
Long-Chain Acyl Coenzyme A Dehydrogenase, a Key Player in Metabolic Rewiring/Invasiveness in Experimental Tumors and Human Mesothelioma Cell Lines

 ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
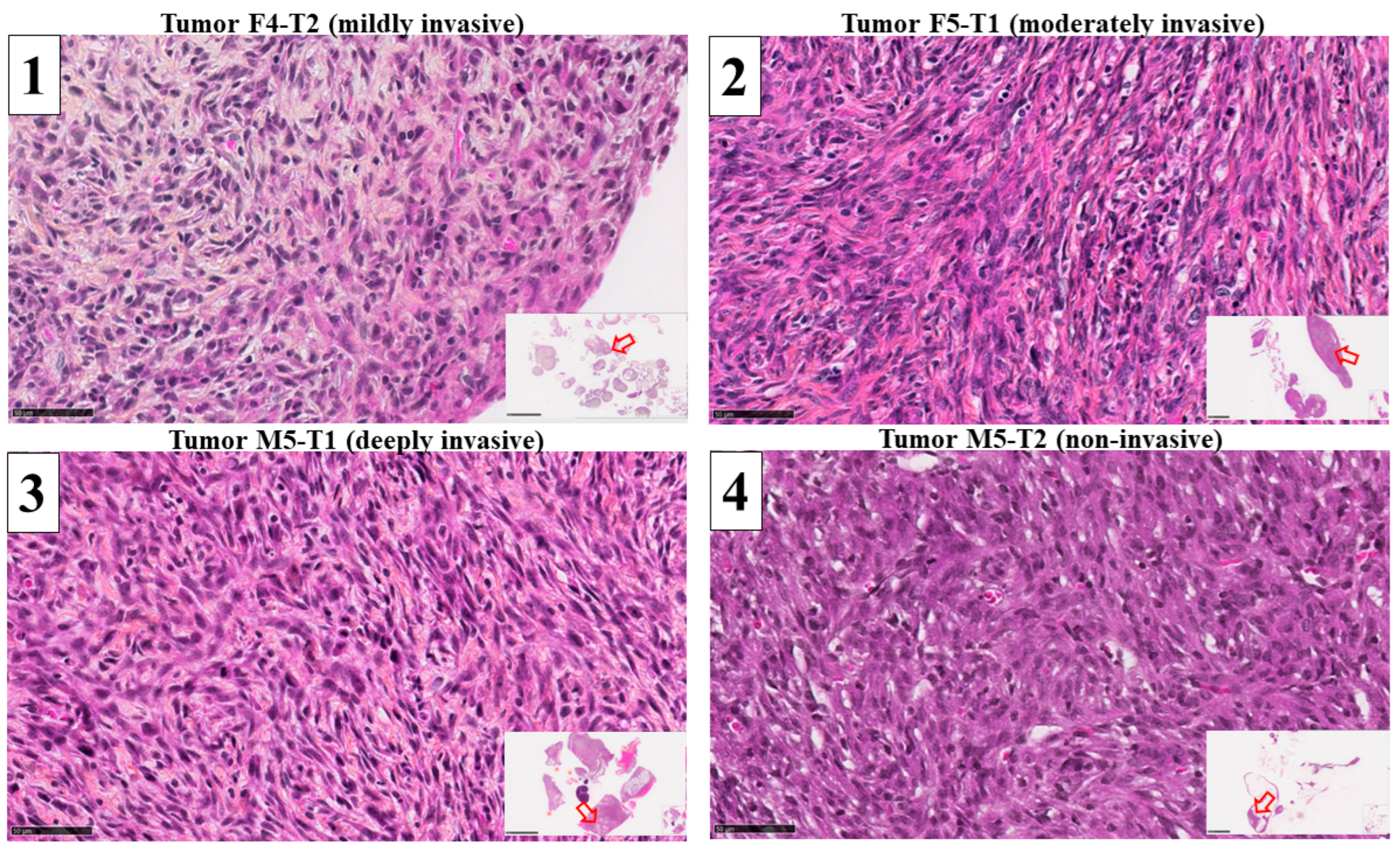
2.1. Collection of Rat Tumor Tissues for Proteomic Analyses
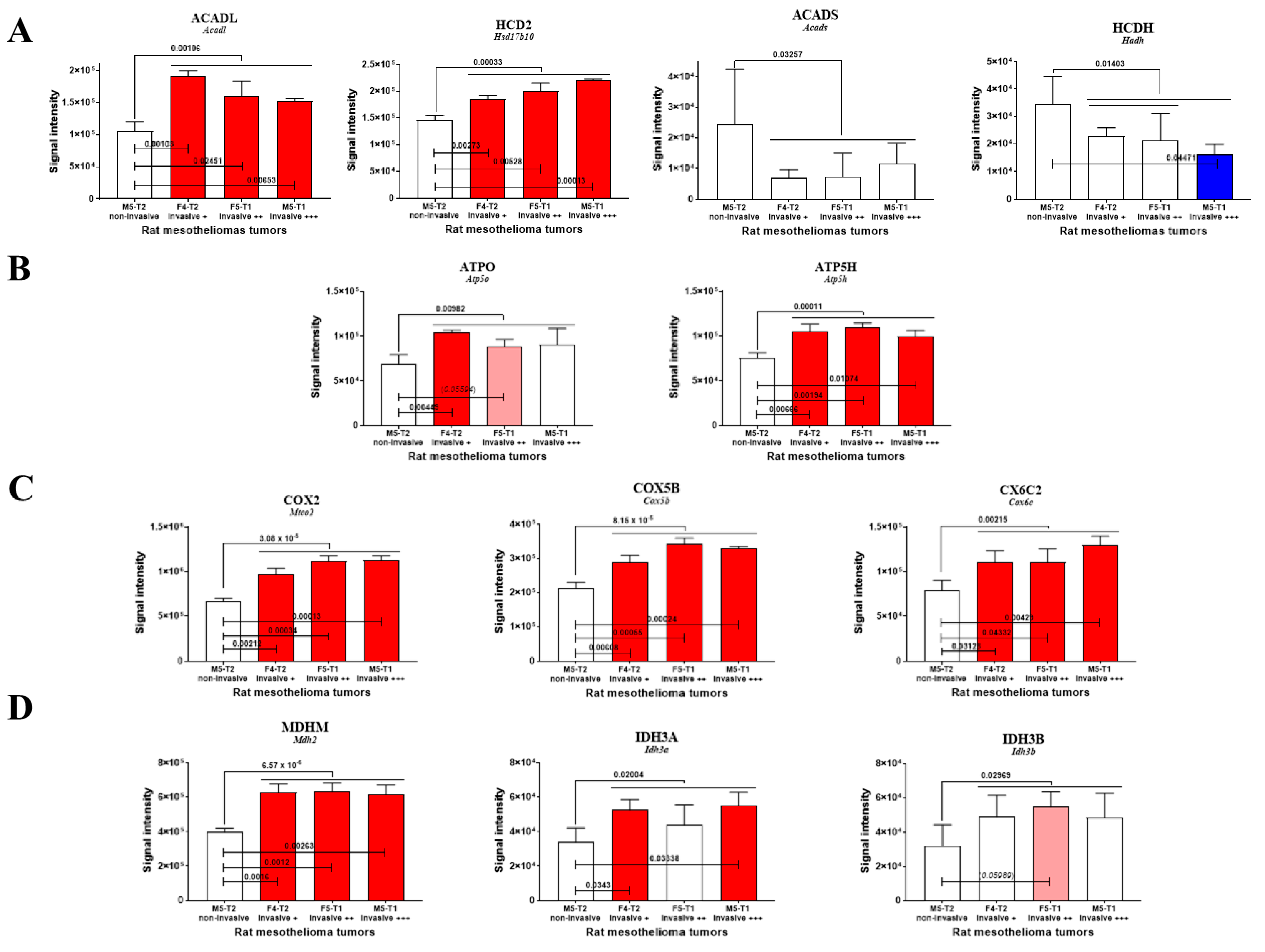
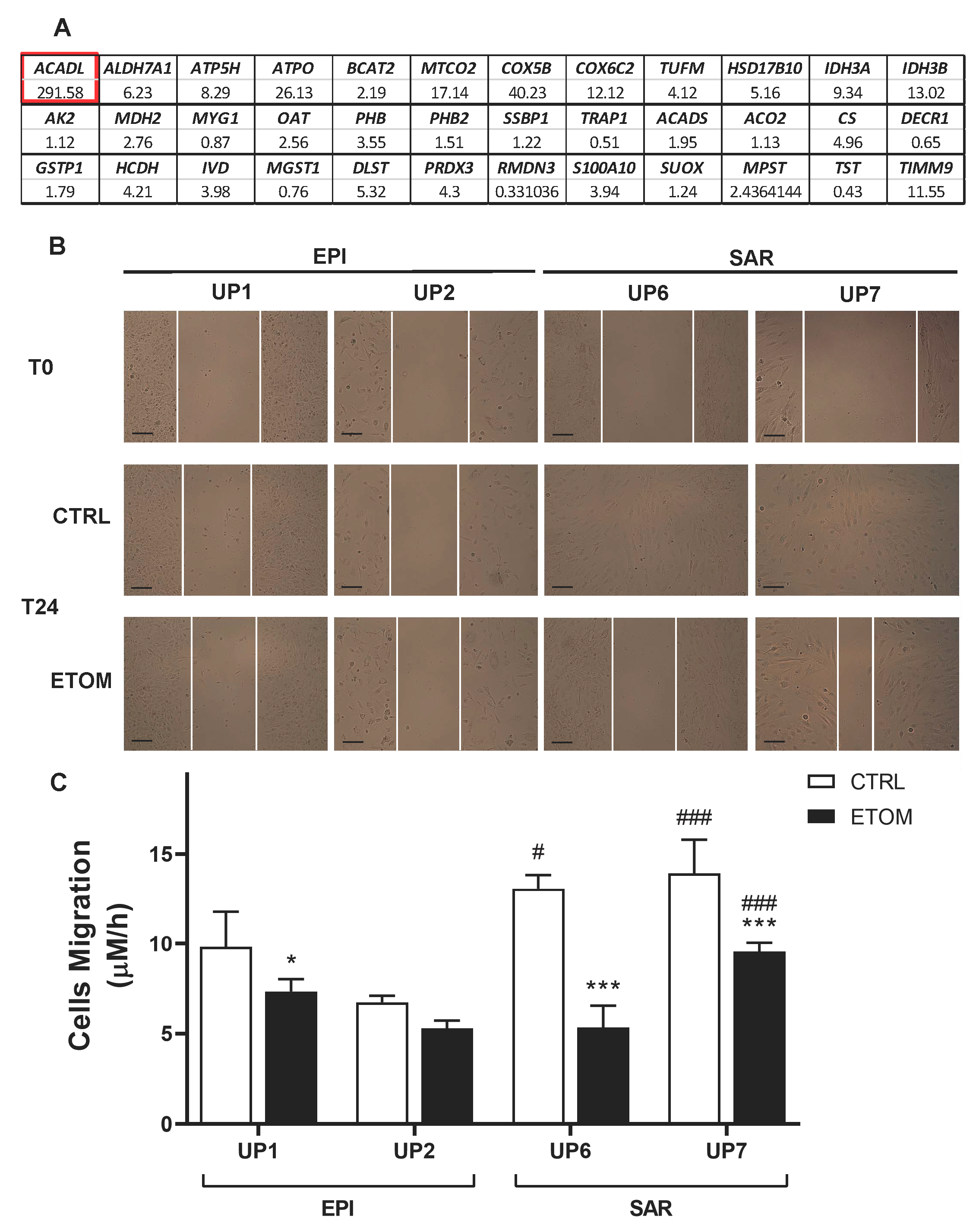
2.2. Proteomic Analyses
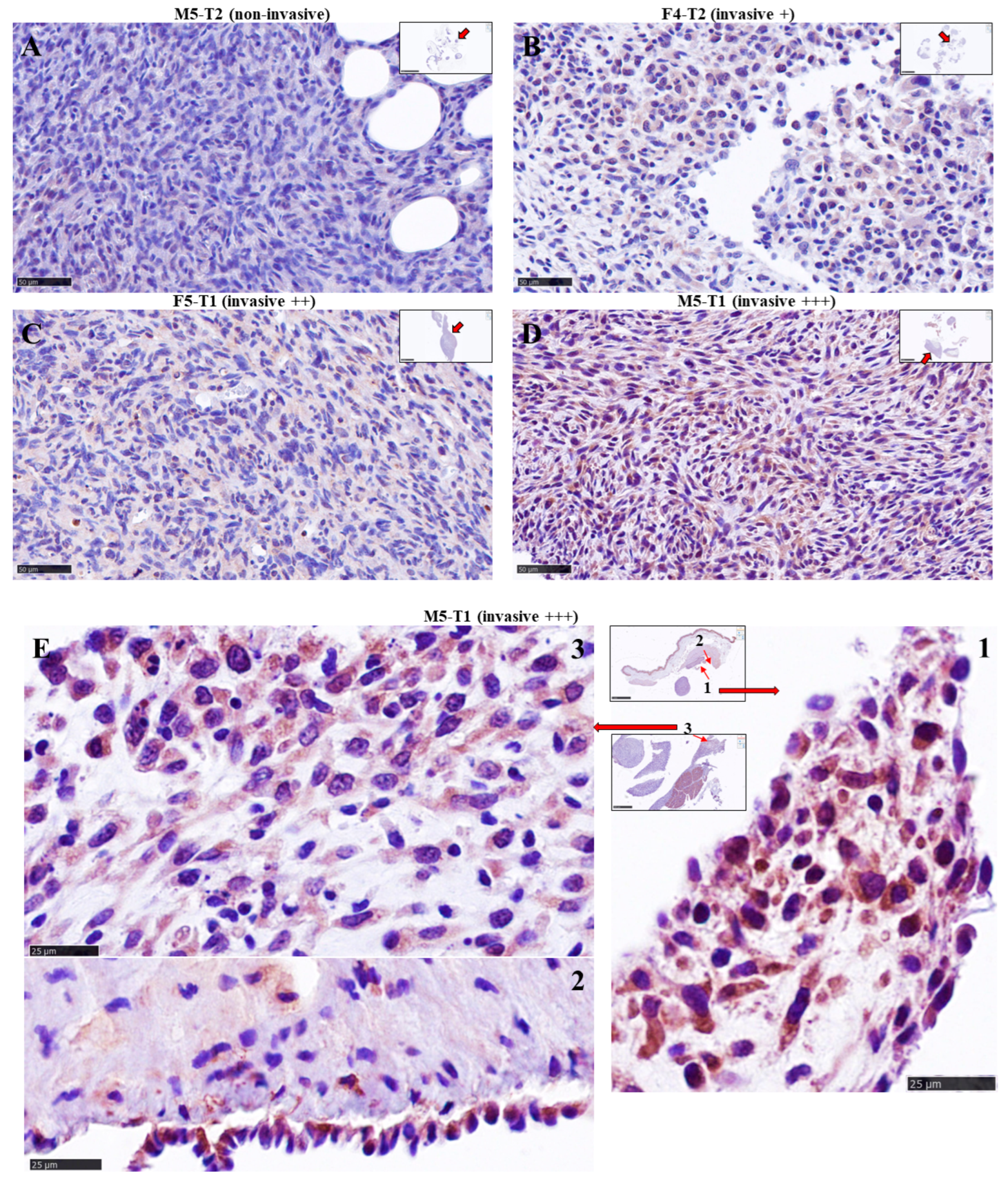
2.3. Histology and Immuno-Histochemical Analyses
2.4. Chemicals
2.5. Cells
2.6. Immunoblotting
2.7. Mitochondria Isolation
2.8. ETC (Electron Transport Chain from Complex I to Complex III)
2.9. ATP
2.10. β-Oxidation of Fatty Acid
2.11. Scratch Assay
2.12. Real Time PCR (RT-PCR)
2.13. Statistical Analysis
3. Results
3.1. Mitochondrial Biomarkers Involved in the Acquisition of Invasiveness in Rat Mesotheliomas
3.2. Immuno-Histochemical Study of ACADL Distribution in Rat Tumors
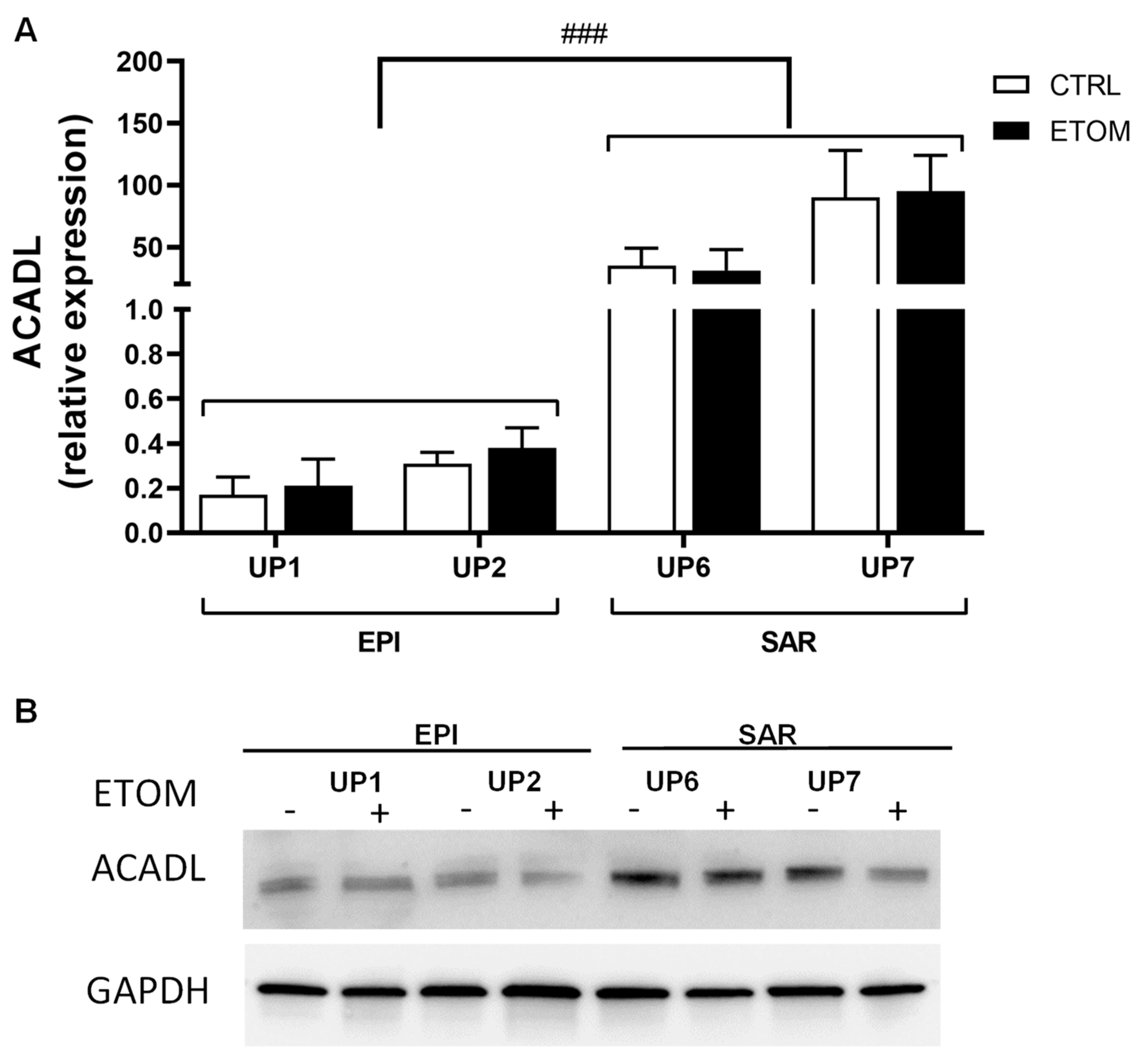
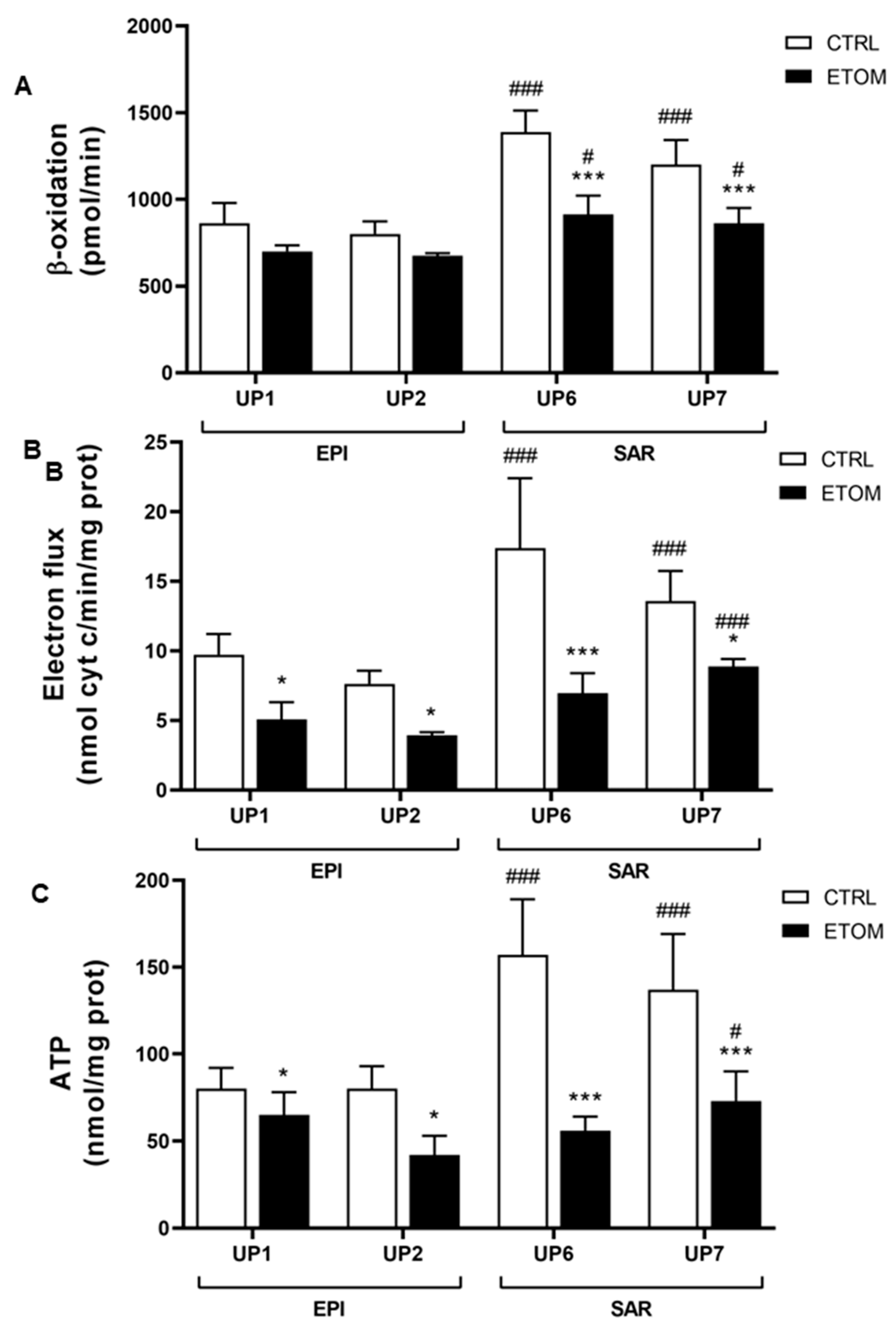
3.3. Fatty Acid β-Oxidation Supports Cell Invasiveness in Human Primary Mesothelioma Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic reprogramming and cancer progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef]
- Scheid, A.D.; Beadnell, T.C.; Welch, D.R. Roles of mitochondria in the hallmarks of metastasis. Br. J. Cancer 2021, 124, 124–135. [Google Scholar] [CrossRef]
- Ishikawa, K.; Takenaga, K.; Akimoto, M.; Koshikawa, N.; Yamaguchi, A.; Imanishi, H.; Nakada, K.; Honma, Y.; Hayashi, J.-I. ROS-generating mitochondrial DNA mutations can regulate tumor cell metastasis. Science 2008, 320, 661–664. [Google Scholar] [CrossRef]
- Zampieri, L.X.; Silva-Almeida, C.; Rondeau, J.D.; Sonveaux, P. Mitochondrial transfer in cancer: A comprehensive review. Int. J. Mol. Sci. 2021, 22, 3245. [Google Scholar] [CrossRef]
- Yanes, B.; Rainero, E. The interplay between cell-extracellular matrix interaction and mitochondria dynamics in cancer. Cancers 2022, 14, 1433. [Google Scholar] [CrossRef] [PubMed]
- Boulton, D.P.; Caino, M.C. Mitochondrial fission and fusion in tumor progression to metastasis. Front. Cell Dev. Biol. 2022, 10, 849962. [Google Scholar] [CrossRef] [PubMed]
- Bononi, G.; Masoni, S.; Di Bussolo, V.; Tuccinardi, T.; Granchi, C.; Minutolo, F. Historical perspective of tumor glycolysis: A century with Otto Warburg. Semin. Cancer Biol. 2022, 86, 325–333. [Google Scholar] [CrossRef]
- Akman, M.; Belisario, D.C.; Salaroglio, I.C.; Kopecka, J.; Donadelli, M.; De Smaele, E.; Riganti, C. Hypoxia, endoplasmic reticulum stress and chemoresistance: Dangerous liaisons. J. Exp. Clin. Cancer Res. 2021, 40, 28. [Google Scholar] [CrossRef] [PubMed]
- Fiorillo, M.; Ózsvári, B.; Sotgia, F.; Lisanti, M.P. High ATP production fuels cancer drug resistance and metastasis: Implications for mitochondrial ATP depletion therapy. Front. Oncol. 2021, 11, 740720. [Google Scholar] [CrossRef]
- Kopecka, J.; Gazzano, E.; Castella, B.; Salaroglio, I.C.; Mungo, E.; Massaia, M.; Riganti, C. Mitochondrial metabolism: Inducer or therapeutic target in tumor immune-resistance? Semin. Cell Dev. Biol. 2020, 98, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zhou, T.; Xie, Y.; Bode, A.M.; Cao, Y. Mitochondria-shaping proteins and chemotherapy. Front. Oncol. 2021, 11, 769036. [Google Scholar] [CrossRef]
- Concolino, A.; Olivo, E.; Tammè, L.; Fiumara, C.; De Angelis, M.T.; Quaresima, B.; Agosti, V.; Costanzo, F.S.; Cuda, G.; Scumaci, D. Proteomics analysis to assess the role of mitochondria in BRCA1-mediated breast tumorigenesis. Proteomes 2018, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Arif, T.; Stern, O.; Pittala, S.; Chalifa-Caspi, V.; Shoshan-Barmatz, V. Rewiring of cancer cell metabolism by mitochondrial VDAC1 depletion results in time-dependent tumor reprogramming: Glioblastoma as a proof of concept. Cells 2019, 8, 1330. [Google Scholar] [CrossRef] [PubMed]
- Nader, J.S.; Boissard, A.; Henry, C.; Valo, I.; Verrièle, V.; Grégoire, M.; Coqueret, O.; Guette, C.; Pouliquen, D.L. Cross-species proteomics identifies CAPG and SBP1 as crucial invasiveness biomarkers in rat and human malignant mesothelioma. Cancers 2020, 12, 2430. [Google Scholar] [CrossRef]
- Nader, J.S.; Abadie, J.; Deshayes, S.; Boissard, A.; Blandin, S.; Blanquart, C.; Boisgerault, N.; Coqueret, O.; Guette, C.; Grégoire, M.; et al. Characterization of increasing stages of invasiveness identifies stromal/cancer cell crosstalk in rat models of mesothelioma. Oncotarget 2018, 9, 16311–16329. [Google Scholar] [CrossRef]
- Yu, D.-L.; Li, H.-W.; Wang, Y.; Li, C.-Q.; You, D.; Jiang, L.; Song, Y.-P.; Li, X.-H. Acyl-CoA dehydrogenase long chain expression is associated with esophageal squamous cell carcinoma progression and poor prognosis. OncoTargets Ther. 2018, 11, 7643–7653. [Google Scholar] [CrossRef]
- Salas, S.; Jézéquel, P.; Campion, L.; Deville, J.-L.; Chibon, F.; Bartoli, C.; Gentet, J.-C.; Charbonnel, C.; Gouraud, W.; Voutsinos-Porche, B.; et al. Molecular characterization of the response to chemotherapy in conventional osteosarcomas: Predictive value of HSD17B10 and IFITM2. Int. J. Cancer 2009, 125, 851–860. [Google Scholar] [CrossRef]
- Klepinin, A.; Zhang, S.; Klepinina, L.; Rebane-Klemm, E.; Terzic, A.; Kaambre, T.; Dzeja, P. Adenylate kinase and metabolic signaling in cancer cells. Front. Oncol. 2020, 10, 660. [Google Scholar] [CrossRef]
- Yusenko, M.V.; Ruppert, T.; Kovacs, G. Analysis of differentially expressed mitochondrial proteins in chromophobe renal cell carcinomas and renal oncocytomas by 2-D gel electrophoresis. Int. J. Biol. Sci. 2010, 6, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Wiebringhaus, R.; Pecoraro, M.; Neubauer, H.A.; Trachtova, K.; Trimmel, B.; Wieselberg, M.; Pencik, J.; Egger, G.; Krall, C.; Moriggl, R.; et al. Proteomic analysis identifies NDUFS1 and ATP5O as novel markers for survival outcome in prostate cancer. Cancers 2021, 13, 6036. [Google Scholar] [CrossRef]
- Lamb, R.; Ozsvari, B.; Bonuccelli, G.; Smith, D.L.; Pestell, R.G.; Martinez-Outschoorn, U.E.; Clarke, R.B.; Sotgia, F.; Lisanti, M.P. Dissecting tumor metabolic heterogeneity: Telomerase and large cell size metabolically define a sub-population of stem-like, mitochondrial-rich, cancer cells. Oncotarget 2015, 6, 21892–21905. [Google Scholar] [CrossRef]
- Zeng, L.; Morinibu, A.; Kobayashi, M.; Zhu, Y.; Wang, X.; Goto, Y.; Yeom, C.J.; Zhao, T.; Hirota, K.; Shinomiya, K.; et al. Aberrant IDH3α expression promotes malignant tumor growth by inducing HIF-1-mediated metabolic reprogramming and angiogenesis. Oncogene 2015, 34, 4758–4766. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Harvey, C.T.; Geng, H.; Xue, C.; Chen, V.; Beer, T.M.; Qian, D.Z. Malate dehydrogenase 2 confers docetaxel resistance via regulations of JNK signaling and oxidative metabolism. Prostate 2013, 73, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Bavelloni, A.; Piazzi, M.; Raffini, M.; Faenza, I.; Blalock, W.L. Prohibitin 2: At a communications crossroads. IUBMB Life 2015, 67, 239–254. [Google Scholar] [CrossRef]
- Wang, B.; Ao, J.; Yu, D.; Rao, T.; Ruan, Y.; Yao, X. Inhibition of mitochondrial translation effectively sensitizes renal cell carcinoma to chemotherapy. Biochem. Biophys. Res. Commun. 2017, 490, 767–773. [Google Scholar] [CrossRef]
- Xu, S.; Wu, Y.; Chen, Q.; Cao, J.; Hu, K.; Tang, J.; Sang, Y.; Lai, F.; Wang, L.; Zhang, R.; et al. hSSB1 regulates both the stability and the transcriptional activity of p53. Cell Res. 2013, 23, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.; Bhattacharya, R.; Mukherjee, P. Hydrogen sulfide signaling in mitochondria and disease. FASEB J. 2019, 33, 13098–13125. [Google Scholar] [CrossRef] [PubMed]
- Ismail, T.; Kim, Y.; Lee, H.; Lee, D.-S.; Lee, H.-S. Interplay between mitochondrial peroxiredoxins and ROS in cancer development and progression. Int. J. Mol. Sci. 2019, 20, 4407. [Google Scholar] [CrossRef]
- Seo, J.-S.; Svenningsson, P. Modulation of ion channels and receptors by p11 (S100A10). Trends Pharmacol. Sci. 2020, 41, 487–497. [Google Scholar] [CrossRef]
- Kim, T.-M.; Yang, S.; Seung, B.-J.; Lee, S.; Kim, D.; Ha, Y.-J.; Seo, M.-k.; Kim, K.-K.; Kim, H.S.; Cheong, J.-H.; et al. Cross-species oncogenic signatures of breast cancer in canine mammary tumors. Nat. Commun. 2020, 11, 3616. [Google Scholar] [CrossRef]
- Al-Harazi, O.; Kaya, I.H.; Al-Eid, M.; Alfantoukh, L.; Al Zahrani, A.S.; Al Sebayel, M.; Kaya, N.; Colak, D. Identification of gene signature as diagnostic and prognostic blood biomarker for early hepatocellular carcinoma using integrated cross-species transcriptomic and network analyses. Front. Genet. 2021, 12, 710049. [Google Scholar] [CrossRef] [PubMed]
- Rensch, T.; Villar, D.; Horvath, J.; Odom, D.T.; Flicek, P. Mitochondrial heteroplasmy in vertebrates using ChIP-sequencing data. Genome Biol. 2016, 17, 139. [Google Scholar] [CrossRef]
- Spinelli, J.B.; Haigis, M.C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 2018, 20, 745–754. [Google Scholar] [CrossRef]
- Carracedo, A.; Cantley, L.C.; Pandolfi, P.P. Cancer metabolism: Fatty acid oxidation in the limelight. Nat. Rev. Cancer 2013, 13, 227–232. [Google Scholar] [CrossRef]
- Xie, B.-X.; Zhang, H.; Wang, J.; Pang, B.; Wu, R.-Q.; Qian, X.-L.; Yu, L.; Li, S.-H.; Shi, Q.-G.; Huang, C.-F.; et al. Analysis of differentially expressed genes in LNCaP prostate cancer progression model. J. Androl. 2011, 32, 170–182. [Google Scholar] [CrossRef]
- Carlson, E.A.; Marquez, R.T.; Du, F.; Wang, Y.; Xu, L.; Yan, S.S. Overexpression of 17b-hydroxysteroid dehydrogenase type 10 increases pheochromocytoma cell growth and resistance to cell death. BMC Cancer 2015, 15, 166. [Google Scholar] [CrossRef] [PubMed]
- Condon, K.J.; Orozco, J.M.; Adelman, C.H.; Spinelli, J.B.; van der Helm, P.W.; Roberts, J.M.; Kunchok, T.; Sabatini, D.M. Genome-wide CRISPR screens reveal multitiered mechanisms through which mTORC1 senses mitochondrial dysfunction. Proc. Natl. Acad. Sci. USA 2021, 118, e2022120118. [Google Scholar] [CrossRef]
- Braun, C.; Weichhart, T. mTOR-dependent immunometabolism as Achilles’ heel of anticancer therapy. Eur. J. Immunol. 2021, 51, 3161–3175. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ma, F.; Qian, H.-l. Defueling the cancer: ATP synthase as an emerging target in cancer therapy. Mol. Ther. Oncolytics 2021, 23, 82–95. [Google Scholar] [CrossRef]
- Galber, C.; Acosta, M.J.; Minervivi, G.; Giorgio, V. The role of mitochondrial ATP synthase in cancer. Biol. Chem. 2020, 401, 1199–1214. [Google Scholar] [CrossRef]
- Chang, H.J.; Lee, M.R.; Hong, S.-H.; Yoo, B.C.; Shin, Y.-K.; Jeong, J.Y.; Lim, S.-B.; Choi, H.S.; Park, J.-G. Identification of mitochondrial F0F1-ATP synthase involved in liver metastasis of colorectal cancer. Cancer Sci. 2007, 98, 1184–1191. [Google Scholar] [CrossRef]
- Gaude, E.; Frezza, C. Defects in mitochondrial metabolism and cancer. Cancer Metab. 2014, 2, 10. [Google Scholar] [CrossRef]
- Gao, S.-P.; Sun, H.-F.; Fu, W.-Y.; Li, L.-D.; Zhao, Y.; Chen, M.-T.; Jin, W. High expression of COX5B is associated with poor prognosis in breast cancer. Future Oncol. 2017, 13, 1711–1719. [Google Scholar] [CrossRef]
- Chu, Y.-D.; Lin, W.-R.; Lin, Y.-H.; Kuo, W.-H.; Tseng, C.-J.; Lim, S.-N.; Huang, Y.-L.; Huang, S.-C.; Wu, T.-J.; Lin, K.-H.; et al. COX5B-mediated bioenergetic alteration regulates tumor growth and migration by modulating AMPK-UHMK1-ERK cascade in hepatoma. Cancers 2020, 12, 1646. [Google Scholar] [CrossRef]
- Chu, Y.-D.; Lim, S.-N.; Yeh, C.-T.; Lin, W.-R. COX5B-mediated bioenergetic alterations modulate cell growth and anticancer drug susceptibility by orchestrating claudin-2 expression in colorectal cancers. Biomedicines 2022, 10, 60. [Google Scholar] [CrossRef]
- Tian, B.-X.; Sun, W.; Wang, S.-H.; Liu, P.-J.; Wang, Y.-C. Differential expression and clinical significance of COX6C in human diseases. Am. J. Transl. Res. 2021, 13, 1–10. [Google Scholar]
- Jang, S.C.; Crescitelli, R.; Cvjetkovic, A.; Belgrano, V.; Bagge, R.O.; Sundfeldt, K.; Ochiya, T.; Kalluri, R.; Lötvall, J. Mitochondrial protein enriched extracellular vesicles discovered in human melanoma tissues can be detected in patient plasma. J. Extracell. Vesicles 2019, 8, 1635420. [Google Scholar] [CrossRef]
- Laurenti, G.; Tennant, D.A. Isocitrate dehydrogenase (IDH), succinate dehydrogenase (SDH), fumarate hydratase (FH): Three players for one phenotype in cancer? Biochem. Soc. Trans. 2016, 44, 1111–1116. [Google Scholar] [CrossRef]
- Cruz, I.N.; Coley, H.M.; Kramer, H.B.; Madhuri, T.K.; Safuwan, N.A.M.; Angelino, A.R.; Yang, M. Proteomics analysis of ovarian cancer cell lines and tissues reveals drug resistance-associated proteins. Cancer Genom. Proteom. 2017, 14, 35–52. [Google Scholar] [CrossRef]
- Norberg, E.; Lako, A.; Chen, P.-H.; Stanley, I.A.; Zhou, F.; Ficarro, S.B.; Chapuy, B.; Chen, L.; Rodig, S.; Shin, D.; et al. Differential contribution of the mitochondrial translation pathway to the survival of diffuse large B-cell lymphoma subsets. Cell Death Differ. 2017, 24, 251–262. [Google Scholar] [CrossRef]
- Chatla, S.; Du, W.; Wilson, A.F.; Meetei, A.R.; Pang, Q. Fancd2-deficient hematopoietic stem and progenitor cells depend on augmented mitochondrial translation for survival and proliferation. Stem Cell Res. 2019, 40, 101550. [Google Scholar] [CrossRef]
- Brocker, C.; Cantore, M.; Failli, P.; Vasiliou, V. Aldehyde dehydrogenase 7A1 (ALDH7A1) attenuates reactive aldehyde and oxidative stress induced cytotoxicity. Chem. Biol. Interact. 2011, 191, 269–277. [Google Scholar] [CrossRef]
- Prabhu, V.V.; Lulla, A.R.; Madhukar, N.S.; Ralff, M.D.; Zhao, D.; Kline, C.L.B.; Van den Heuvel, A.P.; Lev, A.; Garnett, M.J.; McDermott, U.; et al. Cancer stem cell-related gene expression as a potential biomarker of response for first-in-class imipridone ONC201 in solid tumors. PLoS ONE 2017, 12, e0180541. [Google Scholar] [CrossRef]
- Bizzaro, V.; Belvedere, R.; Milone, M.R.; Pucci, B.; Lombardi, R.; Bruzzese, F.; Popolo, A.; Parente, L.; Budillon, A.; Petrella, A. Annexin A1 is involved in the acquisition and maintenance of a stem cell-like/aggressive phenotype in prostate cancer cells with acquired resistance to zoledronic acid. Oncotarget 2015, 6, 25074–25092. [Google Scholar] [CrossRef]
- Van den Hoogen, C.; van der Horst, G.; Cheung, H.; Buijs, J.T.; Pelger, R.C.M.; van der Pluijm, G. The aldehyde dehydrogenase enzyme 7A1 is functionally involved in prostate cancer bone metastasis. Clin. Exp. Metastasis 2011, 28, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Giacalone, N.J.; Den, R.B.; Eisenberg, R.; Chen, H.; Olson, S.J.; Massion, P.P.; Carbone, D.P.; Lu, B. ALDH7A1 expression is associated with recurrence in patients with surgically resected non-small-cell lung carcinoma. Future Oncol. 2013, 9, 737–745. [Google Scholar] [CrossRef]
- Lee, J.-S.; Lee, H.; Woo, S.M.; Jang, H.; Jeon, Y.; Kim, H.Y.; Song, J.; Lee, W.J.; Hong, E.K.; Park, S.-J.; et al. Overall survival of pancreatic ductal adenocarcinoma is doubled by Aldh7a1 deletion in the KPC mouse. Theranostics 2021, 11, 3472–3488. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.W.-Y.; Chan, C.-L.; Tang, W.-K.; Cheng, C.H.-K.; Fong, W.-P. Is antiquitin a mitochondrial enzyme? J. Cell. Biochem. 2010, 109, 74–81. [Google Scholar] [CrossRef]
- Babbi, G.; Baldazzi, D.; Savojardo, C.; Martelli, P.L.; Casadio, R. Highlighting human enzymes active in different metabolic pathways and diseases: The case study of EC 1.2.3.1 and EC 2.3.1.9. Biomedicines 2020, 8, 250. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UNP (Number) | Histotype | Gender | Age (Years) | Asbestos Exposure | First Line of Treatment | Second Line of Treatment | TTP (Months) | OS (Months) |
|---|---|---|---|---|---|---|---|---|
| 1 | Epithelioid | M | 74 | Unknown | Carbo + Pem | No | 7 | 11 |
| 2 | Epithelioid | F | 58 | Yes | Carbo + Pem | Pem | 6 | 13 |
| 3 | Epithelioid | M | 76 | Unknown | CisPt + Pem | No | 3 | 8 |
| 4 | Epithelioid | M | 68 | Yes | Carbo + Pem | Pem | 4 | 9 |
| 5 | Epithelioid | F | 84 | Yes | CisPt + Pem | No | 7 | 8 |
| 6 | Sarcomatoid | M | 80 | Yes | Carbo + Pem | Trabectedin | 3 | 5 |
| 7 | Sarcomatoid | F | 78 | Unknown | Pem | No | 4 | 6 |
| 8 | Sarcomatoid | M | 69 | Yes | Carbo + Pem | Trabectedin | 7 | 10 |
| 9 | Sarcomatoid | F | 74 | Unknown | Carbo + Pem | No | 5 | 7 |
| 10 | Sarcomatoid | M | 78 | Yes | Carbo + Pem | Trabectedin | 4 | 9 |
| Code # | Gene # | Full Name # | [1 + 2 + 3] vs. 4 |
|---|---|---|---|
| ACADL | Acadl | Long-chain specific acyl-CoA dehydrogenase, mitochondrial | ↑ |
| AL7A1 * | Aldh7a1 | Alpha-aminoadipic semialdehyde dehydrogenase | ↑ |
| ATP5H | Atp5h | ATP synthase subunit d, mitochondrial | ↑ |
| ATPO | Atp5o | ATP synthase subunit O, mitochondrial | ↑ |
| BCAT2 * | Bcat2 | Branched-chain-amino-acid aminotransferase, mitochondrial | ↑ |
| COX2 | Mtco2 | Cytochrome c oxidase subunit 2 | ↑ |
| COX5B | Cox5b | Cytochrome c oxidase subunit 5B, mitochondrial | ↑ |
| CX6C2 | Cox6c2 | Cytochrome c oxidase subunit 6C-2 | ↑ |
| EFTU | Tufm | Elongation factor Tu, mitochondrial | ↑ |
| HCD2 | Hsd17b10 | 3-hydroxyacyl-CoA dehydrogenase type-2 | ↑ |
| IDH3A | Idh3a | Isocitrate dehydrogenase [NAD] subunit alpha, mitochondrial | ↑ |
| IDH3B | Idh3b | Isocitrate dehydrogenase [NAD] subunit beta, mitochondrial | ↑ |
| KAD2 | Ak2 | Adenylate kinase 2, mitochondrial | ↑ |
| MDHM | Mdh2 | Malate dehydrogenase, mitochondrial | ↑ |
| MYG1 * | Myg1 | UPF0160 protein MYG1, mitochondrial | ↑ |
| OAT * | Oat | Ornithine aminotransferase, mitochondrial | ↑ |
| PHB | Phb | Prohibitin | ↑ |
| PHB2 | Phb2 | Prohibitin-2 | ↑ |
| SSBP | Ssbp1 | Single-stranded DNA-binding protein, mitochondrial | ↑ |
| TRAP1 | Trap1 | Heat shock protein 75 kDa, mitochondrial | ↑ |
| ACADS | Acads | Short-chain specific acyl-CoA dehydrogenase, mitochondrial | ↓ |
| ACON | Aco2 | Aconitate hydratase, mitochondrial | ↓ |
| CISY * | Cs | Citrate synthase, mitochondrial | ↓ |
| DECR * | Decr1 | 2, 4 dienoyl-CoA reductase, mitochondrial | ↓ |
| GSTP1 * | Gstp1 | Glutathione S-transferase P | ↓ |
| HCDH | Hadh | Hydroxyacyl-CoA dehydrogenase, mitochondrial | ↓ |
| IVD * | Ivd | Isovaleryl-CoA dehydrogenase, mitochondrial | ↓ |
| MGST1 * | Mgst1 | Microsomal glutathione S-transferase 1 | ↓ |
| ODO2 | Dlst | Dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex, mitochondrial | ↓ |
| PRDX3 | Prdx3 | Thioredoxin-dependent peroxide reductase, mitochondrial | ↓ |
| RMD3 * | Rmdn3 | Regulator of microtubule dynamics protein 3 | ↓ |
| S10AA | S100a10 | Protein S100-A10 | ↓ |
| SUOX | Suox | Sulfite oxidase, mitochondrial | ↓ |
| THTM | Mpst | 3-mercaptopyruvate sulfurtransferase | ↓ |
| THTR | Tst | Thiosulfate sulfurtransferase | ↓ |
| TIM9 | Timm9 | Mitochondrial import inner membrane translocase subunit Tim9 | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouliquen, D.L.; Ortone, G.; Rumiano, L.; Boissard, A.; Henry, C.; Blandin, S.; Guette, C.; Riganti, C.; Kopecka, J. Long-Chain Acyl Coenzyme A Dehydrogenase, a Key Player in Metabolic Rewiring/Invasiveness in Experimental Tumors and Human Mesothelioma Cell Lines. Cancers 2023, 15, 3044. https://doi.org/10.3390/cancers15113044
Pouliquen DL, Ortone G, Rumiano L, Boissard A, Henry C, Blandin S, Guette C, Riganti C, Kopecka J. Long-Chain Acyl Coenzyme A Dehydrogenase, a Key Player in Metabolic Rewiring/Invasiveness in Experimental Tumors and Human Mesothelioma Cell Lines. Cancers. 2023; 15(11):3044. https://doi.org/10.3390/cancers15113044
Chicago/Turabian StylePouliquen, Daniel L., Giacomo Ortone, Letizia Rumiano, Alice Boissard, Cécile Henry, Stéphanie Blandin, Catherine Guette, Chiara Riganti, and Joanna Kopecka. 2023. "Long-Chain Acyl Coenzyme A Dehydrogenase, a Key Player in Metabolic Rewiring/Invasiveness in Experimental Tumors and Human Mesothelioma Cell Lines" Cancers 15, no. 11: 3044. https://doi.org/10.3390/cancers15113044
APA StylePouliquen, D. L., Ortone, G., Rumiano, L., Boissard, A., Henry, C., Blandin, S., Guette, C., Riganti, C., & Kopecka, J. (2023). Long-Chain Acyl Coenzyme A Dehydrogenase, a Key Player in Metabolic Rewiring/Invasiveness in Experimental Tumors and Human Mesothelioma Cell Lines. Cancers, 15(11), 3044. https://doi.org/10.3390/cancers15113044