

Immune Checkpoint and Other Receptor-Ligand Pairs Modulating Macrophages in Cancer: Present and Prospects


Abstract
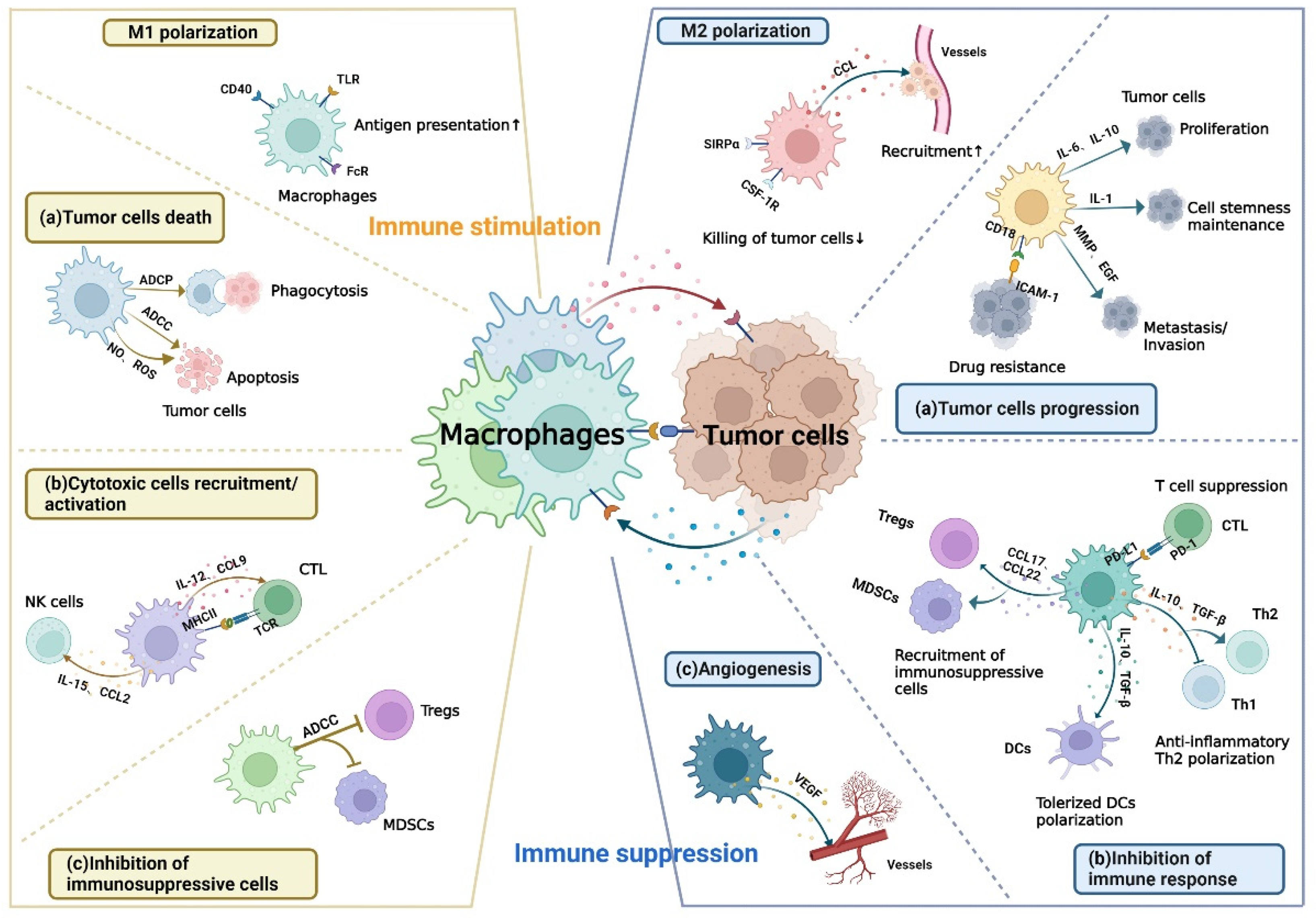
:Simple Summary
Abstract
1. Introduction
2. Immune Checkpoint Receptor-Ligand Interaction Dependent on Cell-Cell Contact
2.1. Rreceptor-Ligand Interaction in Anti-Tumor Immunity
2.1.1. FcγR-Antibody-Antigen Axis
2.1.2. CD40-CD40L
2.1.3. Fas-FasL
2.1.4. TRAIL-TRAIL-R
2.1.5. SLAMF7-SLAMF7 Axis
2.1.6. LRP1-CRT
2.1.7. CLEC10A-sTn Axis
2.2. Rreceptor-Ligand Interaction in Pro-Tumor Immunity
2.2.1. SIRPα-CD47 Axis
2.2.2. PD1-PD-L1 Axis
2.2.3. LILRB1-MHC-I Axis
2.2.4. MerTK-PROS1/GAS6-PtdSer Axis
2.2.5. Siglec15-sTn Axis
2.2.6. Siglec10-CD24 Axis
2.2.7. Ephrin-EphA4 Axis
2.2.8. EGF-EGFR Axis
2.2.9. SCF- c-Kit Axis
2.2.10. CEACAM1-Metadherin Axis
2.2.11. P-Selectin-PSGL-1 Axis
2.2.12. LSECtin-BTN3A3 Axis
2.2.13. Dectin1-Galectin9 Axis
2.2.14. α4-Integrin-VCAM-1 Axis
2.2.15. MAC1-CD90 Axis
2.2.16. MAC1-ICAM-1 Axis
3. Immune Checkpoint Receptor-Ligand Interaction Independent of Cell-Cell Contact
3.1. TLRs
3.2. CSF-1R
3.3. Chemokine Receptor
3.4. Interleukin Receptor
3.5. ATP/Adenosine Receptor
3.6. C5aR
3.7. VEGFR
3.8. TGF-βR
3.9. SUCNR1
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- He, X.; Xu, C. Immune checkpoint signaling and cancer immunotherapy. Cell Res. 2020, 30, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Couzin-Frankel, J. Cancer Immunotherapy. Science 2013, 342, 1432–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.C.; Duffy, C.R.; Allison, J.P. Fundamental Mechanisms of Immune Checkpoint Blockade Therapy. Cancer Discov. 2018, 8, 1069–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaikwad, S.; Agrawal, M.Y.; Kaushik, I.; Ramachandran, S.; Srivastava, S.K. Immune checkpoint proteins: Signaling mechanisms and molecular interactions in cancer immunotherapy. Semin. Cancer Biol. 2022, 86, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Vandsemb, E.N.; Herbst, R.S.; Chen, L. Adaptive immune resistance at the tumour site: Mechanisms and therapeutic opportunities. Nat. Rev. Drug Discov. 2022, 21, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Qin, L.; Liao, Z.; Song, J.; Yuan, C.; Liu, Y.; Wang, Y.; Xu, H.; Zhang, Q.; Pei, Y.; et al. Microenvironment characterization and multi-omics signatures related to prognosis and immunotherapy response of hepatocellular carcinoma. Exp. Hematol. Oncol. 2020, 9, 34. [Google Scholar] [CrossRef]
- Lopez-Yrigoyen, M.; Cassetta, L.; Pollard, J.W. Macrophage targeting in cancer. Ann. N. Y. Acad. Sci. 2020, 1499, 18–41. [Google Scholar] [CrossRef]
- Molgora, M.; Colonna, M. Turning enemies into allies—Reprogramming tumor-associated macrophages for cancer therapy. Med 2021, 2, 666–681. [Google Scholar] [CrossRef]
- Xiang, X.; Wang, J.; Di Lu, D.; Xu, X. Targeting tumor-associated macrophages to synergize tumor immunotherapy. Signal Transduct. Target. Ther. 2021, 6, 75. [Google Scholar] [CrossRef]
- Feng, M.; Jiang, W.; Kim, B.Y.S.; Zhang, C.C.; Fu, Y.-X.; Weissman, I.L. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 568–586. [Google Scholar] [CrossRef]
- Yu, J.; Song, Y.; Tian, W. How to select IgG subclasses in developing anti-tumor therapeutic antibodies. J. Hematol. Oncol. 2020, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Gupta, A.; Ravetch, J.V. The role of IgG Fc receptors in antibody-dependent enhancement. Nat. Rev. Immunol. 2020, 20, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Bakema, J.E.; Van Egmond, M. Fc receptor-dependent mechanisms of monoclonal antibody therapy of cancer. Curr. Top. Microbiol. Immunol. 2014, 382, 373–392. [Google Scholar] [CrossRef] [PubMed]
- Leidi, M.; Gotti, E.; Bologna, L.; Miranda, E.; Rimoldi, M.; Sica, A.; Roncalli, M.; Palumbo, G.A.; Introna, M.; Golay, J. M2 Macrophages Phagocytose Rituximab-Opsonized Leukemic Targets More Efficiently than M1 Cells In Vitro. J. Immunol. 2009, 182, 4415–4422. [Google Scholar] [CrossRef] [Green Version]
- Taskinen, M.; Karjalainen-Lindsberg, M.-L.; Nyman, H.; Eerola, L.-M.; Leppä, S. A High Tumor-Associated Macrophage Content Predicts Favorable Outcome in Follicular Lymphoma Patients Treated with Rituximab and Cyclophosphamide-Doxorubicin-Vincristine-Prednisone. Clin. Cancer Res. 2007, 13, 5784–5789. [Google Scholar] [CrossRef] [Green Version]
- Chao, M.P.; Alizadeh, A.A.; Tang, C.; Jan, M.; Weissman-Tsukamoto, R.; Zhao, F.; Park, C.Y.; Weissman, I.L.; Majeti, R. Therapeutic Antibody Targeting of CD47 Eliminates Human Acute Lymphoblastic Leukemia. Cancer Res. 2010, 71, 1374–1384. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Wei, H.; Gao, P.; Yu, H.; Wang, K.; Fu, Z.; Ju, B.; Zhao, M.; Dong, S.; Li, Z.; et al. CD47 promotes ovarian cancer progression by inhibiting macrophage phagocytosis. Oncotarget 2017, 8, 39021–39032. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.; Opzoomer, J.; Ilieva, K.M.; Gazinska, P.; Hoffmann, R.M.; Mirza, H.; Marlow, R.; Francesch-Domenech, E.; Fittall, M.; Rodriguez, D.D.; et al. Anti-Folate Receptor Alpha–Directed Antibody Therapies Restrict the Growth of Triple-negative Breast Cancer. Clin. Cancer Res. 2018, 24, 5098–5111. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, M.A.; Williamson, A.P.; Steinbach, A.M.; Roberts, E.W.; Kern, N.; Headley, M.B.; Vale, R.D. Chimeric antigen receptors that trigger phagocytosis. eLife 2018, 7, e36688. [Google Scholar] [CrossRef]
- Elgueta, R.; Benson, M.J.; De Vries, V.C.; Wasiuk, A.; Guo, Y.; Noelle, R.J. Molecular mechanism and function of CD40/CD40L engagement in the immune system. Immunol. Rev. 2009, 229, 152–172. [Google Scholar] [CrossRef]
- Baxendale, A.J.; Dawson, C.W.; E Stewart, S.; Mudaliar, V.; Reynolds, G.; Gordon, J.; Murray, P.G.; Young, L.S.; Eliopoulos, A.G. Constitutive activation of the CD40 pathway promotes cell transformation and neoplastic growth. Oncogene 2005, 24, 7913–7923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Cheng, X.; Truong, B.; Sun, L.; Yang, X.; Wang, H. Molecular basis and therapeutic implications of CD40/CD40L immune checkpoint. Pharmacol. Ther. 2020, 219, 107709. [Google Scholar] [CrossRef] [PubMed]
- Vonderheide, R.H. CD40 Agonist Antibodies in Cancer Immunotherapy. Annu. Rev. Med. 2020, 71, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, E.; Moreno, R.; Milenova, I.; Liljenfeldt, L.; Dieterich, L.C.; Christiansson, L.; Karlsson, H.; Ullenhag, G.; Mangsbo, S.M.; Dimberg, A.; et al. Activation of myeloid and endothelial cells by CD40L gene therapy supports T-cell expansion and migration into the tumor microenvironment. Gene Ther. 2016, 24, 92–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, M.; Imaizumi, K.; Kawabe, T.; Wakayama, H.; Horio, Y.; Sekido, Y.; Hara, T.; Hashimoto, N.; Takahashi, M.; Shimokata, K.; et al. Induction of antitumor immunity by transduction of CD40 ligand gene and interferon-γ gene into lung cancer. Cancer Gene Ther. 2001, 8, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Liljenfeldt, L.; Dieterich, L.C.; A Dimberg, A.; Mangsbo, S.M.; I Loskog, A.S. CD40L gene therapy tilts the myeloid cell profile and promotes infiltration of activated T lymphocytes. Cancer Gene Ther. 2014, 21, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Oehm, A.; Behrmann, I.; Falk, W.; Pawlita, M.; Maier, G.; Klas, C.; Li-Weber, M.; Richards, S.; Dhein, J.; Trauth, B. Purification and molecular cloning of the APO-1 cell surface antigen, a member of the tumor necrosis factor/nerve growth factor receptor superfamily. Sequence identity with the Fas antigen. J. Biol. Chem. 1992, 267, 10709–10715. [Google Scholar] [CrossRef]
- Suda, T.; Takahashi, T.; Golstein, P.; Nagata, S. Molecular cloning and expression of the fas ligand, a novel member of the tumor necrosis factor family. Cell 1993, 75, 1169–1178. [Google Scholar] [CrossRef]
- Villa-Morales, M.; Fernández-Piqueras, J. Targeting the Fas/FasL signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 85–101. [Google Scholar] [CrossRef]
- Scott, I.A. Positive and negative consequences of Fas/Fas ligand interactions in the antitumor response. Front. Biosci. 2005, 10, 809–821. [Google Scholar] [CrossRef]
- De Looff, M.; De Jong, S.; Kruyt, F.A.E. Multiple Interactions Between Cancer Cells and the Tumor Microenvironment Modulate TRAIL Signaling: Implications for TRAIL Receptor Targeted Therapy. Front. Immunol. 2019, 10, 1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Karstedt, S.; Montinaro, A.; Walczak, H. Exploring the TRAILs less travelled: TRAIL in cancer biology and therapy. Nat. Rev. Cancer 2017, 17, 352–366. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.S.; Yang, A.; Yang, B.; Couto, S.; Stern, H.; Gogineni, A.; Pitti, R.; Marsters, S.; Weimer, R.M.; Singh, M.; et al. Proapoptotic Activation of Death Receptor 5 on Tumor Endothelial Cells Disrupts the Vasculature and Reduces Tumor Growth. Cancer Cell 2012, 22, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragovich, M.A.; Mor, A. The SLAM family receptors: Potential therapeutic targets for inflammatory and autoimmune diseases. Autoimmun. Rev. 2018, 17, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Schwartzberg, P.L.; Mueller, K.L.; Qi, H.; Cannons, J.L. SLAM receptors and SAP influence lymphocyte interactions, development and function. Nat. Rev. Immunol. 2009, 9, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Claus, M.; Urlaub, D.; Fasbender, F.; Watzl, C. SLAM family receptors in natural killer cells–Mediators of adhesion, activation and inhibition via cis and trans interactions. Clin. Immunol. 2018, 204, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Abram, C.L.; Lowell, C.A. The Ins and Outs of Leukocyte Integrin Signaling. Annu. Rev. Immunol. 2009, 27, 339–362. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhong, M.-C.; Guo, H.; Davidson, D.; Mishel, S.; Lu, Y.; Rhee, I.; Pérez-Quintero, L.-A.; Zhang, S.; Cruz-Munoz, M.-E.; et al. SLAMF7 is critical for phagocytosis of haematopoietic tumour cells via Mac-1 integrin. Nature 2017, 544, 493–497. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Bouwstra, R.; Wiersma, V.R.; De Jong, M.; Lourens, H.J.; Fehrmann, R.; De Bruyn, M.; Ammatuna, E.; Huls, G.; Van Meerten, T.; et al. Cancer cell-expressed SLAMF7 is not required for CD47-mediated phagocytosis. Nat. Commun. 2019, 10, 533. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Williams, B.O. Low-Density Lipoprotein Receptor-Related Proteins in Skeletal Development and Disease. Physiol. Rev. 2017, 97, 1211–1228. [Google Scholar] [CrossRef]
- Calvier, L.; Boucher, P.; Herz, J.; Hansmann, G. LRP1 Deficiency in Vascular SMC Leads to Pulmonary Arterial Hypertension That Is Reversed by PPARγ Activation. Circ. Res. 2019, 124, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Zhao, Z.; Sagare, A.P.; Wu, Y.; Wang, M.; Owens, N.C.; Verghese, P.B.; Herz, J.; Holtzman, D.M.; Zlokovic, B.V. Blood-brain barrier-associated pericytes internalize and clear aggregated amyloid-β42 by LRP1-dependent apolipoprotein E isoform-specific mechanism. Mol. Neurodegener. 2018, 13, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, P.; Zhu, L.; Tavori, H.; Huynh, K.; Giunzioni, I.; Stafford, J.; Linton, M.F.; Fazio, S. Deletion of Macrophage Low-Density Lipoprotein Receptor-Related Protein 1 (LRP1) Accelerates Atherosclerosis Regression and Increases C-C Chemokine Receptor Type 7 (CCR7) Expression in Plaque Macrophages. Circulation 2018, 138, 1850–1863. [Google Scholar] [CrossRef] [PubMed]
- Wujak, L.; Schnieder, J.; Schaefer, L.; Wygrecka, M. LRP1: A chameleon receptor of lung inflammation and repair. Matrix Biol. 2018, 68-69, 366–381. [Google Scholar] [CrossRef]
- Gardai, S.J.; McPhillips, K.A.; Frasch, S.C.; Janssen, W.J.; Starefeldt, A.; Murphy-Ullrich, J.E.; Bratton, D.L.; Oldenborg, P.-A.; Michalak, M.; Henson, P.M. Cell-Surface Calreticulin Initiates Clearance of Viable or Apoptotic Cells through trans-Activation of LRP on the Phagocyte. Cell 2005, 123, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Fucikova, J.; Spisek, R.; Kroemer, G.; Galluzzi, L. Calreticulin and cancer. Cell Res. 2021, 31, 5–16. [Google Scholar] [CrossRef]
- Schcolnik-Cabrera, A.; Oldak, B.; Juárez, M.; Cruz-Rivera, M.; Flisser, A.; Mendlovic, F. Calreticulin in phagocytosis and cancer: Opposite roles in immune response outcomes. Apoptosis 2019, 24, 245–255. [Google Scholar] [CrossRef]
- Chao, M.P.; Jaiswal, S.; Weissman-Tsukamoto, R.; Alizadeh, A.A.; Gentles, A.J.; Volkmer, J.; Weiskopf, K.; Willingham, S.B.; Raveh, T.; Park, C.Y.; et al. Calreticulin Is the Dominant Pro-Phagocytic Signal on Multiple Human Cancers and Is Counterbalanced by CD47. Sci. Transl. Med. 2010, 2, 63ra94. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Yamamoto, K.; Toyoshima, S.; Osawa, T.; Irimura, T. Molecular cloning and expression of cDNA encoding human macrophage C-type lectin. Its unique carbohydrate binding specificity for Tn antigen. J. Immunol. 1996, 156, 128–135. [Google Scholar]
- van Vliet, S.J.; Saeland, E.; van Kooyk, Y. Sweet preferences of MGL: Carbohydrate specificity and function. Trends Immunol. 2008, 29, 83–90. [Google Scholar] [CrossRef] [PubMed]
- van Kooyk, Y.; Ilarregui, J.M.; van Vliet, S.J. Novel insights into the immunomodulatory role of the dendritic cell and macrophage-expressed C-type lectin MGL. Immunobiology 2015, 220, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Wang, L.; Zhang, L.; Li, J.; Liao, L.; Huang, L.; Yang, J. Immunological role and prognostic potential of CLEC10A in pan-cancer. Am. J. Transl. Res. 2022, 14, 2844–2860. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Zhang, Y.; Lin, X.; Wang, H.; Yong, L.; Zhang, H.; Cai, F. CLEC10A can serve as a potential therapeutic target and its level correlates with immune infiltration in breast cancer. Oncol. Lett. 2022, 24, 285. [Google Scholar] [CrossRef] [PubMed]
- Mortezai, N.; Behnken, H.N.; Kurze, A.-K.; Ludewig, P.; Buck, F.; Meyer, B.; Wagener, C. Tumor-associated Neu5Ac-Tn and Neu5Gc-Tn antigens bind to C-type lectin CLEC10A (CD301, MGL). Glycobiology 2013, 23, 844–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veillette, A.; Chen, J. SIRPα–CD47 Immune Checkpoint Blockade in Anticancer Therapy. Trends Immunol. 2018, 39, 173–184. [Google Scholar] [CrossRef]
- Adams, S.; Van Der Laan, L.J.; Vernon-Wilson, E.; De Lavalette, C.R.; A Döpp, E.; Dijkstra, C.D.; Simmons, D.L.; Berg, T.K.V.D. Signal-regulatory protein is selectively expressed by myeloid and neuronal cells. J. Immunol. 1998, 161, 1853–1859. [Google Scholar]
- Bian, H.-T.; Shen, Y.-W.; Zhou, Y.-D.; Nagle, D.G.; Guan, Y.-Y.; Zhang, W.-D.; Luan, X. CD47: Beyond an immune checkpoint in cancer treatment. Biochim. Biophys. Acta 2022, 1877, 188771. [Google Scholar] [CrossRef]
- Seiffert, M.; Cant, C.; Chen, Z.; Rappold, I.; Brugger, W.; Kanz, L.; Brown, E.J.; Ullrich, A.; Bühring, H.-J. Human Signal-Regulatory Protein Is Expressed on Normal, But Not on Subsets of Leukemic Myeloid Cells and Mediates Cellular Adhesion Involving Its Counterreceptor CD47. Blood 1999, 94, 3633–3643. [Google Scholar] [CrossRef]
- Campbell, I.G.; Freemont, P.S.; Foulkes, W.; Trowsdale, J. An Ovarian Tumor Marker with Homology to Vaccinia Virus Contains an IgV-like Region and Multiple Transmembrane Domains. Cancer Res. 1992, 52, 5416–5420. [Google Scholar]
- Russ, A.; Hua, A.B.; Montfort, W.R.; Rahman, B.; Bin Riaz, I.; Khalid, M.U.; Carew, J.S.; Nawrocki, S.T.; Persky, D.; Anwer, F. Blocking “don’t eat me” signal of CD47-SIRPα in hematological malignancies, an in-depth review. Blood Rev. 2018, 32, 480–489. [Google Scholar] [CrossRef]
- Liu, Y.; Merlin, D.; Burst, S.L.; Pochet, M.; Madara, J.L.; Parkos, C.A. The Role of CD47 in Neutrophil Transmigration. J. Biol. Chem. 2001, 276, 40156–40166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhold, M.I.; Lindberg, F.P.; Kersh, G.J.; Allen, P.M.; Brown, E.J. Costimulation of T Cell Activation by Integrin-associated Protein (CD47) Is an Adhesion-dependent, CD28-independent Signaling Pathway. J. Exp. Med. 1997, 185, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barclay, A.N.; Brown, M.H. The SIRP family of receptors and immune regulation. Nat. Rev. Immunol. 2006, 6, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Tsai, R.K.; Discher, D.E. Inhibition of “self” engulfment through deactivation of myosin-II at the phagocytic synapse between human cells. J. Cell Biol. 2008, 180, 989–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eladl, E.; Tremblay-LeMay, R.; Rastgoo, N.; Musani, R.; Chen, W.; Liu, A.; Chang, H. Role of CD47 in Hematological Malignancies. J. Hematol. Oncol. 2020, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.P.; Alizadeh, A.A.; Tang, C.; Myklebust, J.H.; Varghese, B.; Gill, S.; Jan, M.; Cha, A.C.; Chan, C.K.; Tan, B.T.; et al. Anti-CD47 Antibody Synergizes with Rituximab to Promote Phagocytosis and Eradicate Non-Hodgkin Lymphoma. Cell 2010, 142, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Sockolosky, J.T.; Dougan, M.; Ingram, J.R.; Ho, C.C.M.; Kauke, M.J.; Almo, S.C.; Ploegh, H.L.; Garcia, K.C. Durable antitumor responses to CD47 blockade require adaptive immune stimulation. Proc. Natl. Acad. Sci. USA 2016, 113, E2646–E2654. [Google Scholar] [CrossRef] [Green Version]
- Deuse, T.; Hu, X.; Agbor-Enoh, S.; Jang, M.K.; Alawi, M.; Saygi, C.; Gravina, A.; Tediashvili, G.; Nguyen, V.Q.; Liu, Y.; et al. The SIRPα–CD47 immune checkpoint in NK cells. J. Exp. Med. 2021, 218, e20200839. [Google Scholar] [CrossRef]
- Velliquette, R.W.; Aeschlimann, J.; Kirkegaard, J.; Shakarian, G.; Lomas-Francis, C.; Westhoff, C.M. Monoclonal anti-CD47 interference in red cell and platelet testing. Transfusion 2018, 59, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Ishida, Y.; Agata, Y.; Shibahara, K.; Honjo, T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. 1992, 11, 3887–3895. [Google Scholar] [CrossRef]
- Sharpe, A.H.; Pauken, K.E. The diverse functions of the PD1 inhibitory pathway. Nat. Rev. Immunol. 2017, 18, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.M.; McCracken, M.N.; Gupta, R.; Tsai, J.M.; Sinha, R.; Corey, D.; et al. PD-1 expression by tumour-associated macrophages inhibits phagocytosis and tumour immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, J.; Qi, Q.; Qian, X.; Han, J.; Zhu, X.; Zhang, Q.; Xia, R. The role of PD-1/PD-L1 axis and macrophage in the progression and treatment of cancer. J. Cancer Res. Clin. Oncol. 2019, 145, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Strome, S.E.; Salomao, D.R.; Tamura, H.; Hirano, F.; Flies, D.B.; Roche, P.C.; Lu, J.; Zhu, G.; Tamada, K.; et al. Tumor-associated B7-H1 promotes T-cell apoptosis: A potential mechanism of immune evasion. Nat. Med. 2002, 8, 793–800. [Google Scholar] [CrossRef]
- Dhupkar, P.; Gordon, N.; Stewart, J.; Kleinerman, E.S. Anti-PD-1 therapy redirects macrophages from an M2 to an M1 phenotype inducing regression of OS lung metastases. Cancer Med. 2018, 7, 2654–2664. [Google Scholar] [CrossRef]
- Colonna, M.; Nakajima, H.; Navarro, F.; López-Botet, M. A novel family of Ig-like receptors for HLA class I molecules that modulate function of lymphoid and myeloid cells. J. Leukoc. Biol. 1999, 66, 375–381. [Google Scholar] [CrossRef]
- Mori, Y.; Tsuji, S.; Inui, M.; Sakamoto, Y.; Endo, S.; Ito, Y.; Fujimura, S.; Koga, T.; Nakamura, A.; Takayanagi, H.; et al. Inhibitory Immunoglobulin-Like Receptors LILRB and PIR-B Negatively Regulate Osteoclast Development. J. Immunol. 2008, 181, 4742–4751. [Google Scholar] [CrossRef] [Green Version]
- Munitz, A.; Cole, E.T.; Beichler, A.; Groschwitz, K.; Ahrens, R.; Steinbrecher, K.; Willson, T.; Han, X.; Denson, L.; Rothenberg, M.E.; et al. Paired Immunoglobulin-Like Receptor B (PIR-B) Negatively Regulates Macrophage Activation in Experimental Colitis. Gastroenterology 2010, 139, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Cella, M.; Nakajima, H.; Facchetti, F.; Hoffmann, T.; Colonna, M. ILT Receptors at the Interface Between Lymphoid and Myeloid. Cells 2000, 251, 161–166. [Google Scholar] [CrossRef]
- E Willcox, B.; Thomas, L.M.; Bjorkman, P.J. Crystal structure of HLA-A2 bound to LIR-1, a host and viral major histocompatibility complex receptor. Nat. Immunol. 2003, 4, 913–919. [Google Scholar] [CrossRef] [Green Version]
- Rock, K.L.; Reits, E.; Neefjes, J. Present Yourself! By MHC Class I and MHC Class II Molecules. Trends Immunol. 2016, 37, 724–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkal, A.A.; Weiskopf, K.; Kao, K.S.; Gordon, S.R.; Rosental, B.; Yiu, Y.Y.; George, B.M.; Markovic, M.; Ring, N.G.; Tsai, J.M.; et al. Engagement of MHC class I by the inhibitory receptor LILRB1 suppresses macrophages and is a target of cancer immunotherapy. Nat. Immunol. 2018, 19, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Pan, P.-Y.; Eisenstein, S.; Divino, C.M.; Lowell, C.A.; Takai, T.; Chen, S.-H. Paired Immunoglobin-like Receptor-B Regulates the Suppressive Function and Fate of Myeloid-Derived Suppressor Cells. Immunity 2011, 34, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Linger, R.M.; Keating, A.K.; Earp, H.S.; Graham, D.K. TAM Receptor Tyrosine Kinases: Biologic Functions, Signaling, and Potential Therapeutic Targeting in Human Cancer. Adv. Cancer Res. 2008, 100, 35–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, D.K.; DeRyckere, D.; Davies, K.D.; Earp, H.S. The TAM family: Phosphatidylserine-sensing receptor tyrosine kinases gone awry in cancer. Nat. Rev. Cancer 2014, 14, 769–785. [Google Scholar] [CrossRef] [PubMed]
- Stitt, T.N.; Conn, G.; Goret, M.; Lai, C.; Bruno, J.; Radzlejewski, C.; Mattsson, K.; Fisher, J.; Gies, D.R.; Jones, P.F.; et al. The anticoagulation factor protein S and its relative, Gas6, are ligands for the Tyro 3/Axl family of receptor tyrosine kinases. Cell 1995, 80, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Aehnlich, P.; Powell, R.; Peeters, M.; Rahbech, A.; Straten, P.T. TAM Receptor Inhibition–Implications for Cancer and the Immune System. Cancers 2021, 13, 1195. [Google Scholar] [CrossRef]
- Wang, J.; Yu, C.; Zhuang, J.; Qi, W.; Jiang, J.; Liu, X.; Zhao, W.; Cao, Y.; Wu, H.; Qi, J.; et al. The role of phosphatidylserine on the membrane in immunity and blood coagulation. Biomark. Res. 2022, 10, 4. [Google Scholar] [CrossRef]
- Burstyn-Cohen, T.; Maimon, A. TAM receptors, Phosphatidylserine, inflammation, and Cancer. Cell Commun. Signal. 2019, 17, 156. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Fei, M.; Zhang, G.; Liang, W.-C.; Lin, W.; Wu, Y.; Piskol, R.; Ridgway, J.; McNamara, E.; Huang, H.; et al. Blockade of the Phagocytic Receptor MerTK on Tumor-Associated Macrophages Enhances P2X7R-Dependent STING Activation by Tumor-Derived cGAMP. Immunity 2020, 52, 357–373.e9. [Google Scholar] [CrossRef]
- Sun, J.; Lu, Q.; Sanmanmed, M.F.; Wang, J. Siglec-15 as an Emerging Target for Next-generation Cancer Immunotherapy. Clin. Cancer Res. 2021, 27, 680–688. [Google Scholar] [CrossRef]
- Wang, J.; Sun, J.; Liu, L.N.; Flies, D.B.; Nie, X.; Toki, M.; Zhang, J.; Song, C.; Zarr, M.; Zhou, X.; et al. Siglec-15 as an immune suppressor and potential target for normalization cancer immunotherapy. Nat. Med. 2019, 25, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Angata, T.; Tabuchi, Y.; Nakamura, K.; Nakamura, M. Siglec-15: An immune system Siglec conserved throughout vertebrate evolution. Glycobiology 2007, 17, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, R.; Ohtsubo, K.; Takamatsu, S.; Taniguchi, N.; Angata, T. The interaction between Siglec-15 and tumor-associated sialyl-Tn antigen enhances TGF- secretion from monocytes/macrophages through the DAP12-Syk pathway. Glycobiology 2012, 23, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angata, T.; Varki, A. Discovery, classification, evolution and diversity of Siglecs. Mol. Asp. Med. 2022, 101117. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.C.; Macauley, M.S.; Kawasaki, N. Siglecs as sensors of self in innate and adaptive immune responses. Ann. N. Y. Acad. Sci. 2012, 1253, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhang, W.; Wan, T.; Zhang, J.; Chen, T.; Yu, Y.; Wang, J.; Cao, X. Cloning and Characterization of Siglec-10, a Novel Sialic Acid Binding Member of the Ig Superfamily, from Human Dendritic Cells. J. Biol. Chem. 2001, 276, 28106–28112. [Google Scholar] [CrossRef] [Green Version]
- Altevogt, P.; Sammar, M.; Hüser, L.; Kristiansen, G. Novel insights into the function ofCD24: A driving force in cancer. Int. J. Cancer 2020, 148, 546–559. [Google Scholar] [CrossRef]
- Toubai, T.; Hou, G.; Mathewson, N.; Liu, C.; Wang, Y.; Oravecz-Wilson, K.; Cummings, E.; Rossi, C.; Evers, R.; Sun, Y.; et al. Siglec-G–CD24 axis controls the severity of graft-versus-host disease in mice. Blood 2014, 123, 3512–3523. [Google Scholar] [CrossRef] [Green Version]
- Kristiansen, G.; Sammar, M.; Altevogt, P. Tumour Biological Aspects of CD24, A Mucin-Like Adhesion Molecule. Histochem. J. 2003, 35, 255–262. [Google Scholar] [CrossRef]
- Chen, G.-Y.; Tang, J.; Zheng, P.; Liu, Y. CD24 and Siglec-10 Selectively Repress Tissue Damage–Induced Immune Responses. Science 2009, 323, 1722–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkal, A.A.; Brewer, R.E.; Markovic, M.; Kowarsky, M.; Barkal, S.A.; Zaro, B.W.; Krishnan, V.; Hatakeyama, J.; Dorigo, O.; Barkal, L.J.; et al. CD24 signalling through macrophage Siglec-10 is a target for cancer immunotherapy. Nature 2019, 572, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Aroldi, A.; Mauri, M.; Parma, M.; Terruzzi, E.; Fedele, M.; Perfetti, P.; Cocito, F.; Mologni, L.; Chiarle, R.; Piazza, R.; et al. CD24/Siglec-10 “Don’t Eat Me” Signal Blockade Is a Potential Immunotherapeutic Target in Mantle-Cell Lymphoma. Blood 2021, 138, 2276. [Google Scholar] [CrossRef]
- Bradley, C.A. CD24—a novel ‘don’t eat me’ signal. Nat. Rev. Drug Discov. 2019, 18, 747. [Google Scholar] [CrossRef]
- Stanczak, M.A.; Läubli, H. Siglec receptors as new immune checkpoints in cancer. Mol. Asp. Med. 2022, 101112. [Google Scholar] [CrossRef]
- Darling, T.K.; Lamb, T.J. Emerging Roles for Eph Receptors and Ephrin Ligands in Immunity. Front. Immunol. 2019, 10, 1473. [Google Scholar] [CrossRef] [Green Version]
- Rudno-Rudzińska, J.; Kielan, W.; Frejlich, E.; Kotulski, K.; Hap, W.; Kurnol, K.; Dzierżek, P.; Zawadzki, M.; Hałoń, A.; 2-nd Department of General and Oncological Surgery. A review on Eph/ephrin, angiogenesis and lymphangiogenesis in gastric, colorectal and pancreatic cancers. Chin. J. Cancer Res. 2017, 29, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Pasquale, E.B. Eph-Ephrin Bidirectional Signaling in Physiology and Disease. Cell 2008, 133, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Clauser, K.R.; Tam, W.L.; Frose, J.; Ye, X.; Eaton, E.N.; Reinhardt, F.; Donnenberg, V.S.; Bhargava, R.; Carr, S.A.; et al. A breast cancer stem cell niche supported by juxtacrine signalling from monocytes and macrophages. Nat. Cell Biol. 2014, 16, 1105–1117. [Google Scholar] [CrossRef] [Green Version]
- Koseska, A.; Bastiaens, P.I. Processing Temporal Growth Factor Patterns by an Epidermal Growth Factor Receptor Network Dynamically Established in Space. Annu. Rev. Cell Dev. Biol. 2020, 36, 359–383. [Google Scholar] [CrossRef]
- Kumar, R.; George, B.; Campbell, M.R.; Verma, N.; Paul, A.M.; Melo-Alvim, C.; Ribeiro, L.; Pillai, M.R.; da Costa, L.M.; Moasser, M.M. HER family in cancer progression: From discovery to 2020 and beyond. Adv. Cancer Res. 2020, 147, 109–160. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, G.; Cohen, S. Epidermal growth factor. J. Biol. Chem. 1990, 265, 7709–7712. [Google Scholar] [CrossRef] [PubMed]
- Vouri, M.; Hafizi, S. TAM Receptor Tyrosine Kinases in Cancer Drug Resistance. Cancer Res. 2017, 77, 2775–2778. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.C. EGF receptor ligands. Exp. Cell Res. 2003, 284, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Denning, M.F.; Dlugosz, A.A.; Cheng, C.; Dempsey, P.J.; Coffey, R.J., Jr.; Threadgill, D.W.; Magnuson, T.; Yuspa, S.H. Cross-talk between epidermal growth factor receptor and protein kinase C during calcium-induced differentiation of keratinocytes. Exp. Dermatol. 2000, 9, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.B.; Tsukada, T.; Zent, R.; Harris, R.C. Membrane-associated HB-EGF modulates HGF-induced cellular responses in MDCK cells. J. Cell Sci. 2004, 117, 1365–1379. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, R.; Handa, K.; Mekada, E. Contact-dependent Growth Inhibition and Apoptosis of Epidermal Growth Factor (EGF) Receptor-expressing Cells by the Membrane-anchored Form of Heparin-binding EGF-like Growth Factor. J. Biol. Chem. 1999, 274, 25906–25912. [Google Scholar] [CrossRef] [Green Version]
- Onal, S.; Turker-Burhan, M.; Bati-Ayaz, G.; Yanik, H.; Pesen-Okvur, D. Breast cancer cells and macrophages in a paracrine-juxtacrine loop. Biomaterials 2020, 267, 120412. [Google Scholar] [CrossRef]
- Pathania, S.; Pentikäinen, O.T.; Singh, P.K. A holistic view on c-Kit in cancer: Structure, signaling, pathophysiology and its inhibitors. Biochim. Biophys. Acta 2021, 1876, 188631. [Google Scholar] [CrossRef]
- Han, Z.-B.; Ren, H.; Zhao, H.; Chi, Y.; Chen, K.; Zhou, B.; Liu, Y.-J.; Zhang, L.; Xu, B.; Liu, B.; et al. Hypoxia-inducible factor (HIF)-1 directly enhances the transcriptional activity of stem cell factor (SCF) in response to hypoxia and epidermal growth factor (EGF). Carcinogenesis 2008, 29, 1853–1861. [Google Scholar] [CrossRef]
- Levina, V.; Marrangoni, A.; Wang, T.; Parikh, S.; Su, Y.; Herberman, R.; Lokshin, A.; Gorelik, E. Elimination of Human Lung Cancer Stem Cells through Targeting of the Stem Cell Factor–c-kit Autocrine Signaling Loop. Cancer Res. 2010, 70, 338–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanulla, M.; Welte, K.; Hadam, M.R.; Pietsch, T. Coexpression of stem cell factor and its receptor c-Kit in human malignant glioma cell lines. Acta Neuropathol. 1995, 89, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Lin, J.; Gao, L.; Deng, A.; Lu, X.; Li, Y.; Wang, L.; Yu, L. High Expression of c-kit mRNA Predicts Unfavorable Outcome in Adult Patients with t(8;21) Acute Myeloid Leukemia. PLoS ONE 2015, 10, e0124241. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Anton, A.; Gras, D.; Bourdin, A.; Dubreuil, P.; Chanez, P. KIT as a therapeutic target for non-oncological diseases. Pharmacol. Ther. 2018, 197, 11–37. [Google Scholar] [CrossRef]
- Lennartsson, J.; Rönnstrand, L. Stem Cell Factor Receptor/c-Kit: From Basic Science to Clinical Implications. Physiol. Rev. 2012, 92, 1619–1649. [Google Scholar] [CrossRef] [Green Version]
- Rönnstrand, L. Signal transduction via the stem cell factor receptor/c-Kit. Cell. Mol. Life Sci. 2004, 61, 2535–2548. [Google Scholar] [CrossRef]
- Mazzoldi, E.L.; Pavan, S.; Pilotto, G.; Leone, K.; Pagotto, A.; Frezzini, S.; Nicoletto, M.O.; Amadori, A.; Pastò, A. A juxtacrine/paracrine loop between C-Kit and stem cell factor promotes cancer stem cell survival in epithelial ovarian cancer. Cell Death Dis. 2019, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.M.; Huang, Y.-H.; Gandhi, A.; Blumberg, R.S. CEACAM1 structure and function in immunity and its therapeutic implications. Semin. Immunol. 2019, 42, 101296. [Google Scholar] [CrossRef]
- Müller, M.; Klaile, E.; Vorontsova, O.; Singer, B.B.; Öbrink, B. Homophilic adhesion and CEACAM1-S regulate dimerization of CEACAM1-L and recruitment of SHP-2 and c-Src. J. Cell Biol. 2009, 187, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Beauchemin, N.; Arabzadeh, A. Carcinoembryonic antigen-related cell adhesion molecules (CEACAMs) in cancer progression and metastasis. Cancer Metastasis Rev. 2013, 32, 643–671. [Google Scholar] [CrossRef]
- To, S.K.Y.; Tang, M.K.S.; Tong, Y.; Zhang, J.; Chan, K.K.L.; Ip, P.P.C.; Shi, J.; Wong, A.S.T. A Selective β −Catenin-Metadherin/CEACAM1-CCL3 Axis Mediates Metastatic Heterogeneity upon Tumor–Macrophage Interaction. Adv. Sci. 2022, 9, 2103230. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, Q. Astrocyte elevated gene-1 and breast cancer (Review). Oncol. Lett. 2011, 2, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Ortenberg, R.; Sapir, Y.; Raz, L.; Hershkovitz, L.; Ben Arav, A.; Sapoznik, S.; Barshack, I.; Avivi, C.; Berkun, Y.; Besser, M.J.; et al. Novel Immunotherapy for Malignant Melanoma with a Monoclonal Antibody That Blocks CEACAM1 Homophilic Interactions. Mol. Cancer Ther. 2012, 11, 1300–1310. [Google Scholar] [CrossRef]
- McEver, R.P.; Beckstead, J.H.; Moore, K.L.; Marshall-Carlson, L.; Bainton, D.F. GMP-140, a platelet alpha-granule membrane protein, is also synthesized by vascular endothelial cells and is localized in Weibel-Palade bodies. J. Clin. Investig. 1989, 84, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, R.; Otero, D.C.; Takahashi, A.A.; Bradley, L.M. PSGL-1: A New Player in the Immune Checkpoint Landscape. Trends Immunol. 2017, 38, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Frenette, P.S.; Denis, C.; Weiss, L.; Jurk, K.; Subbarao, S.; Kehrel, B.; Hartwig, J.H.; Vestweber, D.; Wagner, D.D. P-Selectin Glycoprotein Ligand 1 (Psgl-1) Is Expressed on Platelets and Can Mediate Platelet–Endothelial Interactions in Vivo. J. Exp. Med. 2000, 191, 1413–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Zuchtriegel, G.; Uhl, B.; Puhr-Westerheide, D.; Pörnbacher, M.; Lauber, K.; Krombach, F.; Reichel, C.A. Platelets Guide Leukocytes to Their Sites of Extravasation. PLOS Biol. 2016, 14, e1002459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Yang, J.; Qian, J.; Qiu, P.; Hanabuchi, S.; Lu, Y.; Wang, Z.; Liu, Z.; Li, H.; He, J.; et al. PSGL-1/selectin and ICAM-1/CD18 interactions are involved in macrophage-induced drug resistance in myeloma. Leukemia 2012, 27, 702–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, P.; Phennicie, R.; Kauffman, K.; Nowakowska, D.; Zafari, M.; Komoroski, V.; Sazinsky, S.; Wahle, J.; Manfra, D.; Vaidya, S.; et al. 862 Targeting PSGL-1, a novel macrophage checkpoint, repolarizes suppressive macrophages, induces an inflammatory tumor microenvironment, and suppresses tumor growth. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef]
- Liu, W.; Tang, L.; Zhang, G.; Wei, H.; Cui, Y.; Guo, L.; Gou, Z.; Chen, X.; Jiang, D.; Zhu, Y.; et al. Characterization of a Novel C-type Lectin-like Gene, LSECtin. J. Biol. Chem. 2004, 279, 18748–18758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Hao, B.; Kuai, X.; Xing, G.; Yang, J.; Chen, J.; Tang, L.; Zhang, L.; He, F. C-type lectin LSECtin interacts with DC-SIGNR and is involved in hepatitis C virus binding. Mol. Cell. Biochem. 2009, 327, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Powlesland, A.S.; Fisch, T.; Taylor, M.; Smith, D.F.; Tissot, B.; Dell, A.; Pöhlmann, S.; Drickamer, K. A Novel Mechanism for LSECtin Binding to Ebola Virus Surface Glycoprotein through Truncated Glycans. J. Biol. Chem. 2008, 283, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Gramberg, T.; Hofmann, H.; Möller, P.; Lalor, P.; Marzi, A.; Geier, M.; Krumbiegel, M.; Winkler, T.; Kirchhoff, F.; Adams, D.; et al. LSECtin interacts with filovirus glycoproteins and the spike protein of SARS coronavirus. Virology 2005, 340, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Wang, M.; Wang, X.; Zhao, D.; Liu, D.; Liu, J.; Chen, P.-J.; Yang, D.; He, F.; Tang, L. Liver Sinusoidal Endothelial Cell Lectin Inhibits CTL-Dependent Virus Clearance in Mouse Models of Viral Hepatitis. J. Immunol. 2013, 190, 4185–4195. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Lu, Q.; Wang, X.; Wang, J.; Lu, N.; Jiang, Z.; Hao, X.; Li, J.; Liu, J.; Cao, P.; et al. LSECtin on tumor-associated macrophages enhances breast cancer stemness via interaction with its receptor BTN3A3. Cell Res. 2019, 29, 365–378. [Google Scholar] [CrossRef]
- Chen, S.; Li, Z.; Huang, W.; Wang, Y.; Fan, S. Prognostic and Therapeutic Significance of BTN3A Proteins in Tumors. J. Cancer 2021, 12, 4505–4512. [Google Scholar] [CrossRef]
- Peedicayil, A.; Vierkant, R.A.; Hartmann, L.C.; Fridley, B.L.; Fredericksen, Z.S.; White, K.L.; Elliott, E.A.; Phelan, C.M.; Tsai, Y.-Y.; Berchuck, A.; et al. Risk of Ovarian Cancer and Inherited Variants in Relapse-Associated Genes. PLoS ONE 2010, 5, e8884. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.; Tsoni, S.V.; Willment, J.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S.; et al. Dectin-1 is required for β-glucan recognition and control of fungal infection. Nat. Immunol. 2006, 8, 31–38. [Google Scholar] [CrossRef]
- Tone, K.; Stappers, M.H.; Willment, J.A.; Brown, G.D. C-type lectin receptors of the Dectin-1 cluster: Physiological roles and involvement in disease. Eur. J. Immunol. 2019, 49, 2127–2133. [Google Scholar] [CrossRef] [Green Version]
- Daley, D.; Mani, V.R.; Mohan, N.; Akkad, N.; Ochi, A.; Heindel, D.W.; Lee, K.B.; Zambirinis, C.P.; Pandian, G.S.B.; Savadkar, S.; et al. Dectin 1 activation on macrophages by galectin 9 promotes pancreatic carcinoma and peritumoral immune tolerance. Nat. Med. 2017, 23, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Wada, J.; Kanwar, Y.S. Identification and Characterization of Galectin-9, a Novel β-Galactoside-binding Mammalian Lectin. J. Biol. Chem. 1997, 272, 6078–6086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, I.G.; Yasinska, I.M.; Sakhnevych, S.S.; Fiedler, W.; Wellbrock, J.; Bardelli, M.; Varani, L.; Hussain, R.; Siligardi, G.; Ceccone, G.; et al. The Tim-3-galectin-9 Secretory Pathway is Involved in the Immune Escape of Human Acute Myeloid Leukemia Cells. EBioMedicine 2017, 22, 44–57. [Google Scholar] [CrossRef]
- Schlichtner, S.; Meyer, N.H.; Yasinska, I.M.; Aliu, N.; Berger, S.M.; Gibbs, B.F.; Fasler-Kan, E.; Sumbayev, V.V. Functional role of galectin-9 in directing human innate immune reactions to Gram-negative bacteria and T cell apoptosis. Int. Immunopharmacol. 2021, 100, 108155. [Google Scholar] [CrossRef]
- Lietha, D.; Izard, T. Roles of Membrane Domains in Integrin-Mediated Cell Adhesion. Int. J. Mol. Sci. 2020, 21, 5531. [Google Scholar] [CrossRef] [PubMed]
- Hood, J.D.; Cheresh, D.A. Role of integrins in cell invasion and migration. Nat. Rev. Cancer 2002, 2, 91–100. [Google Scholar] [CrossRef]
- Osborn, L.; Hession, C.; Tizard, R.; Vassallo, C.; Luhowskyj, S.; Chi-Rosso, G.; Lobb, R. Direct expression cloning of vascular cell adhesion molecule 1, a cytokine-induced endothelial protein that binds to lymphocytes. Cell 1989, 59, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Springer, T.A. Traffic signals for lymphocyte recirculation and leukocyte emigration: The multistep paradigm. Cell 1994, 76, 301–314. [Google Scholar] [CrossRef]
- Kong, D.-H.; Kim, Y.K.; Kim, M.R.; Jang, J.H.; Lee, S. Emerging Roles of Vascular Cell Adhesion Molecule-1 (VCAM-1) in Immunological Disorders and Cancer. Int. J. Mol. Sci. 2018, 19, 1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troncoso, M.F.; Ortiz-Quintero, J.; Garrido-Moreno, V.; Sanhueza-Olivares, F.; Guerrero-Moncayo, A.; Chiong, M.; Castro, P.F.; García, L.; Gabrielli, L.; Corbalán, R.; et al. VCAM-1 as a predictor biomarker in cardiovascular disease. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2021, 1867, 166170. [Google Scholar] [CrossRef]
- Gupta, G.P.; Nguyen, D.X.; Chiang, A.C.; Bos, P.D.; Kim, J.Y.; Nadal, C.; Gomis, R.R.; Manova-Todorova, K.; Massagué, J. Mediators of vascular remodelling co-opted for sequential steps in lung metastasis. Nature 2007, 446, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Ruco, L.P.; De Laat, P.A.; Matteucci, C.; Bernasconi, S.; Sciacca, F.M.; Van Der Kwast, T.H.; Hoogsteden, H.C.; Uccini, S.; Mantovani, A.; Versnel, M.A. Expression of ICAM-1 and VCAM-1 in human malignant mesothelioma. J. Pathol. 1996, 179, 266–271. [Google Scholar] [CrossRef]
- Ding, Y.-B.; Chen, G.-Y.; Xia, J.-G.; Zang, X.-W.; Yang, H.-Y.; Yang, L. Association of VCAM-1 overexpression with oncogenesis, tumor angiogenesis and metastasis of gastric carcinoma. World J. Gastroenterol. 2003, 9, 1409–1414. [Google Scholar] [CrossRef]
- Elices, M.J.; Osborn, L.; Takada, Y.; Crouse, C.; Luhowskyj, S.; Hemler, M.E.; Lobb, R.R. VCAM-1 on activated endothelium interacts with the leukocyte integrin VLA-4 at a site distinct from the VLA-4/Fibronectin binding site. Cell 1990, 60, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Mu, E.; Wei, Y.; Riethdorf, S.; Yang, Q.; Yuan, M.; Yan, J.; Hua, Y.; Tiede, B.J.; Lu, X.; et al. VCAM-1 Promotes Osteolytic Expansion of Indolent Bone Micrometastasis of Breast Cancer by Engaging α4β1-Positive Osteoclast Progenitors. Cancer Cell 2011, 20, 701–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Zhang, X.H.-F.; Massagué, J. Macrophage Binding to Receptor VCAM-1 Transmits Survival Signals in Breast Cancer Cells that Invade the Lungs. Cancer Cell 2011, 20, 538–549. [Google Scholar] [CrossRef] [Green Version]
- Rosetti, F.; Mayadas, T.N. The many faces of Mac-1 in autoimmune disease. Immunol. Rev. 2015, 269, 175–193. [Google Scholar] [CrossRef]
- Lamers, C.; Plüss, C.J.; Ricklin, D. The Promiscuous Profile of Complement Receptor 3 in Ligand Binding, Immune Modulation, and Pathophysiology. Front. Immunol. 2021, 12, 662164. [Google Scholar] [CrossRef]
- Asada, M.; Furukawa, K.; Kantor, C.; Gahmberg, C.G.; Kobata, A. Structural study of the sugar chains of human leukocyte cell adhesion molecules CD11/CD18. Biochemistry 1991, 30, 1561–1571. [Google Scholar] [CrossRef]
- Low, M.G.; Kincade, P.W. Phosphatidylinositol is the membrane-anchoring domain of the Thy-1 glycoprotein. Nature 1985, 318, 62–64. [Google Scholar] [CrossRef]
- Wetzel, A.; Chavakis, T.; Preissner, K.T.; Sticherling, M.; Haustein, U.-F.; Anderegg, U.; Saalbach, A. Human Thy-1 (CD90) on Activated Endothelial Cells Is a Counterreceptor for the Leukocyte Integrin Mac-1 (CD11b/CD18). J. Immunol. 2004, 172, 3850–3859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.H.; Dai, Y.D.; Tong, M.; Chan, Y.P.; Kwan, P.S.; Fu, L.; Qin, Y.R.; Tsao, S.W.; Lung, H.L.; Lung, M.L.; et al. A CD90+ Tumor-Initiating Cell Population with an Aggressive Signature and Metastatic Capacity in Esophageal Cancer. Cancer Res. 2013, 73, 2322–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connor, E.V.; Saygin, C.; Braley, C.; Wiechert, A.C.; Karunanithi, S.; Crean-Tate, K.; Abdul-Karim, F.W.; Michener, C.M.; Rose, P.G.; Lathia, J.D.; et al. Thy-1 predicts poor prognosis and is associated with self-renewal in ovarian cancer. J. Ovarian Res. 2019, 12, 112. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, B.; Xiao, S.; Li, Y.; Chen, Q. miR-125a/b inhibits tumor-associated macrophages mediated in cancer stem cells of hepatocellular carcinoma by targeting CD90. J. Cell. Biochem. 2018, 120, 3046–3055. [Google Scholar] [CrossRef]
- Staunton, D.E.; Marlin, S.D.; Stratowa, C.; Dustin, M.L.; Springer, T.A. Primary structure of ICAM-1 demonstrates interaction between members of the immunoglobulin and integrin supergene families. Cell 1988, 52, 925–933. [Google Scholar] [CrossRef]
- Singh, M.; Thakur, M.; Mishra, M.; Yadav, M.; Vibhuti, R.; Menon, A.M.; Nagda, G.; Dwivedi, V.P.; Dakal, T.C.; Yadav, V. Gene regulation of intracellular adhesion molecule-1 (ICAM-1): A molecule with multiple functions. Immunol. Lett. 2021, 240, 123–136. [Google Scholar] [CrossRef]
- Salvador, A.M.; Nevers, T.; Velázquez, F.; Aronovitz, M.; Wang, B.; Molina, A.A.; Jaffe, I.Z.; Karas, R.H.; Blanton, R.M.; Alcaide, P. Intercellular Adhesion Molecule 1 Regulates Left Ventricular Leukocyte Infiltration, Cardiac Remodeling, and Function in Pressure Overload–Induced Heart Failure. J. Am. Hear. Assoc. 2016, 5, e003126. [Google Scholar] [CrossRef] [Green Version]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Rep. 2009, 61, 22–32. [Google Scholar] [CrossRef]
- Regev, O.; Kizner, M.; Roncato, F.; Dadiani, M.; Saini, M.; Castro-Giner, F.; Yajuk, O.; Kozlovski, S.; Levi, N.; Addadi, Y.; et al. ICAM-1 on Breast Cancer Cells Suppresses Lung Metastasis but Is Dispensable for Tumor Growth and Killing by Cytotoxic T Cells. Front. Immunol. 2022, 13, 849701. [Google Scholar] [CrossRef]
- Lynam, E.; Sklar, L.A.; Taylor, A.D.; Neelamegham, S.; Edwards, B.S.; Smith, C.W.; Simon, S.I. β2 -Integrins mediate stable adhesion in collisional interactions between neutrophils and ICAM-1-expressing cells. J. Leukoc. Biol. 1998, 64, 622–630. [Google Scholar] [CrossRef]
- Bai, J.; Adriani, G.; Dang, T.-M.; Tu, T.-Y.; Penny, H.-X.L.; Wong, S.-C.; Kamm, R.D.; Thiery, J.-P. Contact-dependent carcinoma aggregate dispersion by M2a macrophages via ICAM-1 and β2 integrin interactions. Oncotarget 2015, 6, 25295–25307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, M.; Li, X.; Tan, S.; Zhou, H.J.; Ji, W.; Bellone, S.; Xu, X.; Zhang, H.; Santin, A.D.; Lou, G.; et al. Tumor-associated macrophages drive spheroid formation during early transcoelomic metastasis of ovarian cancer. J. Clin. Investig. 2016, 126, 4157–4173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakoff-Nahoum, S.; Medzhitov, R. Toll-like receptors and cancer. Nat. Rev. Cancer 2008, 9, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Seya, T.; Takeda, Y.; Matsumoto, M. A Toll-like receptor 3 (TLR3) agonist ARNAX for therapeutic immunotherapy. Adv. Drug Deliv. Rev. 2019, 147, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Boushehri, M.A.S.; Lamprecht, A. TLR4-Based Immunotherapeutics in Cancer: A Review of the Achievements and Shortcomings. Mol. Pharm. 2018, 15, 4777–4800. [Google Scholar] [CrossRef]
- Michaelis, K.A.; Norgard, M.A.; Zhu, X.; Levasseur, P.R.; Sivagnanam, S.; Liudahl, S.M.; Burfeind, K.G.; Olson, B.; Pelz, K.R.; Ramos, D.M.A.; et al. Publisher Correction: The TLR7/8 agonist R848 remodels tumor and host responses to promote survival in pancreatic cancer. Nat. Commun. 2019, 10, 5257. [Google Scholar] [CrossRef] [Green Version]
- Kapp, K.; Volz, B.; Oswald, D.; Wittig, B.; Schmidt, M. Abstract 2622: Two new TLR9 agonists for cancer immunotherapy: Combination with checkpoint inhibitors. Cancer Res. 2017, 77, 2622. [Google Scholar] [CrossRef]
- Jang, G.-Y.; Lee, J.W.; Kim, Y.S.; Lee, S.E.; Han, H.D.; Hong, K.-J.; Kang, T.H.; Park, Y.-M. Interactions between tumor-derived proteins and Toll-like receptors. Exp. Mol. Med. 2020, 52, 1926–1935. [Google Scholar] [CrossRef]
- Hume, D.A.; MacDonald, K. Therapeutic applications of macrophage colony-stimulating factor-1 (CSF-1) and antagonists of CSF-1 receptor (CSF-1R) signaling. Blood 2012, 119, 1810–1820. [Google Scholar] [CrossRef]
- Achkova, D.; Maher, J. Role of the colony-stimulating factor (CSF)/CSF-1 receptor axis in cancer. Biochem. Soc. Trans. 2016, 44, 333–341. [Google Scholar] [CrossRef]
- Zhu, Y.; Knolhoff, B.L.; Meyer, M.A.; Nywening, T.M.; West, B.L.; Luo, J.; Wang-Gillam, A.; Goedegebuure, S.P.; Linehan, D.C.; DeNardo, D.G. CSF1/CSF1R Blockade Reprograms Tumor-Infiltrating Macrophages and Improves Response to T-cell Checkpoint Immunotherapy in Pancreatic Cancer Models. Cancer Res. 2014, 74, 5057–5069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulus, P.; Stanley, E.R.; Schafer, R.; Abraham, D.; Aharinejad, S. Colony-Stimulating Factor-1 Antibody Reverses Chemoresistance in Human MCF-7 Breast Cancer Xenografts. Cancer Res. 2006, 66, 4349–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gamal, M.I.; Al-Ameen, S.K.; Al-Koumi, D.M.; Hamad, M.G.; Jalal, N.A.; Oh, C.-H. Recent Advances of Colony-Stimulating Factor-1 Receptor (CSF-1R) Kinase and Its Inhibitors. J. Med. Chem. 2018, 61, 5450–5466. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Vilgelm, A.E.; Richmond, A. Chemokines Modulate Immune Surveillance in Tumorigenesis, Metastasis, and Response to Immunotherapy. Front. Immunol. 2019, 10, 333. [Google Scholar] [CrossRef] [Green Version]
- Argyle, D.; Kitamura, T. Targeting Macrophage-Recruiting Chemokines as a Novel Therapeutic Strategy to Prevent the Progression of Solid Tumors. Front. Immunol. 2018, 9, 2629. [Google Scholar] [CrossRef] [Green Version]
- Mortezaee, K.; Majidpoor, J. Roles for macrophage-polarizing interleukins in cancer immunity and immunotherapy. Cell. Oncol. 2022, 45, 333–353. [Google Scholar] [CrossRef]
- Pan, W.; Zhu, S.; Qu, K.; Meeth, K.; Cheng, J.; He, K.; Ma, H.; Liao, Y.; Wen, X.; Roden, C.; et al. The DNA Methylcytosine Dioxygenase Tet2 Sustains Immunosuppressive Function of Tumor-Infiltrating Myeloid Cells to Promote Melanoma Progression. Immunity 2017, 47, 284–297.e5. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, C.; Wang, S.; Shi, D.; Wei, C.; Song, J.; Lin, X.; Dou, R.; Bai, J.; Xiang, Z.; et al. Wnt5a-induced M2 polarization of tumor-associated macrophages via IL-10 promotes colorectal cancer progression. Cell Commun. Signal. 2020, 18, 51. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Chen, T.; Hu, R.; Zhu, R.; Li, C.; Ruan, Y.; Xie, X.; Li, Y. Next frontier in tumor immunotherapy: Macrophage-mediated immune evasion. Biomark. Res. 2021, 9, 72. [Google Scholar] [CrossRef]
- Moesta, A.K.; Li, X.-Y.; Smyth, M.J. Targeting CD39 in cancer. Nat. Rev. Immunol. 2020, 20, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Da, M.; Chen, L.; Enk, A.; Ring, S.; Mahnke, K. The Multifaceted Actions of CD73 During Development and Suppressive Actions of Regulatory T Cells. Front. Immunol. 2022, 13, 914799. [Google Scholar] [CrossRef] [PubMed]
- Perrot, I.; Michaud, H.-A.; Giraudon-Paoli, M.; Augier, S.; Docquier, A.; Gros, L.; Courtois, R.; Déjou, C.; Jecko, D.; Becquart, O.; et al. Blocking antibodies targeting the CD39/CD73 immunosuppressive pathway unleash immune responses in combination cancer therapies. Cell Rep. 2019, 27, 2411–2425.e9. [Google Scholar] [CrossRef] [PubMed]
- Stagg, J.; Divisekera, U.; Duret, H.; Sparwasser, T.; Teng, M.W.; Darcy, P.K.; Smyth, M.J. CD73-Deficient Mice Have Increased Antitumor Immunity and Are Resistant to Experimental Metastasis. Cancer Res. 2011, 71, 2892–2900. [Google Scholar] [CrossRef] [Green Version]
- Ohta, A.; Gorelik, E.; Prasad, S.J.; Ronchese, F.; Lukashev, D.; Wong, M.K.K.; Huang, X.; Caldwell, S.; Liu, K.; Smith, P.; et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13132–13137. [Google Scholar] [CrossRef] [Green Version]
- Ajona, D.; Ortiz-Espinosa, S.; Pio, R. Complement anaphylatoxins C3a and C5a: Emerging roles in cancer progression and treatment. Semin. Cell Dev. Biol. 2019, 85, 153–163. [Google Scholar] [CrossRef]
- Roumenina, L.T.; Daugan, M.V.; Petitprez, F.; Sautès-Fridman, C.; Fridman, W.H. Context-dependent roles of complement in cancer. Nat. Cancer 2019, 19, 698–715. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; Chintala, N.K.; Vadrevu, S.K.; Patel, J.; Karbowniczek, M.; Markiewski, M.M. Pulmonary Alveolar Macrophages Contribute to the Premetastatic Niche by Suppressing Antitumor T Cell Responses in the Lungs. J. Immunol. 2015, 194, 5529–5538. [Google Scholar] [CrossRef] [Green Version]
- Ajona, D.; Ortiz-Espinosa, S.; Moreno, H.; Lozano, T.; Pajares, M.J.; Agorreta, J.; Bértolo, C.; Lasarte, J.J.; Vicent, S.; Hoehlig, K.; et al. A Combined PD-1/C5a Blockade Synergistically Protects against Lung Cancer Growth and Metastasis. Cancer Discov. 2017, 7, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-L.; Zhao, H.; Ren, X.-B. Relationship of VEGF/VEGFR with immune and cancer cells: Staggering or forward? Cancer Biol. Med. 2016, 13, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Gavalas, N.G.; Tsiatas, M.; Tsitsilonis, O.; Politi, E.; Ioannou, K.; Ziogas, A.C.; Rodolakis, A.; Vlahos, G.; Thomakos, N.; Haidopoulos, D.; et al. VEGF directly suppresses activation of T cells from ascites secondary to ovarian cancer via VEGF receptor type 2. Br. J. Cancer 2012, 107, 1869–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, T.E.; Kirkpatrick, N.D.; Huang, Y.; Farrar, C.T.; Marijt, K.A.; Kloepper, J.; Datta, M.; Amoozgar, Z.; Seano, G.; Jung, K.; et al. Dual inhibition of Ang-2 and VEGF receptors normalizes tumor vasculature and prolongs survival in glioblastoma by altering macrophages. Proc. Natl. Acad. Sci. USA 2016, 113, 4470–4475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flavell, R.A.; Sanjabi, S.; Wrzesinski, S.H.; Licona-Limón, P. The polarization of immune cells in the tumour environment by TGFbeta. Nat. Rev. Immunol. 2010, 10, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2012, 229, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Faivre, S.; Santoro, A.; Kelley, R.K.; Gane, E.; Costentin, C.E.; Gueorguieva, I.; Smith, C.; Cleverly, A.; Lahn, M.M.; Raymond, E.; et al. Novel transforming growth factor beta receptor I kinase inhibitor galunisertib (LY2157299) in advanced hepatocellular carcinoma. Liver Int. 2019, 39, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.; O’Neill, L.A. Succinate: A metabolic signal in inflammation. Trends Cell Biol. 2014, 24, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.-Y.; Huang, T.-W.; Hsieh, Y.-T.; Wang, Y.-F.; Yen, C.-C.; Lee, G.-L.; Yeh, C.-C.; Peng, Y.-J.; Kuo, Y.-Y.; Wen, H.-T.; et al. Cancer-Derived Succinate Promotes Macrophage Polarization and Cancer Metastasis via Succinate Receptor. Mol. Cell 2020, 77, 213–227.e5. [Google Scholar] [CrossRef]
- Dobosz, P.; Stempor, P.A.; Roszik, J.; Herman, A.; Layani, A.; Berger, R.; Avni, D.; Sidi, Y.; Leibowitz-Amit, R. Checkpoint Genes at the Cancer Side of the Immunological Synapse in Bladder Cancer. Transl. Oncol. 2020, 13, 193–200. [Google Scholar] [CrossRef]
- Stempor, P.; Avni, D.; Leibowitz, R.; Sidi, Y.; Stępień, M.; Dzieciątkowski, T.; Dobosz, P. Comprehensive Analysis of Correlations in the Expression of miRNA Genes and Immune Checkpoint Genes in Bladder Cancer Cells. Int. J. Mol. Sci. 2021, 22, 2553. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Immunomodulators Type | Drug Name | Trade Name | Target | Representative Indications | Approved Date |
|---|---|---|---|---|---|
| Checkpoint inhibitors | Ipilimumab | Yervoy | CTLA-4 | Melanoma | 2011.03 |
| Pembrolizumab | Keytruda | PD-1/PD-L1 | Melanoma | 2014.09 | |
| Nivolumab | Opdivo | PD-1/PD-L1 | Esophageal carcinoma | 2014.12 | |
| Avelumab | Bavencio | PD-1/PD-L1 | Merkel cell tumor | 2017.03 | |
| Durvalumab | Imfinzi | PD-1/PD-L1 | Transitional cell carcinoma | 2017.05 | |
| Cemiplimab | Libtayo | PD-1/PD-L1 | Cutaneous squamous cell carcinoma | 2018.09 | |
| Dostarlimab | Jemperli | PD-1 | Endometrial carcinoma | 2021.08 | |
| Relatlimab | Opdualag (In combination with nivolumab) | LAG-3 | Melanoma | 2022.03 | |
| Cytokines | Interferon alfa-2a | NA | IFNAR1/2 | Chronic myelogenous leukemia | 1986.06 (Withdraw) |
| Interferon alfa-2b | Intron A | IFNAR1/2 | Hairy cell leukemia | 1986.06 | |
| rGM-CSF | Sargramostim | CSF2R | Leukemia | 1991.03 | |
| Aldesleukin | Proleukin | IL-2/IL-2R | Renal cell carcinoma | 1992.05 | |
| Peginterferon alfa-2b | Peg-Intron | IFNAR1 | Melanoma | 2011.03 | |
| Small molecules | Imiquimod | ALDARA | TLR7 | Basal cell carcinoma | 2004.07 |
| Pexidartinib | Turalio | KIT, CSF1R, and FLT3 | Giant cell tumor of tendon sheath | 2019.08 | |
| Rintatolimod | Rintamod | TLR3 | Renal cell carcinoma | 2021.06 |
| Strategy | TAM-Target | Intervention | Clinical Phase | Tumor Type | Combination | ClinicalTrials.gov Identifier |
|---|---|---|---|---|---|---|
| TAM reprogramming | CD40 | RO7009789 (Selicrelumab) | I (completed) | Solid tumors | Atezolizumab (anti-PD-L1 mAb) | NCT02304393 |
| APX005M (Sotigalimab) | II (ongoing) | Unresectable and metastatic melanoma | NA | NCT04337931 | ||
| ADC-1013 (Mitazalimab) | I (completed) | Solid tumors | NA | NCT02379741 | ||
| TLR7 | Imiquimod | I/II (completed) | Breast cancer | Radiation, Cyclophosphamide | NCT01421017 | |
| TLR8 | VTX-2337 | I (completed) | Epithelial ovarian cancer, Fallopian tube cancer | Pegylated liposomal doxorubicin | NCT01666444 | |
| TLR9 | SD-101 | I/II (completed) | Lymphoma | Epacadostat, Radiation | NCT03322384 | |
| CSF-1R | Cabiralizumab | I (completed) | Advanced solid tumors | Nivolumab (anti-PD-1 mAb) | NCT02526017 | |
| FPA008 | I/II (completed) | Tenosynovial giant cell tumors | NA | NCT02471716 | ||
| Macrophage-mediated apoptosis | TRAIL-R | Mapatumumab | I/II (completed) | Advanced hepatocellular carcinoma | Sorafenib | NCT01258608 |
| II (completed) | Advanced hepatocellular carcinoma | Bortezomib | NCT00315757 | |||
| II (completed) | NSCLC * | Paclitaxel, Carboplatin | NCT00583830 | |||
| Macrophage-mediated phagocytosis | SLAMF7 | Elotuzumab | II (completed) | MM * | Lenalidomide | NCT03411031 |
| III (completed) | Lymphoma, MM | Lenalidmide, Deamthasone | NCT01239797 | |||
| III (completed) | MM | Nivolumab (anti-PD-1 mAb), Pomalidmide, Deamthasone | NCT02726581 | |||
| CD47 | Hu5F9-G4 (Magrolimab) | III (ongoing) | Acute Myeloid Leukemia | Venetoclax, Azacitidine | NCT05079230 | |
| CC-90002 | I (completed) | Hematologic neoplasms | Rituximab (anti-CD20 mAb) | NCT02367196 | ||
| SIRPα | TTI-621 | I (ongoing) | Hematologic malignancies, Solid tumor | Rituximab (anti-CD20 mAb), Nivolumab (anti-PD-1 mAb) | NCT02663518 | |
| LILRB1 | AGEN1571 | I (ongoing) | Advanced solid tumors | Balstilimab (anti-PD-1 mAb), Botensilimab (anti-CTLA-4 mAb) | NCT05377528 | |
| PtdSer | Bavituximab | II (completed) | Metastatic oancreatic cancer | Gemcitabine | NCT01272791 | |
| II (completed) | Non-squamous NSCLC | Paclitaxel/Carboplatin | NCT01160601 | |||
| MerTk | MRX-2843 | I (ongoing) | NSCLC | Osimertinib | NCT04762199 | |
| Siglec10 | alemtuzumab | III (completed) | B cell chronic lymphocytic leukemia | NA | NCT00046683 | |
| III (completed) | T-cell lymphoma | CHOP14 chemotherapy | NCT00646854 | |||
| Inhibiting the immuno-suppressive activity | PD-1 | SHR-121 (Camrelizumab) | III (completed) | Non-squmous NSCLC | Carboplatin, Pemetrexed | NCT03134872 |
| PD-L1 | Atezolizumab | II (completed) | Advanced solid tumors | NA | NCT02458638 | |
| IV (completed) | NSCLC | NA | NCT03285763 | |||
| Siglec15 | NC318 | I/II (ongoing) | Advanced or metastatic solid tumors | NA | NCT03665285 | |
| II (ongoing) | Advanced NSCLC | Pembrolizumab (anti-PD-1 mAb) | NCT04699123 | |||
| EGFR | Erlotinib | IV (completed) | Non-squmous NSCLC | NA | NCT01609543 | |
| Galectin9 | LYT-200 | I/II (ongoing) | Metastatic solid tumors | Chemotherapy, anti PD-1 inhibitor | NCT04666688 | |
| CD39 | JS019 | I (ongoing) | Advanced solid tumors, Lymphomas | NA | NCT05374226 | |
| CD73 | AK119 | I (ongoing) | Advanced or metastatic solid tumors | Cadonilimab (anti-PD-1/CTLA-4 mAb) | NCT04572152 | |
| AR2A | Ciforadenant | I (completed) | Renal cell cancer, Metastatic castration resistant prostate cacer | Atezolizumab (anti-PD-L1 mAb) | NCT02655822 | |
| C5aR | TJ210001 | I (ongoing) | Advanced solid tumor | NA | NCT04947033 | |
| I (ongoing) | Relapsed or refractory advanced solid tumors | NA | NCT04678921 | |||
| TGF-βR | LY2157299 | II (completed) | Hepatocelllar carcinoma | Sorafenib | NCT02178358 | |
| AVID200 | I (ongoing) | Hepatocelllar carcinoma | NA | NCT01025206 | ||
| TAMs recruitment | ICAM-1 | BI-505 | I (completed) | MM | NA | NCT01025206 |
| CCR2/CCR5 | BMS-813160 | II (ongoing) | NSCLC, Hepatocellular Carcinoma | Nivolumab (anti-PD-1 mAb), BMS-986253 (anti-IL-8 mAb) | NCT04123379 | |
| CCR2 | MLN1202 | II (completed) | Metastatic cancer | NA | NCT01015560 | |
| CXCR4 | BL-8040 | II (completed) | Acute myloid leukemia | Cytarabine | NCT01838395 | |
| IL-1 | Anakinra | II (completed) | Multiple myeloma, Plasma cell neoplasm | Dexamethaso-ne acetate | NCT00635154 | |
| I (completed) | Metastatic/Refractory cancer | Denosumab, Everolimus | NCT01624766 | |||
| VEGFR | HMPL-013 (Fruquintinib) | III (ongoing) | Metastatic colorectal cancer | NA | NCT04322539 | |
| Avastin (Bevacizumab) | IV (completed) | Non-squmous NSCLC | Platinum-based chemotherapy | NCT00451906 |
| Target | Drug Name | Trade Name | Representative Indications | Approved Date |
|---|---|---|---|---|
| VEGF/VEGFR | Bevacizumab | Avastin | Colorectal cancer | 2004.02 |
| Ramucirumab | Cyramza | Stomach cancer | 2014.04 | |
| EGFR | Cetuximab | Erbitux | Colorectal cancer | 2004.02 |
| Panitumumab | Vectibix | Colorectal cancer | 2006.09 | |
| Necitumumab | Portrazza | NSCLC * | 2015.11 | |
| CD38 | Daratumumab | Darzalex | MM * | 2015.11 |
| Isatuximab | Sarclisa | MM | 2020.03 | |
| SLAMF7 | Elotuzumab | Empliciti | MM | 2015.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhang, W.; Lan, P. Immune Checkpoint and Other Receptor-Ligand Pairs Modulating Macrophages in Cancer: Present and Prospects. Cancers 2022, 14, 5963. https://doi.org/10.3390/cancers14235963
Yang Y, Zhang W, Lan P. Immune Checkpoint and Other Receptor-Ligand Pairs Modulating Macrophages in Cancer: Present and Prospects. Cancers. 2022; 14(23):5963. https://doi.org/10.3390/cancers14235963
Chicago/Turabian StyleYang, Yuanyuan, Weijie Zhang, and Peixiang Lan. 2022. "Immune Checkpoint and Other Receptor-Ligand Pairs Modulating Macrophages in Cancer: Present and Prospects" Cancers 14, no. 23: 5963. https://doi.org/10.3390/cancers14235963
APA StyleYang, Y., Zhang, W., & Lan, P. (2022). Immune Checkpoint and Other Receptor-Ligand Pairs Modulating Macrophages in Cancer: Present and Prospects. Cancers, 14(23), 5963. https://doi.org/10.3390/cancers14235963