
Novel Insights on the Use of L-Asparaginase as an Efficient and Safe Anti-Cancer Therapy

 ,
,  ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Bacterial L-ASNase Used in the Clinic for ALL Therapy
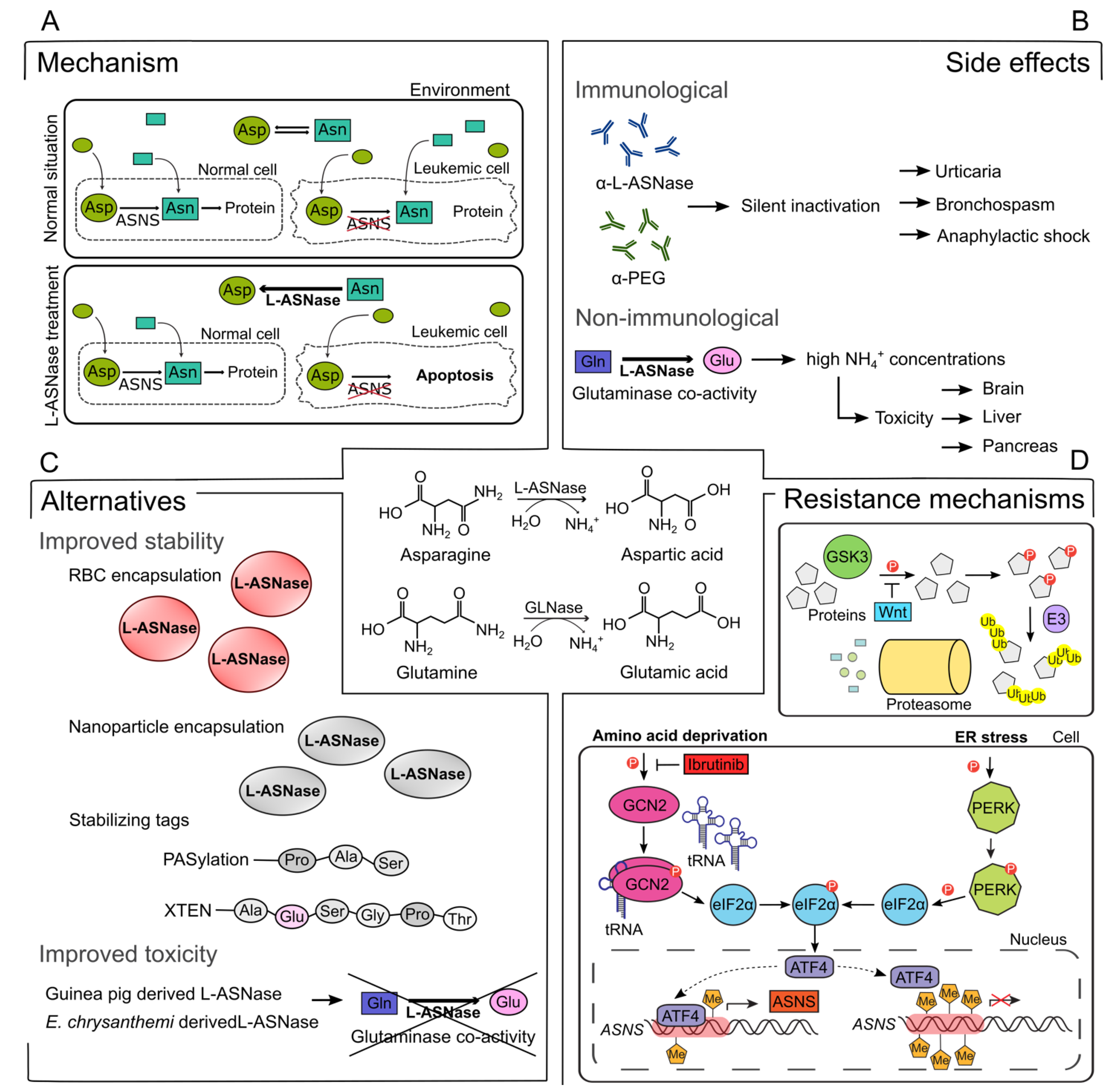
2.1. Immunological Side Effects
2.2. Non-Immunological Side Effects
3. ASNS Promoter Methylation as Biomarker for L-ASNase Sensitivity in ALL
4. Expanding the Use of L-ASNase Therapy beyond ALL in Children
4.1. Acute Monocytic Leukemia
4.2. Solid Cancers
4.3. Glioblastoma
5. The Development of Novel L-ASNase Variants
5.1. L-ASNase Red Blood Cell Encapsulation
5.2. L-ASNase Variants with Reduced or Absent Co-Activity
5.3. Enzyme Engineering and Bioprospecting
6. Combination Therapies
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Choi, B.H.; Coloff, J.L. The Diverse Functions of Non-Essential Amino Acids in Cancer. Cancers 2019, 11, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endicott, M.; Jones, M.; Hull, J. Amino acid metabolism as a therapeutic target in cancer: A review. Amino Acids 2021, 53, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Batra, S.; Zhang, J. Asparagine: A Metabolite to Be Targeted in Cancers. Metabolites 2021, 11, 402. [Google Scholar] [CrossRef] [PubMed]
- Lomelino, C.L.; Andring, J.T.; McKenna, R.; Kilberg, M.S. Asparagine synthetase: Function, structure, and role in disease. J. Biol. Chem. 2017, 292, 19952–19958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, S. Uber desamidierung im Tierkorper. Beitr. Chem. Physiol. Pathol. 1904, 5, 321–345. [Google Scholar]
- Broome, J.D. Evidence that the L-asparaginase of guinea pig serum is responsible for its antilymphoma effects. I. Properties of the L-asparaginase of guinea pig serum in relation to those of the antilymphoma substance. J. Exp. Med. 1963, 118, 99–120. [Google Scholar] [CrossRef] [Green Version]
- Kidd, J.G. Regression of transplanted lymphomas induced in vivo by means of normal guinea pig serum. I. Course of transplanted cancers of various kinds in mice and rats given guinea pig serum, horse serum, or rabbit serum. J. Exp. Med. 1953, 98, 565–582. [Google Scholar] [CrossRef]
- Dolowy, W.C.; Henson, D.; Cornet, J.; Sellin, H. Toxic and antineoplastic effects of L-asparaginase. Study of mice with lymphoma and normal monkeys and report on a child with leukemia. Cancer 1966, 19, 1813–1819. [Google Scholar] [CrossRef]
- Batool, T.; Makky, E.A.; Jalal, M.; Yusoff, M.M. A Comprehensive Review on L-Asparaginase and Its Applications. Appl. Biochem. Biotechnol. 2016, 178, 900–923. [Google Scholar] [CrossRef] [Green Version]
- Cachumba, J.J.; Antunes, F.A.; Peres, G.F.; Brumano, L.P.; Santos, J.C.; Da Silva, S.S. Current applications and different approaches for microbial l-asparaginase production. Braz. J. Microbiol. 2016, 47 (Suppl. S1), 77–85. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Kaur, J.; Walia, S.; Pathak, T.; Aggarwal, D. L-asparaginase: An effective agent in the treatment of acute lymphoblastic leukemia. Leuk. Lymphoma 2014, 55, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Richards, N.G.; Kilberg, M.S. Asparagine synthetase chemotherapy. Annu. Rev. Biochem. 2006, 75, 629–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prager, M.D.; Bachynsky, N. Asparagine synthetase in normal and malignant tissues: Correlation with tumor sensitivity to asparaginase. Arch. Biochem. Biophys. 1968, 127, 645–654. [Google Scholar] [CrossRef]
- Haskell, C.M.; Canellos, G.P. l-asparaginase resistance in human leukemia--asparagine synthetase. Biochem. Pharm. 1969, 18, 2578–2580. [Google Scholar] [CrossRef]
- Chiu, M.; Taurino, G.; Bianchi, M.G.; Kilberg, M.S.; Bussolati, O. Asparagine Synthetase in Cancer: Beyond Acute Lymphoblastic Leukemia. Front. Oncol. 2019, 9, 1480. [Google Scholar] [CrossRef]
- Emadi, A.; Zokaee, H.; Sausville, E.A. Asparaginase in the treatment of non-ALL hematologic malignancies. Cancer Chemother. Pharm. 2014, 73, 875–883. [Google Scholar] [CrossRef]
- Dufour, E.; Gay, F.; Aguera, K.; Scoazec, J.Y.; Horand, F.; Lorenzi, P.L.; Godfrin, Y. Pancreatic tumor sensitivity to plasma L-asparagine starvation. Pancreas 2012, 41, 940–948. [Google Scholar] [CrossRef]
- Lorenzi, P.L.; Reinhold, W.C.; Rudelius, M.; Gunsior, M.; Shankavaram, U.; Bussey, K.J.; Scherf, U.; Eichler, G.S.; Martin, S.E.; Chin, K.; et al. Asparagine synthetase as a causal, predictive biomarker for L-asparaginase activity in ovarian cancer cells. Mol. Cancer 2006, 5, 2613–2623. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Henning, R.; Walker, A.; Kim, G.; Perroy, A.; Alessandro, R.; Virador, V.; Kohn, E.C. L-asparaginase inhibits invasive and angiogenic activity and induces autophagy in ovarian cancer. J. Cell Mol. Med. 2012, 16, 2369–2378. [Google Scholar] [CrossRef]
- Knott, S.R.V.; Wagenblast, E.; Khan, S.; Kim, S.Y.; Soto, M.; Wagner, M.; Turgeon, M.O.; Fish, L.; Erard, N.; Gable, A.L.; et al. Asparagine bioavailability governs metastasis in a model of breast cancer. Nature 2018, 554, 378–381. [Google Scholar] [CrossRef]
- Hijiya, N.; van der Sluis, I.M. Asparaginase-associated toxicity in children with acute lymphoblastic leukemia. Leuk. Lymphoma 2016, 57, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Thu Huynh, V.; Bergeron, S. Asparaginase Toxicities: Identification and Management in Patients with Acute Lymphoblastic Leukemia. Clin. J. Oncol. Nurs. 2017, 21, E248–E259. [Google Scholar] [CrossRef] [PubMed]
- Pieters, R.; Hunger, S.P.; Boos, J.; Rizzari, C.; Silverman, L.; Baruchel, A.; Goekbuget, N.; Schrappe, M.; Pui, C.H. L-asparaginase treatment in acute lymphoblastic leukemia: A focus on Erwinia asparaginase. Cancer 2011, 117, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maese, L.; Rizzari, C.; Coleman, R.; Power, A.; van der Sluis, I.; Rau, R.E. Can recombinant technology address asparaginase Erwinia chrysanthemi shortages? Pediatr. Blood Cancer 2021, 68, e29169. [Google Scholar] [CrossRef] [PubMed]
- Kawedia, J.D.; Rytting, M.E. Asparaginase in acute lymphoblastic leukemia. Clin. Lymphoma Myeloma Leuk. 2014, 14, S14–S17. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.A.; Syed, Y.Y.; Keam, S.J. Pegaspargase: A Review in Acute Lymphoblastic Leukaemia. Drugs 2019, 79, 767–777. [Google Scholar] [CrossRef] [Green Version]
- Lew, G. Space for Calaspargase? A New Asparaginase for Acute Lymphoblastic Leukemia. Clin. Cancer Res. 2020, 26, 325–327. [Google Scholar] [CrossRef] [Green Version]
- Angiolillo, A.L.; Schore, R.J.; Devidas, M.; Borowitz, M.J.; Carroll, A.J.; Gastier-Foster, J.M.; Heerema, N.A.; Keilani, T.; Lane, A.R.; Loh, M.L.; et al. Pharmacokinetic and pharmacodynamic properties of calaspargase pegol Escherichia coli L-asparaginase in the treatment of patients with acute lymphoblastic leukemia: Results from Children’s Oncology Group Study AALL07P4. J. Clin. Oncol. 2014, 32, 3874–3882. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.L.; Guimarães, G.M.; Polakiewicz, B.; de Moraes Pitombo, R.N.; Abrahão-Neto, J. Effects of polyethylene glycol attachment on physicochemical and biological stability of E. coli L-asparaginase. Int. J. Pharm. 2002, 237, 163–170. [Google Scholar] [CrossRef]
- Cecconello, D.K.; Magalhães, M.R.; Werlang, I.C.R.; Lee, M.L.M.; Michalowski, M.B.; Daudt, L.E. Asparaginase: An old drug with new questions. Hematol. Transfus. Cell 2020, 42, 275–282. [Google Scholar] [CrossRef]
- Brumano, L.P.; da Silva, F.V.S.; Costa-Silva, T.A.; Apolinário, A.C.; Santos, J.H.P.M.; Kleingesinds, E.K.; Monteiro, G.; Rangel-Yagui, C.d.O.; Benyahia, B.; Junior, A.P. Development of L-Asparaginase Biobetters: Current Research Status and Review of the Desirable Quality Profiles. Front. Bioeng. Biotechnol. 2019, 6, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, C.; Maese, L.; Carter-Febres, M.; Verma, A. Clinical Utility of Pegaspargase in Children, Adolescents and Young Adult Patients with Acute Lymphoblastic Leukemia: A Review. Blood Lymphat. Cancer 2021, 11, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.J.; Boos, J. Use of l-asparaginase in childhood ALL. Crit. Rev. Oncol./Hematol. 1998, 28, 97–113. [Google Scholar] [CrossRef]
- Mondelaers, V.; Ferster, A.; Uyttebroeck, A.; Brichard, B.; van der Werff Ten Bosch, J.; Norga, K.; Francotte, N.; Piette, C.; Vandemeulebroecke, K.; Verbeke, C.; et al. Prospective, real-time monitoring of pegylated Escherichia coli and Erwinia asparaginase therapy in childhood acute lymphoblastic leukaemia and non-Hodgkin lymphoma in Belgium. Br. J. Haematol. 2020, 190, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, C.; Stewart, E.; Panetta, J.; Wilkinson, M.; Morrison, A.; Finkelman, F.; Sandlund, J.; Pui, C.; Jeha, S.; Relling, M. Successful challenges using native E. coli asparaginase after hypersensitivity reactions to PEGylated E. coli asparaginase. Cancer Chemother. Pharmacol. 2014, 73, 1307–1313. [Google Scholar] [CrossRef] [Green Version]
- Ramya, L.N.; Doble, M.; Rekha, V.P.; Pulicherla, K.K. L-Asparaginase as potent anti-leukemic agent and its significance of having reduced glutaminase side activity for better treatment of acute lymphoblastic leukaemia. Appl. Biochem. Biotechnol. 2012, 167, 2144–2159. [Google Scholar] [CrossRef]
- Völler, S.; Pichlmeier, U.; Zens, A.; Hempel, G. Pharmacokinetics of recombinant asparaginase in children with acute lymphoblastic leukemia. Cancer Chemother. Pharmacol. 2018, 81, 305–314. [Google Scholar] [CrossRef]
- Chan, W.K.; Lorenzi, P.L.; Anishkin, A.; Purwaha, P.; Rogers, D.M.; Sukharev, S.; Rempe, S.B.; Weinstein, J.N. The glutaminase activity of L-asparaginase is not required for anticancer activity against ASNS-negative cells. Blood 2014, 123, 3596–3606. [Google Scholar] [CrossRef] [Green Version]
- Parmentier, J.H.; Maggi, M.; Tarasco, E.; Scotti, C.; Avramis, V.I.; Mittelman, S.D. Glutaminase activity determines cytotoxicity of L-asparaginases on most leukemia cell lines. Leuk. Res. 2015, 39, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.A.; Su, Y.; Zhang, J.Y.; Antanasijevic, A.; Caffrey, M.; Schalk, A.M.; Liu, L.; Rondelli, D.; Oh, A.; Mahmud, D.L.; et al. A Novel l-Asparaginase with low l-Glutaminase Coactivity Is Highly Efficacious against Both T- and B-cell Acute Lymphoblastic Leukemias In Vivo. Cancer Res. 2018, 78, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.K.; Horvath, T.D.; Tan, L.; Link, T.; Harutyunyan, K.G.; Pontikos, M.A.; Anishkin, A.; Du, D.; Martin, L.A.; Yin, E.; et al. Glutaminase Activity of L-Asparaginase Contributes to Durable Preclinical Activity against Acute Lymphoblastic Leukemia. Mol. Cancer 2019, 18, 1587–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Sluis, I.M.; Vrooman, L.M.; Pieters, R.; Baruchel, A.; Escherich, G.; Goulden, N.; Mondelaers, V.; Sanchez de Toledo, J.; Rizzari, C.; Silverman, L.B.; et al. Consensus expert recommendations for identification and management of asparaginase hypersensitivity and silent inactivation. Haematologica 2016, 101, 279–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinnick, S.E.; Browning, M.L.; Koontz, S.E. Managing hypersensitivity to asparaginase in pediatrics, adolescents, and young adults. J. Pediatr. Oncol. Nurs. 2013, 30, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Pui, C.H.; Liu, Y.; Relling, M.V. How to solve the problem of hypersensitivity to asparaginase? Pediatr. Blood Cancer 2018, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Højfeldt, S.G.; Wolthers, B.O.; Tulstrup, M.; Abrahamsson, J.; Gupta, R.; Harila-Saari, A.; Heyman, M.; Henriksen, L.T.; Jónsson, Ò.G.; Lähteenmäki, P.M.; et al. Genetic predisposition to PEG-asparaginase hypersensitivity in children treated according to NOPHO ALL2008. Br. J. Haematol. 2019, 184, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, L.T.; Harila-Saari, A.; Ruud, E.; Abrahamsson, J.; Pruunsild, K.; Vaitkeviciene, G.; Jónsson, Ó.G.; Schmiegelow, K.; Heyman, M.; Schrøder, H.; et al. PEG-asparaginase allergy in children with acute lymphoblastic leukemia in the NOPHO ALL2008 protocol. Pediatr. Blood Cancer 2015, 62, 427–433. [Google Scholar] [CrossRef]
- Schore, R.J.; Devidas, M.; Bleyer, A.; Reaman, G.H.; Winick, N.; Loh, M.L.; Raetz, E.A.; Carroll, W.L.; Hunger, S.P.; Angiolillo, A.L. Plasma asparaginase activity and asparagine depletion in acute lymphoblastic leukemia patients treated with pegaspargase on Children’s Oncology Group AALL07P4. Leuk. Lymphoma 2019, 60, 1740–1748. [Google Scholar] [CrossRef]
- Salzer, W.; Bostrom, B.; Messinger, Y.; Perissinotti, A.J.; Marini, B. Asparaginase activity levels and monitoring in patients with acute lymphoblastic leukemia. Leuk. Lymphoma 2018, 59, 1797–1806. [Google Scholar] [CrossRef]
- Tong, W.H.; Pieters, R.; Kaspers, G.J.; te Loo, D.M.; Bierings, M.B.; van den Bos, C.; Kollen, W.J.; Hop, W.C.; Lanvers-Kaminsky, C.; Relling, M.V.; et al. A prospective study on drug monitoring of PEGasparaginase and Erwinia asparaginase and asparaginase antibodies in pediatric acute lymphoblastic leukemia. Blood 2014, 123, 2026–2033. [Google Scholar] [CrossRef] [Green Version]
- Yen, H.J.; Chang, W.H.; Liu, H.C.; Yeh, T.C.; Hung, G.Y.; Wu, K.H.; Peng, C.T.; Chang, Y.H.; Chang, T.K.; Hsiao, C.C.; et al. Outcomes Following Discontinuation of E. coli l-Asparaginase Upon Severe Allergic Reactions in Children With Acute Lymphoblastic Leukemia. Pediatr. Blood Cancer 2016, 63, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Avramis, V.I.; Panosyan, E.H. Pharmacokinetic/pharmacodynamic relationships of asparaginase formulations: The past, the present and recommendations for the future. Clin. Pharm. 2005, 44, 367–393. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.K.; Hempel, G.; Koling, S.; Chan, L.S.; Fisher, T.; Meiselman, H.J.; Garratty, G. Antibody against poly(ethylene glycol) adversely affects PEG-asparaginase therapy in acute lymphoblastic leukemia patients. Cancer 2007, 110, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Kloos, R.; van der Sluis, I.M.; Mastrobattista, E.; Hennink, W.; Pieters, R.; Verhoef, J.J. Acute lymphoblastic leukaemia patients treated with PEGasparaginase develop antibodies to PEG and the succinate linker. Br. J. Haematol. 2020, 189, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Song, M.; Sim, B.C.; Gu, C.; Podust, V.N.; Wang, C.W.; McLaughlin, B.; Shah, T.P.; Lax, R.; Gast, R.; et al. Multivalent antiviral XTEN-peptide conjugates with long in vivo half-life and enhanced solubility. Bioconjug. Chem. 2014, 25, 1351–1359. [Google Scholar] [CrossRef] [Green Version]
- Schlapschy, M.; Binder, U.; Börger, C.; Theobald, I.; Wachinger, K.; Kisling, S.; Haller, D.; Skerra, A. PASylation: A biological alternative to PEGylation for extending the plasma half-life of pharmaceutically active proteins. Protein Eng. Des. Sel. 2013, 26, 489–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebauer, M.; Skerra, A. Prospects of PASylation® for the design of protein and peptide therapeutics with extended half-life and enhanced action. Bioorg. Med. Chem. 2018, 26, 2882–2887. [Google Scholar] [CrossRef]
- Ortac, I.; Ruff, L.; Yeh, Y.; Esener, S.; Messmer, B.T. Nanoparticle Encapsulated L-Asparaginase. Blood 2013, 122, 2669. [Google Scholar] [CrossRef]
- Teodor, E.; Litescu, S.-C.; Lazar, V.; Somoghi, R. Hydrogel-magnetic nanoparticles with immobilized l-asparaginase for biomedical applications. J. Mater. Sci. Mater. Med. 2009, 20, 1307–1314. [Google Scholar] [CrossRef]
- Woo, M.H.; Hak, L.J.; Storm, M.C.; Evans, W.E.; Sandlund, J.T.; Rivera, G.K.; Wang, B.; Pui, C.H.; Relling, M.V. Anti-asparaginase antibodies following E. coli asparaginase therapy in pediatric acute lymphoblastic leukemia. Leukemia 1998, 12, 1527–1533. [Google Scholar] [CrossRef] [Green Version]
- Zalewska-Szewczyk, B.; Andrzejewski, W.; Młynarski, W.; Jedrychowska-Dańska, K.; Witas, H.; Bodalski, J. The anti-asparagines antibodies correlate with L-asparagines activity and may affect clinical outcome of childhood acute lymphoblastic leukemia. Leuk. Lymphoma 2007, 48, 931–936. [Google Scholar] [CrossRef]
- Vrooman, L.M.; Kirov, I.I.; Dreyer, Z.E.; Kelly, M.; Hijiya, N.; Brown, P.; Drachtman, R.A.; Messinger, Y.H.; Ritchey, A.K.; Hale, G.A.; et al. Activity and Toxicity of Intravenous Erwinia Asparaginase Following Allergy to E. coli-Derived Asparaginase in Children and Adolescents With Acute Lymphoblastic Leukemia. Pediatr. Blood Cancer 2016, 63, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Salzer, W.L.; Asselin, B.; Supko, J.G.; Devidas, M.; Kaiser, N.A.; Plourde, P.; Winick, N.J.; Reaman, G.H.; Raetz, E.; Carroll, W.L.; et al. Erwinia asparaginase achieves therapeutic activity after pegaspargase allergy: A report from the Children’s Oncology Group. Blood 2013, 122, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Wang, C.; Raetz, E.A.; Schore, R.; Salzer, W.L.; Larsen, E.C.; Maloney, K.W.; Mattano, L.A., Jr.; Carroll, W.L.; Winick, N.J.; et al. Impact of Asparaginase Discontinuation on Outcome in Childhood Acute Lymphoblastic Leukemia: A Report From the Children’s Oncology Group. J. Clin. Oncol. 2020, 38, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Panosyan, E.H.; Seibel, N.L.; Martin-Aragon, S.; Gaynon, P.S.; Avramis, I.A.; Sather, H.; Franklin, J.; Nachman, J.; Ettinger, L.J.; La, M.; et al. Asparaginase antibody and asparaginase activity in children with higher-risk acute lymphoblastic leukemia: Children’s Cancer Group Study CCG-1961. J. Pediatr. Hematol. Oncol. 2004, 26, 217–226. [Google Scholar] [CrossRef]
- Silverman, L.B.; Gelber, R.D.; Dalton, V.K.; Asselin, B.L.; Barr, R.D.; Clavell, L.A.; Hurwitz, C.A.; Moghrabi, A.; Samson, Y.; Schorin, M.A.; et al. Improved outcome for children with acute lymphoblastic leukemia: Results of Dana-Farber Consortium Protocol 91-01. Blood 2001, 97, 1211–1218. [Google Scholar] [CrossRef]
- Liu, C.; Kawedia, J.D.; Cheng, C.; Pei, D.; Fernandez, C.A.; Cai, X.; Crews, K.R.; Kaste, S.C.; Panetta, J.C.; Bowman, W.P.; et al. Clinical utility and implications of asparaginase antibodies in acute lymphoblastic leukemia. Leukemia 2012, 26, 2303–2309. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk Højfeldt, S.; Grell, K.; Abrahamsson, J.; Lund, B.; Vettenranta, K.; Jónsson, Ó.G.; Frandsen, T.L.; Wolthers, B.O.; Marquart, H.V.; Vaitkeviciene, G.; et al. Relapse risk following truncation of pegylated asparaginase in childhood acute lymphoblastic leukemia. Blood 2021, 137, 2373–2382. [Google Scholar] [CrossRef]
- Lin, T.; Hernandez-Illas, M.; Rey, A.; Jenkins, J.; Chandula, R.; Silverman, J.A.; Choi, M.R. A Randomized Phase I Study to Evaluate the Safety, Tolerability, and Pharmacokinetics of Recombinant Erwinia Asparaginase (JZP-458) in Healthy Adult Volunteers. Clin. Transl. Sci. 2021, 14, 870–879. [Google Scholar] [CrossRef]
- Lin, T.; Dumas, T.; Kaullen, J.; Berry, N.S.; Choi, M.R.; Zomorodi, K.; Silverman, J.A. Population Pharmacokinetic Model Development and Simulation for Recombinant Erwinia Asparaginase Produced in Pseudomonas fluorescens (JZP-458). Clin. Pharm. Drug Dev. 2021, 10, 1503–1513. [Google Scholar] [CrossRef]
- Aldoss, I.; Douer, D.; Behrendt, C.E.; Chaudhary, P.; Mohrbacher, A.; Vrona, J.; Pullarkat, V. Toxicity profile of repeated doses of PEG-asparaginase incorporated into a pediatric-type regimen for adult acute lymphoblastic leukemia. Eur. J. Haematol. 2016, 96, 375–380. [Google Scholar] [CrossRef]
- Tong, W.H.; Pieters, R.; de Groot-Kruseman, H.A.; Hop, W.C.; Boos, J.; Tissing, W.J.; van der Sluis, I.M. The toxicity of very prolonged courses of PEGasparaginase or Erwinia asparaginase in relation to asparaginase activity, with a special focus on dyslipidemia. Haematologica 2014, 99, 1716–1721. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, R.S.; Panosyan, E.H.; Seibel, N.L.; Gaynon, P.S.; Avramis, I.A.; Avramis, V.I. Changes of amino acid serum levels in pediatric patients with higher-risk acute lymphoblastic leukemia (CCG-1961). In Vivo 2004, 18, 107–112. [Google Scholar] [PubMed]
- Avramis, V.I.; Sencer, S.; Periclou, A.P.; Sather, H.; Bostrom, B.C.; Cohen, L.J.; Ettinger, A.G.; Ettinger, L.J.; Franklin, J.; Gaynon, P.S.; et al. A randomized comparison of native Escherichia coli asparaginase and polyethylene glycol conjugated asparaginase for treatment of children with newly diagnosed standard-risk acute lymphoblastic leukemia: A Children’s Cancer Group study. Blood 2002, 99, 1986–1994. [Google Scholar] [CrossRef] [Green Version]
- Heitink-Pollé, K.M.; Prinsen, B.H.; de Koning, T.J.; van Hasselt, P.M.; Bierings, M.B. High incidence of symptomatic hyperammonemia in children with acute lymphoblastic leukemia receiving pegylated asparaginase. JIMD Rep. 2013, 7, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollenschläger, G.; Roth, E.; Linkesch, W.; Jansen, S.; Simmel, A.; Mödder, B. Asparaginase-induced derangements of glutamine metabolism: The pathogenetic basis for some drug-related side-effects. Eur. J. Clin. Investig. 1988, 18, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Villa, P.; Corada, M.; Bartosek, I. L-asparaginase effects on inhibition of protein synthesis and lowering of the glutamine content in cultured rat hepatocytes. Toxicol. Lett. 1986, 32, 235–241. [Google Scholar] [CrossRef]
- Laterza, O.F.; Gerhardt, G.; Sokoll, L.J. Measurement of plasma ammonia is affected in patients receiving asparaginase therapy. Clin. Chem. 2003, 49, 1710–1711. [Google Scholar] [CrossRef] [Green Version]
- Jörck, C.; Kiess, W.; Weigel, J.F.; Mütze, U.; Bierbach, U.; Beblo, S. Transient hyperammonemia due to L-asparaginase therapy in children with acute lymphoblastic leukemia or non-Hodgkin lymphoma. Pediatr. Hematol. Oncol. 2011, 28, 3–9. [Google Scholar] [CrossRef]
- Nussbaum, V.; Lubcke, N.; Findlay, R. Hyperammonemia secondary to asparaginase: A case series. J. Oncol. Pharm. Pract. 2016, 22, 161–164. [Google Scholar] [CrossRef]
- Kessel, D. Asparaginyl-transfer RNA: A substrate for L-asparaginase. Biochim. Biophys. Acta (BBA)—Nucleic Acids Protein Synth. 1971, 240, 554–557. [Google Scholar] [CrossRef]
- Fidler, I.J.; Montgomery, P.C. Effects of L-asparaginase on lymphocyte surface and blastogenesis. Cancer Res. 1972, 32, 2400–2406. [Google Scholar] [PubMed]
- Ankel, E.G.; Zirneski, J.; Ring, B.J.; Holcenberg, J.S. Effect of asparaginase on cell membranes of sensitive and resistants mouse lymphoma cells. In Vitro 1984, 20, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, D.D.; Pokrovsky, V.S.; Pokrovskaya, M.V.; Alexandrova, S.S.; Eldarov, M.A.; Grishin, D.V.; Basharov, M.M.; Gladilina, Y.A.; Podobed, O.V.; Sokolov, N.N. Rhodospirillum rubrum L-asparaginase targets tumor growth by a dual mechanism involving telomerase inhibition. Biochem. Biophys. Res. Commun. 2017, 492, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Plyasova, A.A.; Pokrovskaya, M.V.; Lisitsyna, O.M.; Pokrovsky, V.S.; Alexandrova, S.S.; Hilal, A.; Sokolov, N.N.; Zhdanov, D.D. Penetration into cancer cells via clathrin-dependent mechanism allows l-asparaginase from rhodospirillum rubrum to inhibit telomerase. Pharmaceuticals 2020, 13, 286. [Google Scholar] [CrossRef]
- Balasubramanian, M.N.; Butterworth, E.A.; Kilberg, M.S. Asparagine synthetase: Regulation by cell stress and involvement in tumor biology. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E789–E799. [Google Scholar] [CrossRef] [Green Version]
- Kilberg, M.S.; Balasubramanian, M.; Fu, L.; Shan, J. The transcription factor network associated with the amino acid response in mammalian cells. Adv. Nutr. 2012, 3, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino acids in cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, X.; Cheng, C.; Yu, W.; Yi, P. Metabolism of Amino Acids in Cancer. Front. Cell Dev. Biol. 2020, 8, 603837. [Google Scholar] [CrossRef]
- Wek, S.A.; Zhu, S.; Wek, R.C. The histidyl-tRNA synthetase-related sequence in the eIF-2 alpha protein kinase GCN2 interacts with tRNA and is required for activation in response to starvation for different amino acids. Mol. Cell Biol. 1995, 15, 4497–4506. [Google Scholar] [CrossRef] [Green Version]
- Wek, R.C.; Ramirez, M.; Jackson, B.M.; Hinnebusch, A.G. Identification of positive-acting domains in GCN2 protein kinase required for translational activation of GCN4 expression. Mol. Cell Biol. 1990, 10, 2820–2831. [Google Scholar] [CrossRef]
- Shan, J.; Fu, L.; Balasubramanian, M.N.; Anthony, T.; Kilberg, M.S. ATF4-dependent regulation of the JMJD3 gene during amino acid deprivation can be rescued in Atf4-deficient cells by inhibition of deacetylation. J. Biol. Chem. 2012, 287, 36393–36403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siu, F.; Bain, P.J.; LeBlanc-Chaffin, R.; Chen, H.; Kilberg, M.S. ATF4 is a mediator of the nutrient-sensing response pathway that activates the human asparagine synthetase gene. J. Biol. Chem. 2002, 277, 24120–24127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, N.; Thiaville, M.M.; Awad, K.; Gjymishka, A.; Brant, J.O.; Yang, T.P.; Kilberg, M.S. Protein or amino acid deprivation differentially regulates the hepatic forkhead box protein A (FOXA) genes through an activating transcription factor-4-independent pathway. Hepatology 2009, 50, 282–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilberg, M.S.; Shan, J.; Su, N. ATF4-dependent transcription mediates signaling of amino acid limitation. Trends Endocrinol. Metab. 2009, 20, 436–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, T.D.; Wek, R.C. Eukaryotic initiation factor 2 phosphorylation and translational control in metabolism. Adv. Nutr. 2012, 3, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, H.P.; Novoa, I.; Zhang, Y.; Zeng, H.; Wek, R.; Schapira, M.; Ron, D. Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol. Cell 2000, 6, 1099–1108. [Google Scholar] [CrossRef]
- Barbosa-Tessmann, I.P.; Chen, C.; Zhong, C.; Siu, F.; Schuster, S.M.; Nick, H.S.; Kilberg, M.S. Activation of the human asparagine synthetase gene by the amino acid response and the endoplasmic reticulum stress response pathways occurs by common genomic elements. J. Biol. Chem. 2000, 275, 26976–26985. [Google Scholar] [CrossRef]
- Ren, Y.; Roy, S.; Ding, Y.; Iqbal, J.; Broome, J.D. Methylation of the asparagine synthetase promoter in human leukemic cell lines is associated with a specific methyl binding protein. Oncogene 2004, 23, 3953–3961. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Srivastava, S.; Seim, G.; Pavlova, N.N.; King, B.; Zou, L.; Zhang, C.; Zhong, M.; Feng, H.; Kapur, R.; et al. Promoter demethylation of the asparagine synthetase gene is required for ATF4-dependent adaptation to asparagine depletion. J. Biol. Chem. 2019, 294, 18674–18684. [Google Scholar] [CrossRef]
- Touzart, A.; Lengliné, E.; Latiri, M.; Belhocine, M.; Smith, C.; Thomas, X.; Spicuglia, S.; Puthier, D.; Pflumio, F.; Leguay, T.; et al. Epigenetic Silencing Affects l-Asparaginase Sensitivity and Predicts Outcome in T-ALL. Clin. Cancer Res. 2019, 25, 2483–2493. [Google Scholar] [CrossRef] [Green Version]
- Horowitz, B.; Madras, B.K.; Meister, A.; Old, L.J.; Boyes, E.A.; Stockert, E. Asparagine synthetase activity of mouse leukemias. Science 1968, 160, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Pan, Y.X.; Zhou, M.; Harvey, R.C.; Hunger, S.P.; Kilberg, M.S. Correlation between asparaginase sensitivity and asparagine synthetase protein content, but not mRNA, in acute lymphoblastic leukemia cell lines. Pediatr. Blood Cancer 2008, 50, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Yin, D.; Kawamata, N.; Bartram, C.R.; Hofmann, W.K.; Wolf, I.; Miller, C.W.; Koeffler, H.P. Methylation analysis of asparagine synthetase gene in acute lymphoblastic leukemia cells. Leukemia 2006, 20, 1303–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, A.; Miyake, K.; Nordlund, J.; Syvänen, A.C.; van der Weyden, L.; Honda, H.; Yamasaki, N.; Nagamachi, A.; Inaba, T.; Ikawa, T.; et al. Association of aberrant ASNS imprinting with asparaginase sensitivity and chromosomal abnormality in childhood BCP-ALL. Blood 2020, 136, 2319–2333. [Google Scholar] [CrossRef]
- Peng, H.; Shen, N.; Qian, L.; Sun, X.L.; Koduru, P.; Goodwin, L.O.; Issa, J.P.; Broome, J.D. Hypermethylation of CpG islands in the mouse asparagine synthetase gene: Relationship to asparaginase sensitivity in lymphoma cells. Partial methylation in normal cells. Br. J. Cancer 2001, 85, 930–935. [Google Scholar] [CrossRef] [Green Version]
- Willems, L.; Jacque, N.; Jacquel, A.; Neveux, N.; Maciel, T.T.; Lambert, M.; Schmitt, A.; Poulain, L.; Green, A.S.; Uzunov, M.; et al. Inhibiting glutamine uptake represents an attractive new strategy for treating acute myeloid leukemia. Blood 2013, 122, 3521–3532. [Google Scholar] [CrossRef] [Green Version]
- Buaboonnam, J.; Cao, X.; Pauley, J.L.; Pui, C.H.; Ribeiro, R.C.; Rubnitz, J.E.; Inaba, H. Sequential administration of methotrexate and asparaginase in relapsed or refractory pediatric acute myeloid leukemia. Pediatr. Blood Cancer 2013, 60, 1161–1164. [Google Scholar] [CrossRef] [Green Version]
- Emadi, A.; Law, J.Y.; Strovel, E.T.; Lapidus, R.G.; Jeng, L.J.B.; Lee, M.; Blitzer, M.G.; Carter-Cooper, B.A.; Sewell, D.; Van Der Merwe, I.; et al. Asparaginase Erwinia chrysanthemi effectively depletes plasma glutamine in adult patients with relapsed/refractory acute myeloid leukemia. Cancer Chemother. Pharm. 2018, 81, 217–222. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, J.; Zeng, H.; Zhang, Y.; Zhang, Y.; Zhou, X.; Zhou, H. Antiproliferative effects of L-asparaginase in acute myeloid leukemia. Exp. Med. 2020, 20, 2070–2078. [Google Scholar] [CrossRef]
- Purwaha, P.; Lorenzi, P.L.; Silva, L.P.; Hawke, D.H.; Weinstein, J.N. Targeted metabolomic analysis of amino acid response to L-asparaginase in adherent cells. Metabolomics 2014, 10, 909–919. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-K.; Park, K.-G. Targeting Glutamine Metabolism for Cancer Treatment. Biomol. Ther. 2018, 26, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toda, K.; Kawada, K.; Iwamoto, M.; Inamoto, S.; Sasazuki, T.; Shirasawa, S.; Hasegawa, S.; Sakai, Y. Metabolic Alterations Caused by KRAS Mutations in Colorectal Cancer Contribute to Cell Adaptation to Glutamine Depletion by Upregulation of Asparagine Synthetase. Neoplasia 2016, 18, 654–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinze, L.; Labrosse, R.; Degar, J.; Han, T.; Schatoff, E.M.; Schreek, S.; Karim, S.; McGuckin, C.; Sacher, J.R.; Wagner, F.; et al. Exploiting the Therapeutic Interaction of WNT Pathway Activation and Asparaginase for Colorectal Cancer Therapy. Cancer Discov. 2020, 10, 1690–1705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Dong, L.W.; Tan, Y.X.; Zhang, J.; Pan, Y.F.; Yang, C.; Li, M.H.; Ding, Z.W.; Liu, L.J.; Jiang, T.Y.; et al. Asparagine synthetase is an independent predictor of surgical survival and a potential therapeutic target in hepatocellular carcinoma. Br. J. Cancer 2013, 109, 14–23. [Google Scholar] [CrossRef]
- Li, W.; Dong, C. Polymorphism in asparagine synthetase is associated with overall survival of hepatocellular carcinoma patients. BMC Gastroenterol. 2017, 17, 79. [Google Scholar] [CrossRef] [Green Version]
- Panosyan, E.H.; Wang, Y.; Xia, P.; Lee, W.N.; Pak, Y.; Laks, D.R.; Lin, H.J.; Moore, T.B.; Cloughesy, T.F.; Kornblum, H.I.; et al. Asparagine depletion potentiates the cytotoxic effect of chemotherapy against brain tumors. Mol. Cancer Res. 2014, 12, 694–702. [Google Scholar] [CrossRef] [Green Version]
- Ohba, S.; Hirose, Y. L-asparaginase and 6-diazo-5-oxo-L-norleucine synergistically inhibit the growth of glioblastoma cells. J. Neurooncol. 2020, 146, 469–475. [Google Scholar] [CrossRef]
- Dübbers, A.; Würthwein, G.; Müller, H.J.; Schulze-Westhoff, P.; Winkelhorst, M.; Kurzknabe, E.; Lanvers, C.; Pieters, R.; Kaspers, G.J.; Creutzig, U.; et al. Asparagine synthetase activity in paediatric acute leukaemias: AML-M5 subtype shows lowest activity. Br. J. Haematol. 2000, 109, 427–429. [Google Scholar] [CrossRef]
- Okada, S.; Hongo, T.; Yamada, S.; Watanabe, C.; Fujii, Y.; Ohzeki, T.; Horikoshi, Y.; Ito, T.; Yazaki, M.; Komada, Y.; et al. In vitro efficacy of l-asparaginase in childhood acute myeloid leukaemia. Br. J. Haematol. 2003, 123, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Michelozzi, I.M.; Granata, V.; De Ponti, G.; Alberti, G.; Tomasoni, C.; Antolini, L.; Gambacorti-Passerini, C.; Gentner, B.; Dazzi, F.; Biondi, A.; et al. Acute myeloid leukaemia niche regulates response to L-asparaginase. Br. J. Haematol. 2019, 186, 420–430. [Google Scholar] [CrossRef]
- Kaspers, G.J.L. Acute myeloid leukaemia niche regulates response to L-asparaginase. Br. J. Haematol. 2019, 186, 397–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherf, U.; Ross, D.T.; Waltham, M.; Smith, L.H.; Lee, J.K.; Tanabe, L.; Kohn, K.W.; Reinhold, W.C.; Myers, T.G.; Andrews, D.T.; et al. A gene expression database for the molecular pharmacology of cancer. Nat. Genet. 2000, 24, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.M.; Der, C.J. KRAS: The Critical Driver and Therapeutic Target for Pancreatic Cancer. Cold Spring Harb. Perspect. Med. 2018, 8, a031435. [Google Scholar] [CrossRef] [PubMed]
- Gaglio, D.; Metallo, C.M.; Gameiro, P.A.; Hiller, K.; Danna, L.S.; Balestrieri, C.; Alberghina, L.; Stephanopoulos, G.; Chiaradonna, F. Oncogenic K-Ras decouples glucose and glutamine metabolism to support cancer cell growth. Mol. Syst. Biol. 2011, 7, 523. [Google Scholar] [CrossRef] [PubMed]
- Son, J.; Lyssiotis, C.A.; Ying, H.; Wang, X.; Hua, S.; Ligorio, M.; Perera, R.M.; Ferrone, C.R.; Mullarky, E.; Shyh-Chang, N.; et al. Glutamine supports pancreatic cancer growth through a KRAS-regulated metabolic pathway. Nature 2013, 496, 101–105. [Google Scholar] [CrossRef]
- Li, D.; Fu, Z.; Chen, R.; Zhao, X.; Zhou, Y.; Zeng, B.; Yu, M.; Zhou, Q.; Lin, Q.; Gao, W.; et al. Inhibition of glutamine metabolism counteracts pancreatic cancer stem cell features and sensitizes cells to radiotherapy. Oncotarget 2015, 6, 31151–31163. [Google Scholar] [CrossRef] [Green Version]
- Porru, M.; Pompili, L.; Caruso, C.; Biroccio, A.; Leonetti, C. Targeting KRAS in metastatic colorectal cancer: Current strategies and emerging opportunities. J. Exp. Clin. Cancer Res. 2018, 37, 57. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Chen, J.; Liu, H.; Cai, Y.; Cao, T.; Han, W.; Yi, X.; Qian, M.; Tian, D.; Nie, Y.; et al. SOX12 promotes colorectal cancer cell proliferation and metastasis by regulating asparagine synthesis. Cell Death Dis. 2019, 10, 239. [Google Scholar] [CrossRef]
- Cao, J.; Wang, X.; Dai, T.; Wu, Y.; Zhang, M.; Cao, R.; Zhang, R.; Wang, G.; Jiang, R.; Zhou, B.P.; et al. Twist promotes tumor metastasis in basal-like breast cancer by transcriptionally upregulating ROR1. Theranostics 2018, 8, 2739–2751. [Google Scholar] [CrossRef]
- Luo, M.; Shang, L.; Brooks, M.D.; Jiagge, E.; Zhu, Y.; Buschhaus, J.M.; Conley, S.; Fath, M.A.; Davis, A.; Gheordunescu, E.; et al. Targeting Breast Cancer Stem Cell State Equilibrium through Modulation of Redox Signaling. Cell Metab. 2018, 28, 69–86. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhang, M.; Xu, F.; Jiang, S. Wnt signaling in breast cancer: Biological mechanisms, challenges and opportunities. Mol. Cancer 2020, 19, 165. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Hui, S.; Ghergurovich, J.M.; Fan, J.; Intlekofer, A.M.; White, R.M.; Rabinowitz, J.D.; Thompson, C.B.; Zhang, J. As Extracellular Glutamine Levels Decline, Asparagine Becomes an Essential Amino Acid. Cell Metab. 2018, 27, 428–438. [Google Scholar] [CrossRef] [Green Version]
- Wise, D.R.; Thompson, C.B. Glutamine addiction: A new therapeutic target in cancer. Trends Biochem. Sci. 2010, 35, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Chiu, M.; Tardito, S.; Pillozzi, S.; Arcangeli, A.; Armento, A.; Uggeri, J.; Missale, G.; Bianchi, M.G.; Barilli, A.; Dall’Asta, V.; et al. Glutamine depletion by crisantaspase hinders the growth of human hepatocellular carcinoma xenografts. Br. J. Cancer 2014, 111, 1159–1167. [Google Scholar] [CrossRef]
- Gross, M.I.; Demo, S.D.; Dennison, J.B.; Chen, L.; Chernov-Rogan, T.; Goyal, B.; Janes, J.R.; Laidig, G.J.; Lewis, E.R.; Li, J.; et al. Antitumor activity of the glutaminase inhibitor CB-839 in triple-negative breast cancer. Mol. Cancer 2014, 13, 890–901. [Google Scholar] [CrossRef] [Green Version]
- Fung, M.K.L.; Chan, G.C.-F. Drug-induced amino acid deprivation as strategy for cancer therapy. J. Hematol. Oncol. 2017, 10, 144. [Google Scholar] [CrossRef]
- Hortobagyi, G.N.; Yap, H.Y.; Wiseman, C.L.; Blumenschein, G.R.; Buzdar, A.U.; Legha, S.S.; Gutterman, J.U.; Hersh, E.M.; Bodey, G.P. Chemoimmunotherapy for metastatic breast cancer with 5-fluorouracil, adriamycin, cyclophosphamide, methotrexate, L-asparaginase, Corynebacterium parvum, and Pseudomonas vaccine. Cancer Treat. Rep. 1980, 64, 157–159. [Google Scholar]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef]
- Bhowmik, A.; Khan, R.; Ghosh, M.K. Blood brain barrier: A challenge for effectual therapy of brain tumors. Biomed. Res. Int. 2015, 2015, 320941. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, D.S.; Park, J.R.; Thomson, B.G.; Felgenhauer, J.L.; Holcenberg, J.S.; Panosyan, E.H.; Avramis, V.I. Asparaginase pharmacokinetics after intensive polyethylene glycol-conjugated L-asparaginase therapy for children with relapsed acute lymphoblastic leukemia. Clin. Cancer Res. 2004, 10, 5335–5341. [Google Scholar] [CrossRef] [Green Version]
- Panetta, J.C.; Gajjar, A.; Hijiya, N.; Hak, L.J.; Cheng, C.; Liu, W.; Pui, C.H.; Relling, M.V. Comparison of native E. coli and PEG asparaginase pharmacokinetics and pharmacodynamics in pediatric acute lymphoblastic leukemia. Clin. Pharm. 2009, 86, 651–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira Pinheiro, J.P.; Wenner, K.; Escherich, G.; Lanvers-Kaminsky, C.; Würthwein, G.; Janka-Schaub, G.; Boos, J. Serum asparaginase activities and asparagine concentrations in the cerebrospinal fluid after a single infusion of 2,500 IU/m(2) PEG asparaginase in children with ALL treated according to protocol COALL-06-97. Pediatr. Blood Cancer 2006, 46, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Lanvers-Kaminsky, C. Asparaginase pharmacology: Challenges still to be faced. Cancer Chemother. Pharm. 2017, 79, 439–450. [Google Scholar] [CrossRef]
- Rizzari, C.; Zucchetti, M.; Conter, V.; Diomede, L.; Bruno, A.; Gavazzi, L.; Paganini, M.; Sparano, P.; Lo Nigro, L.; Aricò, M.; et al. L-asparagine depletion and L-asparaginase activity in children with acute lymphoblastic leukemia receiving i.m. or i.v. Erwinia C. or E. coli L-asparaginase as first exposure. Ann. Oncol. 2000, 11, 189–193. [Google Scholar] [CrossRef]
- Rizzari, C.; Citterio, M.; Zucchetti, M.; Conter, V.; Chiesa, R.; Colombini, A.; Malguzzi, S.; Silvestri, D.; D’Incalci, M. A pharmacological study on pegylated asparaginase used in front-line treatment of children with acute lymphoblastic leukemia. Haematologica 2006, 91, 24–31. [Google Scholar] [PubMed]
- Asselin, B.; Rizzari, C. Asparaginase pharmacokinetics and implications of therapeutic drug monitoring. Leuk. Lymphoma 2015, 56, 2273–2280. [Google Scholar] [CrossRef]
- Henriksen, L.T.; Nersting, J.; Raja, R.A.; Frandsen, T.L.; Rosthøj, S.; Schrøder, H.; Albertsen, B.K. Cerebrospinal fluid asparagine depletion during pegylated asparaginase therapy in children with acute lymphoblastic leukaemia. Br. J. Haematol. 2014, 166, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Riccardi, R.; Holcenberg, J.S.; Glaubiger, D.L.; Wood, J.H.; Poplack, D.G. L-asparaginase pharmacokinetics and asparagine levels in cerebrospinal fluid of rhesus monkeys and humans. Cancer Res. 1981, 41, 4554–4558. [Google Scholar]
- Ballerini, A.; Moro, F.; Nerini, I.F.; Marzo, C.M.; Di Clemente, A.; Ferrari, M.; D’Incalci, M.; Biondi, A.; Colombini, A.; Conter, V.; et al. Pharmacodynamic effects in the cerebrospinal fluid of rats after intravenous administration of different asparaginase formulations. Cancer Chemother. Pharm. 2017, 79, 1267–1271. [Google Scholar] [CrossRef]
- Koleva, L.; Bovt, E.; Ataullakhanov, F.; Sinauridze, E. Erythrocytes as Carriers: From Drug Delivery to Biosensors. Pharmaceutics 2020, 12, 276. [Google Scholar] [CrossRef] [Green Version]
- Rossi, L.; Pierigè, F.; Aliano, M.P.; Magnani, M. Ongoing Developments and Clinical Progress in Drug-Loaded Red Blood Cell Technologies. BioDrugs 2020, 34, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Domenech, C.; Thomas, X.; Chabaud, S.; Baruchel, A.; Gueyffier, F.; Mazingue, F.; Auvrignon, A.; Corm, S.; Dombret, H.; Chevallier, P.; et al. l-asparaginase loaded red blood cells in refractory or relapsing acute lymphoblastic leukaemia in children and adults: Results of the GRASPALL 2005-01 randomized trial. Br. J. Haematol. 2011, 153, 58–65. [Google Scholar] [CrossRef]
- Hunault-Berger, M.; Leguay, T.; Huguet, F.; Leprêtre, S.; Deconinck, E.; Ojeda-Uribe, M.; Bonmati, C.; Escoffre-Barbe, M.; Bories, P.; Himberlin, C.; et al. A Phase 2 study of L-asparaginase encapsulated in erythrocytes in elderly patients with Philadelphia chromosome negative acute lymphoblastic leukemia: The GRASPALL/GRAALL-SA2-2008 study. Am. J. Hematol. 2015, 90, 811–818. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. Graspa: Withdrawal of the Marketing Authorisation Application. Available online: https://www.ema.europa.eu/en/medicines/human/withdrawn-applications/graspa-0 (accessed on 16 December 2021).
- Bachet, J.B.; Gay, F.; Maréchal, R.; Galais, M.P.; Adenis, A.; Ms, C.D.; Cros, J.; Demetter, P.; Svrcek, M.; Bardier-Dupas, A.; et al. Asparagine Synthetase Expression and Phase I Study With L-Asparaginase Encapsulated in Red Blood Cells in Patients With Pancreatic Adenocarcinoma. Pancreas 2015, 44, 1141–1147. [Google Scholar] [CrossRef]
- Hammel, P.; Bachet, J.B.; Portales, F.; Mineur, L.; Metges, J.P.; de la Fouchardiere, C.; Louvet, C.; El Hajbi, F.; Faroux, R.; Guimbaud, R.; et al. 621PD—A Phase 2b of eryaspase in combination with gemcitabine or FOLFOX as second-line therapy in patients with metastatic pancreatic adenocarcinoma (NCT02195180). Ann. Oncol. 2017, 28, v211. [Google Scholar] [CrossRef]
- Hammel, P.; Fabienne, P.; Mineur, L.; Metges, J.P.; Andre, T.; De La Fouchardiere, C.; Louvet, C.; El Hajbi, F.; Faroux, R.; Guimbaud, R.; et al. Erythrocyte-encapsulated asparaginase (eryaspase) combined with chemotherapy in second-line treatment of advanced pancreatic cancer: An open-label, randomized Phase IIb trial. Eur. J. Cancer 2020, 124, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Erytech. ERYTECH Granted, U.S. FDA Fast Track Designation for Eryaspase in Hypersensitive ALL. Available online: https://investors.erytech.com/news-releases/news-release-details/erytech-granted-us-fda-fast-track-designation-eryaspase (accessed on 20 December 2021).
- Distasio, J.A.; Durden, D.L.; Paul, R.D.; Nadji, M. Alteration in spleen lymphoid populations associated with specific amino acid depletion during L-asparaginase treatment. Cancer Res. 1982, 42, 252–258. [Google Scholar]
- Distasio, J.A.; Niederman, R.A.; Kafkewitz, D. Antilymphoma activity of a glutaminase-free L-asparaginase of microbial origin. Proc. Soc. Exp. Biol. Med. 1977, 155, 528–531. [Google Scholar] [CrossRef]
- Durden, D.L.; Distasio, J.A. Comparison of the immunosuppressive effects of asparaginases from Escherichia coli and Vibrio succinogenes. Cancer Res. 1980, 40, 1125–1129. [Google Scholar]
- Distasio, J.A.; Niederman, R.A.; Kafkewitz, D.; Goodman, D. Purification and characterization of L-asparaginase with anti-lymphoma activity from Vibrio succinogenes. J. Biol. Chem. 1976, 251, 6929–6933. [Google Scholar] [CrossRef]
- Bendich, A.; Kafkewitz, D.; Abuchowski, A.; Davis, F.F. Immunological effects of native and polyethylene glycol-modified asparaginases from Vibrio succinogenes and Escherichia coli in normal and tumour-bearing mice. Clin. Exp. Immunol. 1982, 48, 273–278. [Google Scholar]
- Nguyen, H.A.; Durden, D.L.; Lavie, A. The differential ability of asparagine and glutamine in promoting the closed/active enzyme conformation rationalizes the Wolinella succinogenes L-asparaginase substrate specificity. Sci. Rep. 2017, 7, 41643. [Google Scholar] [CrossRef] [Green Version]
- Baar, C.; Eppinger, M.; Raddatz, G.; Simon, J.; Lanz, C.; Klimmek, O.; Nandakumar, R.; Gross, R.; Rosinus, A.; Keller, H.; et al. Complete genome sequence and analysis of Wolinella succinogenes. Proc. Natl. Acad. Sci. USA 2003, 100, 11690–11695. [Google Scholar] [CrossRef] [Green Version]
- Lubkowski, J.; Palm, G.J.; Gilliland, G.L.; Derst, C.; Röhm, K.H.; Wlodawer, A. Crystal structure and amino acid sequence of Wolinella succinogenes L-asparaginase. Eur. J. Biochem. 1996, 241, 201–207. [Google Scholar] [CrossRef]
- Reinert, R.B.; Oberle, L.M.; Wek, S.A.; Bunpo, P.; Wang, X.P.; Mileva, I.; Goodwin, L.O.; Aldrich, C.J.; Durden, D.L.; McNurlan, M.A.; et al. Role of glutamine depletion in directing tissue-specific nutrient stress responses to L-asparaginase. J. Biol. Chem. 2006, 281, 31222–31233. [Google Scholar] [CrossRef]
- Emadi, A.; Kapadia, B.; Bollino, D.; Bhandary, B.; Baer, M.R.; Niyongere, S.; Strovel, E.T.; Kaizer, H.; Chang, E.; Choi, E.Y.; et al. Venetoclax and pegcrisantaspase for complex karyotype acute myeloid leukemia. Leukemia 2021, 35, 1907–1924. [Google Scholar] [CrossRef]
- Nikonorova, I.A.; Zhu, Q.; Signore, C.C.; Mirek, E.T.; Jonsson, W.O.; Kong, B.; Guo, G.L.; Belden, W.J.; Anthony, T.G. Age modulates liver responses to asparaginase-induced amino acid stress in mice. J. Biol. Chem. 2019, 294, 13864–13875. [Google Scholar] [CrossRef] [Green Version]
- Schalk, A.M.; Lavie, A. Structural and kinetic characterization of guinea pig L-asparaginase type III. Biochemistry 2014, 53, 2318–2328. [Google Scholar] [CrossRef]
- Schalk, A.M.; Nguyen, H.A.; Rigouin, C.; Lavie, A. Identification and structural analysis of an L-asparaginase enzyme from guinea pig with putative tumor cell killing properties. J. Biol. Chem. 2014, 289, 33175–33186. [Google Scholar] [CrossRef] [Green Version]
- Rigouin, C.; Nguyen, H.A.; Schalk, A.M.; Lavie, A. Discovery of human-like L-asparaginases with potential clinical use by directed evolution. Sci. Rep. 2017, 7, 10224. [Google Scholar] [CrossRef] [Green Version]
- Tower, D.B.; Peters, E.L.; Curtis, W.C. Guinea pig serum L-asparaginase. Properties, purification, and application to determination of asparagine in biological samples. J. Biol. Chem. 1963, 238, 983–993. [Google Scholar] [CrossRef]
- Cantor, J.R.; Stone, E.M.; Chantranupong, L.; Georgiou, G. The human asparaginase-like protein 1 hASRGL1 is an Ntn hydrolase with beta-aspartyl peptidase activity. Biochemistry 2009, 48, 11026–11031. [Google Scholar] [CrossRef] [Green Version]
- Karamitros, C.S.; Konrad, M. Human 60-kDa lysophospholipase contains an N-terminal L-asparaginase domain that is allosterically regulated by L-asparagine. J. Biol. Chem. 2014, 289, 12962–12975. [Google Scholar] [CrossRef] [Green Version]
- Maggi, M.; Mittelman, S.D.; Parmentier, J.H.; Colombo, G.; Meli, M.; Whitmire, J.M.; Merrell, D.S.; Whitelegge, J.; Scotti, C. A protease-resistant Escherichia coli asparaginase with outstanding stability and enhanced anti-leukaemic activity in vitro. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.A.; Su, Y.; Lavie, A. Design and characterization of erwinia chrysanthemi l-asparaginase variants with diminished l-glutaminase activity. J. Biol. Chem. 2016, 291, 17664–17676. [Google Scholar] [CrossRef] [Green Version]
- Offman, M.N.; Krol, M.; Patel, N.; Krishnan, S.; Liu, J.; Saha, V.; Bates, P.A. Rational engineering of L-asparaginase reveals importance of dual activity for cancer cell toxicity. Blood J. Am. Soc. Hematol. 2011, 117, 1614–1621. [Google Scholar] [CrossRef] [Green Version]
- Jianhua, C.; Yujun, W.; Ruibo, J.; Min, W.; Wutong, W. Probing the antigenicity of E. coli L-asparaginase by mutational analysis. Mol. Biotechnol. 2006, 33, 57–65. [Google Scholar]
- Ghasemi, A.; Asad, S.; Kabiri, M.; Dabirmanesh, B. Cloning and characterization of Halomonas elongata L-asparaginase, a promising chemotherapeutic agent. Appl. Microbiol. Biotechnol. 2017, 101, 7227–7238. [Google Scholar] [CrossRef]
- Bansal, S.; Srivastava, A.; Mukherjee, G.; Pandey, R.; Verma, A.K.; Mishra, P.; Kundu, B. Hyperthermophilic asparaginase mutants with enhanced substrate affinity and antineoplastic activity: Structural insights on their mechanism of action. FASEB J. 2012, 26, 1161–1171. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Kumanova, M.; Hart, L.S.; Sloane, K.; Zhang, H.; De Panis, D.N.; Bobrovnikova-Marjon, E.; Diehl, J.A.; Ron, D.; Koumenis, C. The GCN2-ATF4 pathway is critical for tumour cell survival and proliferation in response to nutrient deprivation. EMBO J. 2010, 29, 2082–2096. [Google Scholar] [CrossRef] [Green Version]
- Bunpo, P.; Dudley, A.; Cundiff, J.K.; Cavener, D.R.; Wek, R.C.; Anthony, T.G. GCN2 protein kinase is required to activate amino acid deprivation responses in mice treated with the anti-cancer agent L-asparaginase. J. Biol. Chem. 2009, 284, 32742–32749. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Nambu, T.; Ebara, S.; Hasegawa, Y.; Toyoshima, K.; Tsuchiya, Y.; Tomita, D.; Fujimoto, J.; Kurasawa, O.; Takahara, C.; et al. Inhibition of GCN2 sensitizes ASNS-low cancer cells to asparaginase by disrupting the amino acid response. Proc. Natl. Acad. Sci. USA 2018, 115, E7776–E7785. [Google Scholar] [CrossRef] [Green Version]
- Apfel, V.; Begue, D.; Cordo, V.; Holzer, L.; Martinuzzi, L.; Buhles, A.; Kerr, G.; Barbosa, I.; Naumann, U.; Piquet, M.; et al. Therapeutic Assessment of Targeting ASNS Combined with l-Asparaginase Treatment in Solid Tumors and Investigation of Resistance Mechanisms. ACS Pharm. Transl. Sci. 2021, 4, 327–337. [Google Scholar] [CrossRef]
- Fujimoto, J.; Kurasawa, O.; Takagi, T.; Liu, X.; Banno, H.; Kojima, T.; Asano, Y.; Nakamura, A.; Nambu, T.; Hata, A.; et al. Identification of Novel, Potent, and Orally Available GCN2 Inhibitors with Type I Half Binding Mode. ACS Med. Chem. Lett. 2019, 10, 1498–1503. [Google Scholar] [CrossRef]
- Butler, M.; van Ingen Schenau, D.S.; Yu, J.; Jenni, S.; Dobay, M.P.; Hagelaar, R.; Vervoort, B.M.T.; Tee, T.M.; Hoff, F.W.; Meijerink, J.P.; et al. BTK inhibition sensitizes Acute Lymphoblastic Leukemia to asparaginase by suppressing the Amino Acid Response pathway. Blood 2021, 138, 2383–2395. [Google Scholar] [CrossRef]
- Sun, J.; Nagel, R.; Zaal, E.A.; Ugalde, A.P.; Han, R.; Proost, N.; Song, J.Y.; Pataskar, A.; Burylo, A.; Fu, H.; et al. SLC1A3 contributes to L-asparaginase resistance in solid tumors. EMBO J. 2019, 38, e102147. [Google Scholar] [CrossRef]
- Hinze, L.; Pfirrmann, M.; Karim, S.; Degar, J.; McGuckin, C.; Vinjamur, D.; Sacher, J.; Stevenson, K.E.; Neuberg, D.S.; Orellana, E.; et al. Synthetic Lethality of Wnt Pathway Activation and Asparaginase in Drug-Resistant Acute Leukemias. Cancer Cell 2019, 35, 664–676. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Name | Origin | Additions/Tags | Remarks | References |
|---|---|---|---|---|
| Elspar® | E. coli Type 2 | Native | [35] | |
| Kidrolase® | E. coli Type 2 | Native | [36] | |
| Spectrila® | E. coli Type 2 | Native | [37] | |
| Oncaspar® | E. coli Type 2 | PEG (Succinimidyl succinate linker) | [25] | |
| Asparlas® | E. coli Type 2 | PEG (Succinimidyl carbonate linker) | Only FDA approved | [27] [28] |
| Erwinase® | E. chrysanthemi ansB gene | Native | [23] [24] | |
| Rylaze® | E. chrysanthemi ansB gene | Native | Only FDA approved | [23] [24] |
| Cancer Type | L-ASNase Sensitivity Mechanism | Combination Therapy | Findings | References |
|---|---|---|---|---|
| Acute myeloid leukemia | ASNS-low | Methotrexate | Gln bio-availability is important for growth. Importance glutaminase co-activity is unclear | [106] [107] [108] [109] |
| Cytarabine | ||||
| Ovarian clear cell carcinoma | ASNS-low | Glutaminase co-activity might be important for cytotoxic effect | [18] [110] | |
| Pancreatic ductal adeno-carcinoma (PDAC) | ASNS-low | 52% of PDAC classified as ASNS-low | [17] | |
| KRAS-mutated | Gln and Asn dependencies often observed | [111] | ||
| Colorectal cancer (CRC) | KRAS-mutated | Rapamycin | Suppression KRAS-mutated CRC | [112] |
| WNT-mutated | Decreased Asn recycling increases susceptibility to L-ASNase | [113] | ||
| Metastatic breast cancer | Asn bioavailability governs metastatic potential | [20] | ||
| Hepatocellular carcinoma | ASNS-low | Low ASNS associated with worse prognosis, increased invasion, and metastatic potential but increased sensitivity to L-ASNase | [114] | |
| ASNS promoter methylation | Hypermethylation inhibits acquisition of resistance to L-ASNase | [115] | ||
| Glioblastoma | Temozolomide | Potential synergistic effect in vivo. Not confirmed yet if due to Asn/Gln depletion | [116] | |
| 6-Diazo-5-oxo-l-norleucine | Synergistic anti-proliferative effect in glioma | [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Trimpont, M.; Peeters, E.; De Visser, Y.; Schalk, A.M.; Mondelaers, V.; De Moerloose, B.; Lavie, A.; Lammens, T.; Goossens, S.; Van Vlierberghe, P. Novel Insights on the Use of L-Asparaginase as an Efficient and Safe Anti-Cancer Therapy. Cancers 2022, 14, 902. https://doi.org/10.3390/cancers14040902
Van Trimpont M, Peeters E, De Visser Y, Schalk AM, Mondelaers V, De Moerloose B, Lavie A, Lammens T, Goossens S, Van Vlierberghe P. Novel Insights on the Use of L-Asparaginase as an Efficient and Safe Anti-Cancer Therapy. Cancers. 2022; 14(4):902. https://doi.org/10.3390/cancers14040902
Chicago/Turabian StyleVan Trimpont, Maaike, Evelien Peeters, Yanti De Visser, Amanda M. Schalk, Veerle Mondelaers, Barbara De Moerloose, Arnon Lavie, Tim Lammens, Steven Goossens, and Pieter Van Vlierberghe. 2022. "Novel Insights on the Use of L-Asparaginase as an Efficient and Safe Anti-Cancer Therapy" Cancers 14, no. 4: 902. https://doi.org/10.3390/cancers14040902
APA StyleVan Trimpont, M., Peeters, E., De Visser, Y., Schalk, A. M., Mondelaers, V., De Moerloose, B., Lavie, A., Lammens, T., Goossens, S., & Van Vlierberghe, P. (2022). Novel Insights on the Use of L-Asparaginase as an Efficient and Safe Anti-Cancer Therapy. Cancers, 14(4), 902. https://doi.org/10.3390/cancers14040902