Cytotoxic Effects of Benzodioxane, Naphthalene Diimide, Porphyrin and Acetamol Derivatives on HeLa Cells. SAGE Open Med. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Jeyamogan, S.; Khan, N.A.; Siddiqui, R. Application and Importance of Theranostics in the Diagnosis and Treatment of Cancer. Arch. Med. Res. 2021, 52, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. Emerging Roles of the Microbiome in Cancer. Carcinogenesis 2014, 35, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, B. New Therapeutic Targets for Cancer: The Interplay between Immune and Metabolic Checkpoints and Gut Microbiota. Clin. Transl. Med. 2019, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLoS One 2012, 7, e39743. [Google Scholar] [CrossRef]
- Wu, N.; Yang, X.; Zhang, R.; Li, J.; Xiao, X.; Hu, Y.; Chen, Y.; Yang, F.; Lu, N.; Wang, Z.; et al. Dysbiosis Signature of Fecal Microbiota in Colorectal Cancer Patients. Microb. Ecol. 2013, 66, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, J.; Rai, R.P.; Prasad, K.N. Role of Helicobacter pylori in Gastric Cancer: Updates. World J. Gastroenterol. 2016, 8, 147. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N.; Ushijima, T. Epigenetic Impact of Infection on Carcinogenesis: Mechanisms and Applications. Genome Med. 2016, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Arthur, J.C.; Perez-Chanona, E.; Mühlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal Inflammation Targets Cancer-Inducing Activity of the Microbiota. Science (1979) 2012, 338, 120–123. [Google Scholar] [CrossRef] [Green Version]
- Nougayrède, J.P.; Homburg, S.; Taieb, F.; Boury, M.; Brzuszkiewicz, E.; Gottschalk, G.; Buchrieser, C.; Hacker, J.; Dobrindt, U.; Oswald, E. Escherichia coli Induces DNA Double-Strand Breaks in Eukaryotic Cells. Science (1979) 2006, 313, 848–851. [Google Scholar] [CrossRef]
- Dennis, K.L.; Blatner, N.R.; Gounari, F.; Gounari, F. Current Status of Interleukin-10 and Regulatory T-Cells in Cancer. Curr. Opin. Oncol. 2013, 25, 637–645. [Google Scholar] [CrossRef]
- Al-Hebshi, N.N.; Borgnakke, W.S.; Johnson, N.W. The Microbiome of Oral Squamous Cell Carcinomas: A Functional Perspective. Curr. Oral Health Rep. 2019, 6, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Vivarelli, S.; Salemi, R.; Candido, S.; Falzone, L.; Santagati, M.; Stefani, S.; Torino, F.; Banna, G.L.; Tonini, G.; Libra, M. Gut Microbiota and Cancer: From Pathogenesis to Therapy. Cancers (Basel) 2019, 11, 38. [Google Scholar] [CrossRef] [Green Version]
- Borges-Canha, M.; Portela-Cidade, J.P.; Dinis-Ribeiro, M.; Leite-Moreira, A.F.; Pimentel-Nunes, P. Role of Colonic Microbiota in Colorectal Carcinogenesis: A Systematic Review. Rev. Esp. Enferm. Dig. 2015, 107, 659–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klampfer, L. Cytokines, Inflammation and Colon Cancer. Curr. Cancer Drug Targets 2011, 11, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Bergounioux, J.; Elisee, R.; Prunier, A.L.; Donnadieu, F.; Sperandio, B.; Sansonetti, P.; Arbibe, L. Calpain Activation by the Shigella flexneri Effector VirA Regulates Key Steps in the Formation and Life of the Bacterium’s Epithelial Niche. Cell Host Microbe 2012, 11, 240–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via Its FadA Adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.Y.; Wu, C.Y.; Yu, J. The Role of Gut Microbiota in Cancer Treatment: Friend or Foe? Gut 2013, 69, 1867–1876. [Google Scholar] [CrossRef]
- Goodwin, A.C.; Shields, C.E.D.; Wu, S.; Huso, D.L.; Wu, X.; Murray-Stewart, T.R.; Hacker-Prietz, A.; Rabizadeh, S.; Woster, P.M.; Sears, C.L.; et al. Polyamine Catabolism Contributes to Enterotoxigenic Bacteroides fragilis-Induced Colon Tumorigenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 15354–15359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, H.; Fujiya, M.; Tanaka, H.; Ueno, N.; Moriichi, K.; Sasajima, J.; Ikuta, K.; Akutsu, H.; Tanabe, H.; Kohgo, Y. Probiotic-Derived Ferrichrome Inhibits Colon Cancer Progression via JNK-Mediated Apoptosis. Nat. Commun. 2016, 7, 12365. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.; Jakobsson, H.E.; Holmén-Larsson, J.; Schütte, A.; Ermund, A.; Rodríguez-Piñeiro, A.M.; Arike, L.; Wising, C.; Svensson, F.; Bäckhed, F.; et al. Normalization of Host Intestinal Mucus Layers Requires Long-Term Microbial Colonization. Cell Host Microbe 2015, 18, 582–592. [Google Scholar] [CrossRef] [Green Version]
- Spiljar, M.; Merkler, D.; Trajkovski, M. The Immune System Bridges the Gut Microbiota with Systemic Energy Homeostasis: Focus on TLRs, Mucosal Barrier, and SCFAs. Front. Immunol. 2017, 8, 1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Rao, Y.; Guo, X.; Liu, N.; Liu, S.; Wen, P.; Li, S.; Li, Y. Oral Microbiome in Patients with Oesophageal Squamous Cell Carcinoma. Sci. Rep. 2019, 9, 19055. [Google Scholar] [CrossRef] [Green Version]
- Nair, M.; Sandhu, S.S.; Sharma, A.K. Cancer Molecular Markers: A Guide to Cancer Detection and Management. Semin. Cancer Biol. 2018, 52, 39–55. [Google Scholar] [CrossRef]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug Resistance in Cancer: An Overview. Cancers (Basel) 2014, 6, 1769–1792. [Google Scholar] [CrossRef]
- Rueff, J.; Rodrigues, A.S. Cancer Drug Resistance: A Brief Overview from a Genetic Viewpoint. Methods Mol. Biol. 2016, 1395, 1–18. [Google Scholar] [PubMed]
- Alfarouk, K.O.; Stock, C.-M.; Taylor, S.; Walsh, M.; Muddathir, A.K.; Verduzco, D.; Bashir, A.H.H.; Mohammed, O.Y.; Elhassan, G.O.; Harguindey, S.; et al. Resistance to Cancer Chemotherapy: Failure in Drug Response from ADME to P-Gp. Cancer Cell Int. 2015, 15, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwabe, R.F.; Jobin, C. The Microbiome and Cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zitvogel, L.; Daillère, R.; Roberti, M.P.; Routy, B.; Kroemer, G. Anticancer Effects of the Microbiome and Its Products. Nat. Rev. Microbiol. 2017, 15, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Iida, N.; Dzutsev, A.; Stewart, C.; Smith, L.; Bouladoux, N.; Weingarten, R.; Molina, D.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal Bacteria Control Cancer Response to Therapy by Modulating the Tumor Microenvironment. Science (1979) 2013, 342, 967–970. [Google Scholar] [CrossRef]
- Kuwahara, A.; Matsuda, K.; Kuwahara, Y.; Asano, S.; Inui, T.; Marunaka, Y. Microbiota-Gut-Brain Axis: Enteroendocrine Cells and the Enteric Nervous System Form an Interface between the Microbiota and the Central Nervous System. Biomed. Res. 2020, 41, 199–216. [Google Scholar] [CrossRef]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.; Pfirschke, C.; Engblom, C.; Pittet, M.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science (1979) 2013, 342, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Man Lei, Y.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium Promotes Antitumor Immunity and Facilitates Anti–PD-L1 Efficacy. Science (1979) 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Cai, X.; Zhang, J.; Wang, W.; Sheng, Q.; Hua, H.; Zhou, X. Role of Gut Microbiota in the Development and Treatment of Colorectal Cancer. Digestion 2019, 100, 72–78. [Google Scholar] [CrossRef]
- Vétizou, M.; Pitt, J.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.; Duong, C.; et al. Anticancer Immunotherapy by CTLA-4 Blockade Relies on the Gut Microbiota. Science (1979) 2015, 350, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Jan, G.B.A.S.; Belzacq, A.S.; Haouzi, D.; Rouault, A.; Metivier, D.; Kroemer, G.; Brenner, C. Propionibacteria Induce Apoptosis of Colorectal Carcinoma Cells via Short-Chain Fatty Acids Acting on Mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Sun, W.; Yu, S.; Yang, Y.; Ai, L. Butyrate Production from High-Fiber Diet Protects against Lymphoma Tumor. Leuk. Lymphoma 2016, 57, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xia, Y.; Sun, J. Breast and Gut Microbiome in Health and Cancer. Genes Dis. 2021, 8, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alcoholado, L.; Ramos-Molina, B.; Otero, A.; Laborda-Illanes, A.; Ordóñez, R.; Medina, J.A.; Gómez-Millán, J.; Queipo-Ortuño, M.I. The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response. Cancers (Basel) 2020, 12, 1406. [Google Scholar] [CrossRef]
- Salcedo, R.; Worschech, A.; Cardone, M.; Jones, Y.; Gyulai, Z.; Dai, R.M.; Wang, E.; Ma, W.; Haines, D.; O’hUigin, C.; et al. MyD88-Mediated Signaling Prevents Development of Adenocarcinomas of the Colon: Role of Interleukin 18. J. Exp. Med. 2010, 207, 1625–1636. [Google Scholar] [CrossRef]
- Paavonen, J.; Naud, P.; Salmerón, J.; Wheeler, C.; Chow, S.; Apter, D.; Kitchener, H.; Castellsague, X.; Teixeira, J.; Skinner, S.; et al. Efficacy of Human Papillomavirus (HPV)-16/18 AS04-Adjuvanted Vaccine against Cervical Infection and Precancer Caused by Oncogenic HPV Types (PATRICIA): Final Analysis of a Double-Blind, Randomised Study in Young Women. Lancet 2009, 374, 301–314. [Google Scholar] [CrossRef]
- Shylaja, M.R.; Peter, K.V. Spices in the Nutraceutical and Health Food Industry. In International Symposium on Medicinal and Nutraceutical Plants; International Society for Horticultural Science: Macon, GA, USA, 2007; pp. 369–378. [Google Scholar]
- Srinivasan, K. Role of Spices beyond Food Flavoring: Nutraceuticals with Multiple Health Effects. Food Rev. Int. 2005, 21, 167–188. [Google Scholar] [CrossRef]
- Kaefer, C.M.; Milner, J.A. The Role of Herbs and Spices in Cancer Prevention. J. Nutr. Biochem. 2008, 19, 347–361. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Ji, H.F. Intestinal Microbiota and Metabolic Diseases: Pharmacological Implications. Trends Pharmacol. Sci. 2016, 37, 169–171. [Google Scholar] [CrossRef]
- Shen, L.; Liu, L.; Ji, H.F. Alzheimer’s Disease Histological and Behavioral Manifestations in Transgenic Mice Correlate with Specific Gut Microbiome State. J. Alzheimer’s Dis. 2017, 56, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T.; Vaughn, A.R.; Sharma, V.; Chopra, D.; Mills, P.J.; Peterson, S.N.; Sivamani, R.K. Effects of Turmeric and Curcumin Dietary Supplementation on Human Gut Microbiota: A Double-Blind, Randomized, Placebo-Controlled Pilot Study. J. Evid. Based Integr. Med. 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Nishida, A.; Sugitani, Y.; Nishino, K.; Inatomi, O.; Sugimoto, M.; Kawahara, M.; Andoh, A. Nanoparticle Curcumin Ameliorates Experimental Colitis via Modulation of Gut Microbiota and Induction of Regulatory T Cells. PLoS One 2017, 12, e0185999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, R.M.T.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.A.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C.; et al. The Role of Curcumin in Modulating Colonic Microbiota during Colitis and Colon Cancer Prevention. Inflamm. Bowel Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Muñoz, M.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Loddenkemper, C.; Göbel, U.B.; Heimesaat, M.M. Anti-Inflammatory Effects of Resveratrol, Curcumin and Simvastatin in Acute Small Intestinal Inflammation. PLoS One 2010, 5, e15099. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Geng, W.; Chen, S.; Wang, L.; Rong, X.; Wang, S.; Wang, T.; Xiong, L.; Huang, J.; Pang, X.; et al. Ginger Alleviates DSS-Induced Ulcerative Colitis Severity by Improving the Diversity and Function of Gut Microbiota. Front. Pharmacol. 2021, 12, 632569. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, X.; He, Q.; Wang, M.; Lu, H.; You, Y.; Chen, L.; Cheng, J.; Li, F.; Fu, X.; et al. Ginger Extract Decreases Susceptibility to Dextran Sulfate Sodium-Induced Colitis in Mice Following Early Antibiotic Exposure. Front. Med. 2022, 8. [Google Scholar]
- Sasaki, K.; Sasaki, D.; Sasaki, K.; Nishidono, Y.; Yamamori, A.; Tanaka, K.; Kondo, A. Growth Stimulation of Bifidobacterium from Human Colon Using Daikenchuto in an In Vitro Model of Human Intestinal Microbiota. Sci. Rep. 2021, 22, 4580. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Chen, Z.; Yuan, Q.; Ma, M.; Gao, C.; Zhou, Y.; Zhou, H.; Wu, X.; Wu, D.; Farag, M.A.; et al. Ginger Polysaccharides Relieve Ulcerative Colitis via Maintaining Intestinal Barrier Integrity and Gut Microbiota Modulation. Int. J. Biol. Macromol. 2022, 219, 730–739. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, D.; Jiang, H.; Zhang, S.; Pang, X.; Gao, S.; Zhang, H.; Zhang, S.; Xiao, Q.; Chen, L.; et al. Gut Microbiota Variation with Short-Term Intake of Ginger Juice on Human Health. Front. Microbiol. 2021, 11, 576061. [Google Scholar] [CrossRef]
- Shao, X.; Sun, C.; Tang, X.; Zhang, X.; Han, D.; Liang, S.; Qu, R.; Hui, X.; Shan, Y.; Hu, L.; et al. Anti-Inflammatory and Intestinal Microbiota Modulation Properties of Jinxiang Garlic (Allium sativum L.) Polysaccharides toward Dextran Sodium Sulfate-Induced Colitis. J. Agric. Food Chem. 2020, 68, 12295–12309. [Google Scholar] [CrossRef] [PubMed]
- Vezza, T.; Algieri, F.; Garrido-Mesa, J.; Utrilla, M.P.; Rodríguez-Cabezas, M.E.; Banos, A.; Guillamón, E.; García, F.; Rodríguez-Nogales, A.; Gálvez, J. The Immunomodulatory Properties of Propyl-propane Thiosulfonate Contribute to Its Intestinal Anti-inflammatory Effect in Experimental Colitis. Mol. Nutr. Food Res. 2019, 63, 1800653. [Google Scholar] [CrossRef]
- Hussein, M.M.; Abd El-Hack, M.E.; Mahgoub, S.A.; Saadeldin, I.M.; Swelum, A.A. Effects of Clove (Syzygium aromaticum) Oil on Quail Growth, Carcass Traits, Blood Components, Meat Quality, and Intestinal Microbiota. Poult. Sci. 2019, 98, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Zhang, Y.; Zhu, X.; Liu, K.; Wang, X.; Chen, M.; Wang, J.; Chen, H.; Hui, S.; Huang, L.; et al. Healthy Subjects Differentially Respond to Dietary Capsaicin Correlating with Specific Gut Enterotypes. J. Clin. Endocrinol. Metab. 2016, 101, 4681–4689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Xiao, Q.; Xiong, Z.; Yu, C.; Zhou, J.; Fu, Z. Crocin-I Ameliorates the Disruption of Lipid Metabolism and Dysbiosis of the Gut Microbiota Induced by Chronic Corticosterone in Mice. Food Funct. 2019, 10, 6779–6791. [Google Scholar] [CrossRef]
- Feng, P.; Li, Q.; Liu, L.; Wang, S.; Wu, Z.; Tao, Y.; Huang, P.; Wang, P. Crocetin Prolongs Recovery Period of DSS-Induced Colitis via Altering Intestinal Microbiome and Increasing Intestinal Permeability. Int. J. Mol. Sci. 2022, 23, 3832. [Google Scholar] [CrossRef]
- Agarwal, N.; Kolba, N.; Jung, Y.; Cheng, J.; Tako, E. Saffron (Crocus sativus L.) Flower Water Extract Disrupts the Cecal Microbiome, Brush Border Membrane Functionality, and Morphology in Vivo (Gallus gallus). Nutrients 2022, 14, 220. [Google Scholar] [CrossRef]
- Power, K.A.; Lepp, D.; Zarepoor, L.; Monk, J.M.; Wu, W.; Tsao, R.; Liu, R. Dietary Flaxseed Modulates the Colonic Microenvironment in Healthy C57Bl/6 Male Mice Which May Alter Susceptibility to Gut-Associated Diseases. J. Nutr. Biochem. 2016, 28, 61–69. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, W.; Deng, Q.; Huang, Q.; Wang, X.; Yang, C.; Huang, F. Flaxseed Oligosaccharides Alleviate DSS-Induced Colitis through Modulation of Gut Microbiota and Repair of the Intestinal Barrier in Mice. Food Funct. 2020, 11, 8077–8088. [Google Scholar] [CrossRef]
- Che, L.; Zhou, Q.; Liu, Y.; Hu, L.; Peng, X.; Wu, C.; Zhang, R.; Tang, J.; Wu, F.; Fang, Z.; et al. Flaxseed Oil Supplementation Improves Intestinal Function and Immunity, Associated with Altered Intestinal Microbiome and Fatty Acid Profile in Pigs with Intrauterine Growth Retardation. Food Funct. 2019, 10, 8149–8160. [Google Scholar] [CrossRef]
- Plissonneau, C.; Sivignon, A.; Chassaing, B.; Capel, F.; Martin, V.; Etienne, M.; Wawrzyniak, I.; Chausse, P.; Dutheil, F.; Mairesse, G.; et al. Beneficial Effects of Linseed Supplementation on Gut Mucosa-Associated Microbiota in a Physically Active Mouse Model of Crohn’s Disease. Int. J. Mol. Sci. 2022, 23, 5891. [Google Scholar] [CrossRef]
- Bachmeier, B.E.; Killian, P.H.; Melchart, D. The Role of Curcumin in Prevention and Management of Metastatic Disease. Int. J. Mol. Sci. 2018, 19, 1716. [Google Scholar] [CrossRef]
- Mirzaei, H.; Bagheri, H.; Ghasemi, F.; Khoi, J.M.; Pourhanifeh, M.H.; Heyden, Y.V.; Mortezapour, E.; Nikdasti, A.; Jeandet, P.; Khan, H.; et al. Anti-Cancer Activity of Curcumin on Multiple Myeloma. Anticancer Agents Med. Chem. 2021, 21, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Duvoix, A.; Blasius, R.; Delhalle, S.; Schnekenburger, M.; Morceau, F.; Henry, E.; Dicato, M.; Diederich, M. Chemopreventive and Therapeutic Effects of Curcumin. Cancer Lett. 2005, 223, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.C.; Kamarudin, M.N.A.; Naidu, R. Anticancer Mechanism of Curcumin on Human Glioblastoma. Nutrients 2021, 13, 950. [Google Scholar] [CrossRef] [PubMed]
- Genua, F.; Raghunathan, V.; Jenab, M.; Gallagher, W.M.; Hughes, D.J. The Role of Gut Barrier Dysfunction and Microbiome Dysbiosis in Colorectal Cancer Development. Front. Oncol. 2021, 11, 6349. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ghosh, S.S.; Ghosh, S. Curcumin Improves Intestinal Barrier Function: Modulation of Intracellular Signaling, and Organization of Tight Junctions. Am. J. Physiol. Cell Physiol. 2017, 312, C438–C445. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Bie, J.; Wang, J.; Ghosh, S. Oral Supplementation with Non-Absorbable Antibiotics or Curcumin Attenuates Western Diet-Induced Atherosclerosis and Glucose Intolerance in LDLR−/− Mice–Role of Intestinal Permeability and Macrophage Activation. PLoS One 2014, 9, e108577. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.S.; He, H.; Wang, J.; Gehr, T.W.; Ghosh, S. Curcumin-Mediated Regulation of Intestinal Barrier Function: The Mechanism Underlying Its Beneficial Effects. Tissue Barriers 2018, 6, e1425085. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Su, C.; Feng, H.; Chen, X.; Dong, Y.; Rao, Y.; Ren, Y.; Yang, J.; Shi, J.; Tian, J.; et al. Curcumin Regulates Insulin Pathways and Glucose Metabolism in the Brains of APPswe/PS1dE9 Mice. Int. J. Immunopathol. Pharmacol. 2017, 30, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Scazzocchio, B.; Minghetti, L.; D’Archivio, M. Interaction between Gut Microbiota and Curcumin: A New Key of Understanding for the Health Effects of Curcumin. Nutrients 2020, 12, 2499. [Google Scholar] [CrossRef]
- Shen, L.; Ji, H.F. Bidirectional Interactions between Dietary Curcumin and Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 2896–2902. [Google Scholar] [CrossRef]
- Zhai, S.S.; Ruan, D.; Zhu, Y.W.; Li, M.C.; Ye, H.; Wang, W.C.; Yang, L. Protective Effect of Curcumin on Ochratoxin A–Induced Liver Oxidative Injury in Duck Is Mediated by Modulating Lipid Metabolism and the Intestinal Microbiota. Poult. Sci. 2020, 99, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, L.; Ji, H.F. Regulative Effects of Curcumin Spice Administration on Gut Microbiota and Its Pharmacological Implications. Food Nutr. Res. 2017, 61, 1361780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, W.; Wang, H.; Zhang, P.; Gao, C.; Tao, J.; Ge, Z.; Zhu, D.; Bi, Y. Modulation of Gut Microbiota Contributes to Curcumin-Mediated Attenuation of Hepatic Steatosis in Rats. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Greiner, A.K.; Papineni, R.V.; Umar, S. Chemoprevention in Gastrointestinal Physiology and Disease. Natural Products and Microbiome. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G1–G15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, O.; Lahti, L.; Kokkola, A.; Karla, T.; Tikkanen, M.; Ehsan, H.; Carpelan-Holmström, M.; Koskensalo, S.; Böhling, T.; Rautelin, H.; et al. Stool Microbiota Composition Differs in Patients with Stomach, Colon, and Rectal Neoplasms. Dig. Dis. Sci. 2018, 63, 2950–2958. [Google Scholar] [CrossRef] [Green Version]
- Mori, G.; Orena, B.S.; Cultrera, I.; Barbieri, G.; Albertini, A.M.; Ranzani, G.N.; Carnevali, I.; Tibiletti, M.G.; Pasca, M.R. Gut Microbiota Analysis in Postoperative Lynch Syndrome Patients. Front. Microbiol. 2019, 10, 1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, P.; De, R.; Pal, I.; Mukhopadhyay, A.K.; Saha, D.R.; Swarnakar, S. Curcumin Alleviates Matrix Metalloproteinase-3 and-9 Activities during Eradication of Helicobacter pylori Infection in Cultured Cells and Mice. PLoS One 2011, 6, e16306. [Google Scholar] [CrossRef] [Green Version]
- Haghi, A.; Azimi, H.; Rahimi, R. A Comprehensive Review on Pharmacotherapeutics of Three Phytochemicals, Curcumin, Quercetin, and Allicin, in the Treatment of Gastric Cancer. J. Gastrointest. Cancer 2017, 48, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; di Minno, A.; Santarcangelo, C.; Khan, H.; Xiao, J.; Arciola, C.R.; Daglia, M. Vegetable Extracts and Nutrients Useful in the Recovery from Helicobacter pylori Infection: A Systematic Review on Clinical Trials. Molecules 2021, 26, 2272. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K. Ginger Rhizomes (Zingiber officinale): A Spice with Multiple Health Beneficial Potentials. PharmaNutrition 2017, 5, 18–28. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xu, X.Y.; Cao, S.Y.; Gan, R.Y.; Corke, H.; Beta, T.; Li, H.B. Bioactive Compounds and Bioactivities of Ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govindarajan, V.S.; Connell, D.W. Ginger—Chemistry, Technology, and Quality Evaluation: Part 1. Crit. Rev. Food Sci. Nutr. 1983, 17, 1–96. [Google Scholar] [CrossRef] [PubMed]
- Baliga, M.S.; Haniadka, R.; Pereira, M.M.; D’Souza, J.J.; Pallaty, P.L.; Bhat, H.P.; Popuri, S. Update on the Chemopreventive Effects of Ginger and Its Phytochemicals. Crit. Rev. Food Sci. Nutr. 2011, 51, 499–523. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tyagi, A.K. Ginger and Its Constituents: Role in Prevention and Treatment of Gastrointestinal Cancer. Gastroenterol. Res. Pract. 2015, 2015, 142979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Abhar, H.S.; Hammad, L.N.; Gawad, H.S.A. Modulating Effect of Ginger Extract on Rats with Ulcerative Colitis. J. Ethnopharmacol. 2008, 118, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, C.; Liu, D.; Han, M.K.; Wang, L.; Merlin, D. Oral Delivery of Nanoparticles Loaded with Ginger Active Compound, 6-Shogaol, Attenuates Ulcerative Colitis and Promotes Wound Healing in a Murine Model of Ulcerative Colitis. J. Crohns Colitis 2018, 12, 217–229. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M.K.; Xiao, B.; Xu, C.; Srinivasan, S.; et al. Edible Ginger-Derived Nanoparticles: A Novel Therapeutic Approach for the Prevention and Treatment of Inflammatory Bowel Disease and Colitis-Associated Cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, A.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Tang, G.Y.; Corke, H.; Mavumengwana, V.; Li, H.B. Bioactive Compounds and Biological Functions of Garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.A.; di Gioia, F.; Polyzos, N.; Tzortzakis, N. Natural Antioxidants, Health Effects and Bioactive Properties of Wild Allium Species. Curr. Pharm. Des. 2020, 26, 1816–1837. [Google Scholar] [CrossRef]
- Martins, N.; Petropoulos, S.; Ferreira, I.C. Chemical Composition and Bioactive Compounds of Garlic (Allium sativum L.) as Affected by Pre-and Post-Harvest Conditions: A Review. Food Chem. 2016, 211, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Miroddi, M.; Calapai, F.; Calapai, G. Potential Beneficial Effects of Garlic in Oncohematology. Mini Rev. Med. Chem. 2011, 11, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.; Ruan, J.; Zhuang, X.; Zhang, X.; Li, Z. Phytochemicals of Garlic: Promising Candidates for Cancer Therapy. Biomed. Pharmacother. 2020, 123, 109730. [Google Scholar] [CrossRef] [PubMed]
- Mondal, A.; Banerjee, S.; Bose, S.; Mazumder, S.; Haber, R.A.; Farzaei, M.H.; Bishayee, A. Garlic Constituents for Cancer Prevention and Therapy: From Phytochemistry to Novel Formulations. Pharmacol. Res. 2022, 175, 105837. [Google Scholar] [CrossRef] [PubMed]
- de Greef, D.; Barton, E.M.; Sandberg, E.N.; Croley, C.R.; Pumarol, J.; Wong, T.L.; Das, N.; Bishayee, A. Anticancer Potential of Garlic and Its Bioactive Constituents: A Systematic and Comprehensive Review. Semin. Cancer Biol. 2021, 73, 219–264. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Xie, K.; Liu, Z.; Nakasone, Y.; Sakao, K.; Hossain, M.A.; Hou, D.X. Preventive Effects and Mechanisms of Garlic on Dyslipidemia and Gut Microbiome Dysbiosis. Nutrients 2019, 11, 1225. [Google Scholar] [CrossRef] [Green Version]
- Ried, K.; Travica, N.; Sali, A. The Effect of Kyolic Aged Garlic Extract on Gut Microbiota, Inflammation, and Cardiovascular Markers in Hypertensives: The GarGIC Trial. Front. Nutr. 2018, 5, 122. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, F.; Huang, M.; Wu, H.; Yang, C.; Zhang, X.; Yang, L.; Chen, G.; Li, S.; Wang, Q.; et al. Fatty Liver and Alteration of the Gut Microbiome Induced by Diallyl Disulfide. Int. J. Mol. Med. 2019, 44, 1908–1920. [Google Scholar] [CrossRef] [PubMed]
- Kamatou, G.P.; Vermaak, I.; Viljoen, A.M. Eugenol—from the Remote Maluku Islands to the International Market Place: A Review of a Remarkable and Versatile Molecule. Molecules 2012, 17, 6953–6981. [Google Scholar] [CrossRef]
- Chaieb, K.; Hajlaoui, H.; Zmantar, T.; Kahla-Nakbi, A.B.; Rouabhia, M.; Mahdouani, K.; Bakhrouf, A. The Chemical Composition and Biological Activity of Clove Essential Oil, Eugenia Caryophyllata (Syzigium aromaticum L. Myrtaceae): A Short Review. Phytother. Res. 2007, 21, 501–506. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Neveu, V.; Vos, F.; Scalbert, A. Identification of the 100 Richest Dietary Sources of Polyphenols: An Application of the Phenol-Explorer Database. Eur. J. Clin. Nutr. 2010, 64, S112–S120. [Google Scholar] [CrossRef] [Green Version]
- Gülçin, İ.; Elmastaş, M.; Aboul-Enein, H.Y. Antioxidant Activity of Clove Oil–A Powerful Antioxidant Source. Arab. J. Chem. 2012, 5, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhao, G.; Zhang, D.; An, W.; Lai, H.; Li, X.; Cao, S.; Lin, X. Active Fraction of Clove Induces Apoptosis via PI3K/Akt/MTOR-Mediated Autophagy in Human Colorectal Cancer HCT-116 Cells. Int. J. Oncol. 2018, 53, 1363–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Schmitz, J.C.; Wei, J.; Cao, S.; Beumer, J.H.; Strychor, S.; Cheng, L.; Liu, M.; Wang, C.; Wu, N.; et al. Clove Extract Inhibits Tumor Growth and Promotes Cell Cycle Arrest and Apoptosis. Oncol. Res. 2014, 21, 247–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wu, X.; Tang, S.; Yin, J.; Song, Z.; He, X.; Yin, Y. Eugenol Alleviates Dextran Sulfate Sodium-Induced Colitis Independent of Intestinal Microbiota in Mice. J. Agric. Food Chem. 2021, 69, 10506–10514. [Google Scholar] [CrossRef] [PubMed]
- Saleh, B.K.; Omer, A.; Teweldemedhin, B. Medicinal Uses and Health Benefits of Chili Pepper (Capsicum Spp.): A Review. MOJ Food Process. Technol. 2018, 6, 325–328. [Google Scholar] [CrossRef]
- de Jong, P.R.; Takahashi, N.; Harris, A.R.; Lee, J.; Bertin, S.; Jeffries, J.; Jung, M.; Duong, J.; Triano, A.I.; Lee, J.; et al. Ion Channel TRPV1-Dependent Activation of PTP1B Suppresses EGFR-Associated Intestinal Tumorigenesis. J. Clin. Invest. 2014, 124, 3793–3806. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.C.; Millwood, I.Y.; Kartsonaki, C.; Du, H.; Guo, Y.; Chen, Y.; Bian, Z.; Walters, R.G.; Lv, J.; He, P.; et al. Spicy Food Consumption and Risk of Gastrointestinal-Tract Cancers: Findings from the China Kadoorie Biobank. Int. J. Epidemiol. 2021, 50, 199–211. [Google Scholar] [CrossRef]
- Hayashi, K.; Shibata, C.; Nagao, M.; Sato, M.; Kakyo, M.; Kinouchi, M.; Saijo, F.; Miura, K.; Ogawa, H.; Sasaki, I. Intracolonic Capsaicin Stimulates Colonic Motility and Defecation in Conscious Dogs. Surgery 2010, 147, 789–797. [Google Scholar] [CrossRef]
- Ashktorab, H.; Soleimani, A.; Singh, G.; Amin, A.; Tabtabaei, S.; Latella, G.; Stein, U.; Akhondzadeh, S.; Solanki, N.; Gondré-Lewis, M.C.; et al. Saffron: The Golden Spice with Therapeutic Properties on Digestive Diseases. Nutrients 2019, 11, 943. [Google Scholar] [CrossRef] [Green Version]
- Melnyk, J.P.; Wang, S.; Marcone, M.F. Chemical and Biological Properties of the World’s Most Expensive Spice: Saffron. Food Res. Int. 2010, 43, 1981–1989. [Google Scholar] [CrossRef]
- Xing, B.; Li, S.; Yang, J.; Lin, D.; Feng, Y.; Lu, J.; Shao, Q. Phytochemistry, Pharmacology, and Potential Clinical Applications of Saffron: A Review. J. Ethnopharmacol. 2021, 281, 114555. [Google Scholar] [CrossRef] [PubMed]
- Gezici, S. Comparative Anticancer Activity Analysis of Saffron Extracts and a Principle Component, Crocetin for Prevention and Treatment of Human Malignancies. J. Food Sci. Technol. 2019, 56, 5435–5443. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Tung, N.H.; Shoyama, Y.; Sugie, S.; Mori, T.; Tanaka, T. Dietary Crocin Inhibits Colitis and Colitis-Associated Colorectal Carcinogenesis in Male ICR Mice. Evid. base Compl. Alternative Med. 2012, 2012, 820415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amerizadeh, F.; Rezaei, N.; Rahmani, F.; Hassanian, S.M.; Moradi-Marjaneh, R.; Fiuji, H.; Boroumand, N.; Nosrati-Tirkani, A.; Ghayour-Mobarhan, M.; Ferns, G.A.; et al. Crocin Synergistically Enhances the Antiproliferative Activity of 5-flurouracil through Wnt/PI3K Pathway in a Mouse Model of Colitis-associated Colorectal Cancer. J. Cell. Biochem. 2018, 119, 10250–10261. [Google Scholar] [CrossRef] [PubMed]
- Güllü, N.; Kobelt, D.; Brim, H.; Rahman, S.; Timm, L.; Smith, J.; Soleimani, A.; di Marco, S.; Bisti, S.; Ashktorab, H.; et al. Saffron Crudes and Compounds Restrict MACC1-Dependent Cell Proliferation and Migration of Colorectal Cancer Cells. Cells 2020, 9, 1829. [Google Scholar] [CrossRef]
- Li, M.; Ding, L.; Hu, Y.L.; Qin, L.L.; Wu, Y.; Liu, W.; Wu, L.L.; Liu, T.H. Herbal Formula LLKL Ameliorates Hyperglycaemia, Modulates the Gut Microbiota and Regulates the Gut-liver Axis in Zucker Diabetic Fatty Rats. J. Cell. Mol. Med. 2021, 25, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Shim, Y.Y.; Gui, B.; Arnison, P.G.; Wang, Y.; Reaney, M.J. Flaxseed (Linum usitatissimum L.) Bioactive Compounds and Peptide Nomenclature: A Review. Trends Food Sci. Technol. 2014, 38, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, L.A.; dos Santos Ferreira, R.; de Cássia Avellaneda Guimarães, R.; de Castro, A.P.; Franco, O.L.; Matias, R.; Carvalho, C.M. The Complex Puzzle of Interactions among Functional Food, Gut Microbiota, and Colorectal Cancer. Front. Oncol. 2018, 8, 325. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, Z.; Huang, F.; Yang, C.; Huang, Q. In Vitro Digestion and Fermentation by Human Fecal Microbiota of Polysaccharides from Flaxseed. Molecules 2020, 25, 4354. [Google Scholar] [CrossRef] [PubMed]
- Määttänen, P.; Lurz, E.; Botts, S.R.; Wu, R.Y.; Yeung, C.W.; Li, B.; Abiff, S.; Johnson-Henry, K.C.; Lepp, D.; Power, K.A.; et al. Ground Flaxseed Reverses Protection of a Reduced-Fat Diet against Citrobacter rodentium-Induced Colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G788–G798. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Spice-Derived Compounds | In vivo Study Model | Dose | Treatment Duration | Effect on Gut Microbiota | Comments | References |
|---|---|---|---|---|---|---|
| Curcumin | Mice/Human | 100 mg/kg | 15 days | ↑Lactobacilli and Bifidobacterium; ↓Enterococci, Enterobacteria, Prevotellaceae, and Coriobacterales | May produce immune modulation and anti-tumor effects in the colon | [121] |
| Curcumin | Mice | NA (meta-analysis) | NA | ↑Bacteroides, Rikenellaceae, Alistipes, and Bacteroidaceae; ↓Prevotella and Prevotellaceae | Prevotella has been observed as higher in patients with CRC | [122] |
| Curcumin | Pilot study | 1000 mg of curcumin + 1.25 mg black pepper | 8 weeks | ↓Ruminococus and Blautia; ↑Clostridium and Enterobacter | Ruminococus species have been observed as higher in patients with CRC | [123] |
| Curcumin nanoparticles | Mice | 0.2 w/w | 7 days | ↑number of butyrate-producing bacteria and feal butyrate levels; ↓NF-ĸB activation in colonic epithelial cells | Increased SCFA production may reduce inflammatory processes and intestinal mucosa and promote antitumor effects | [124] |
| Curcumin | Mice | 8 mg/kg/day–162 mg/kg/day | 20 days | ↓Coriobacterales; ↑Lactobacillales | Decreased oxidative and inflammatory stresses, and hyper-immune activation | [125] |
| Curcumin | Mice | 20 mg/kg, 100 mg/kg, and 200 mg/kg | 10 days | ↓Enterobacteria and Enterococci; ↑Lactobacilli and Bifidobacteria | Suppressed pro-inflammatory processes and promoted anti-inflammatory effects | [126] |
| Ginger | Mice | 500 mg/kg daily | 7 days | ↓Lactobacillus murinus, Lachnospiraceae bacterium, and Ruminiclostridium specie KB18 | Reduced the expression of mRNA of IL-6 and iNOS | [127] |
| Ginger | Mice | 50 mg/kg | 4 weeks | Altered the abundance of Helicobacter and Peptococcaceae species | Ameliorated weight loss, colon shortening, inflammatory processes, intestinal barrier dysfunction, and gut dysbiosis | [128] |
| Daikenchuto, Japanese traditional herbal medicine (processed ginger, ginseng, and Chinese or Japanese pepper) | Human colonic microbiota | 0.5% wt | 48 h | ↑Bifidobacterium adolescentis | Bifidogenic effects may have beneficial effects on colon | [129] |
| Ginger polysaccharides | Mice | 200 mg/Kg | 1,3,5,7 and 9-day dose | Balancing Firmicutes/Bacteroidetes ratio; ↑Lactobacillus and Verrucomicrobiota; ↓Proteobacteria and Bacteroides | Reduced the level of colonic pro-inflammatory mediators (TNF-α, IL-6, IL-1β, IL-17A, and IFN-γ), restored gut barrier function, and restrained apoptosis | [130] |
| Ginger juice | Healthy volunteers | 500 mg/Kg/day | 7 days | ↓Ruminococcus_1 and Ruminococcus_2 and Prevotella/Bacteroides ratio; ↑Proteobacteria, Faecalibacterium, and Firmicutes/Bacteroidetes ratio | Promoted anti-inflammatory effects in intestinal mucosa | [131] |
| Garlic polysaccharides | Mice | NA (systematic review) | NA | ↑Bacteroidetes and Actinobacteria; ↓Firmicutes/Bacteroidetes ratio | Inhibited the expression of inflammatory mediators (TNF-α, IL-1β, and IL-6); Increased colon length and decrease in the disease activity and histological score of colitis | [132] |
| Propyl-propane thiosulfonate | Mice | 0.01, 0.05, 0.1, 0.5, 1, and 10 mg/kg day | 5 days | ↑Firmicutes/Bacteroidetes ratio; ↓Actinobacteria | Improved intestinal epithelial barrier integrity and reduced the expression of pro-inflammatory mediators (TNF-α, IL-1β, IL-8, IL-17, and iNOS) | [133] |
| Clove oil | Quails | 0.75 and 1.5 mL/Kg | 42 days | ↓Eescherechia coli, and Salmonella species | Improved body weight, activities of antioxidant enzymes, lipid profile, and intestinal bacterial diversity | [134] |
| Capsaicin | Healthy adults | 10 mg/day | 6 weeks | ↑Firmicutes/Bacteriodes ratio and Faecalibacterium abundance | Decreased inflammatory processes and risk factors for CRC | [135] |
| Crocin-I | Mice | 20 mg/kg and 40 mg/kg | 3 weeks | ↓Firmicutes; ↑Bacteroidetes | Increased α-diversity of microbes in the cecal contents | [136] |
| Crocetin | Mice | 10 mg/kg | 1 week | ↑Mediterraneibacter and Akkermansia; ↓Dubosiella, Muribaculaceae, Paramuribaculum, Allobaculum, Parasutterella, Duncaniella, Stoquefichus, Coriobacteriaceae UCG-002, and Candidatus. | Promoted inflammation with disturbed intestinal homeostasis | [137] |
| Saffron | Amnion of the Gallus gallus eggs | 1% CFWE, 2% CFWE, 5% CFWE, 10% CFWE. | Incubation until 21 days | ↓Lactobacillus and Clostridium | Disrupted cecal microbiome and brush border membrane functionality | [138] |
| Flaxseed | Mice | 10% FS diet | 1 week | ↓Akkermansia muciniphila; ↑Prevotella species | Decreased susceptibility to gut-associated diseases including inflammatory pathologies and cancer | [139] |
| Flaxseed oligosaccharides | Mice | 50 mg/kg day, 100 mg/kg day, and 200 mg/kg day | 14 days | ↓Clostridiales | Increased colon length, improved colonic histology, decreased oxidative stress markers (malondialdehyde and myeloperoxidase), suppressed pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), and increased anti-inflammatory cytokine (IL-10); Increased propionic and butyric acids | [140] |
| Flaxseed oil | Pigs | Flaxseed oil (FO, purity ≥ 98%) | 3 weeks | ↓Spirochaetes; ↑Actinobacteria, Bifidobacterium and Blautia | Decreased intestinal expression of MyD88, NF-κB, TNF-α, and IL-10 genes | [141] |
| Flaxseed | Mice | 12 weeks | ↑Prevotella, Ruminococcus, Clostridiales, and Paraprevotella | Increased butyrate concentration; Ameliorated the adherent-invasive E. coli induced intestinal inflammation | [142] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dacrema, M.; Ali, A.; Ullah, H.; Khan, A.; Di Minno, A.; Xiao, J.; Martins, A.M.C.; Daglia, M. Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota. Cancers 2022, 14, 5682. https://doi.org/10.3390/cancers14225682
Dacrema M, Ali A, Ullah H, Khan A, Di Minno A, Xiao J, Martins AMC, Daglia M. Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota. Cancers. 2022; 14(22):5682. https://doi.org/10.3390/cancers14225682
Chicago/Turabian StyleDacrema, Marco, Arif Ali, Hammad Ullah, Ayesha Khan, Alessandro Di Minno, Jianbo Xiao, Alice Maria Costa Martins, and Maria Daglia. 2022. "Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota" Cancers 14, no. 22: 5682. https://doi.org/10.3390/cancers14225682
APA StyleDacrema, M., Ali, A., Ullah, H., Khan, A., Di Minno, A., Xiao, J., Martins, A. M. C., & Daglia, M. (2022). Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota. Cancers, 14(22), 5682. https://doi.org/10.3390/cancers14225682