

The Role of Adhesion Molecules and Extracellular Vesicles in an In Vitro Model of the Blood–Brain Barrier for Metastatic Disease
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Lines, HUVECs and HCMEC/D3 Cultures
2.2. The Blood–Brain Barrier and Migration Assay
2.3. Cell Line Characterization by Flow Cytometry
2.4. EV Isolation
2.5. Characterization of EVs
2.5.1. Super-Resolution Microscopy
2.5.2. Transmission Electron Microscopy
2.5.3. Flow Cytometry
2.6. Quantitative RT-PCR (qRT-PCR)
2.7. Conditioned Culture Media Collection
2.8. Evaluation of Conditioned Medium/EV-Induced Apoptosis in Endothelial Cells
2.9. Statistics
3. Results
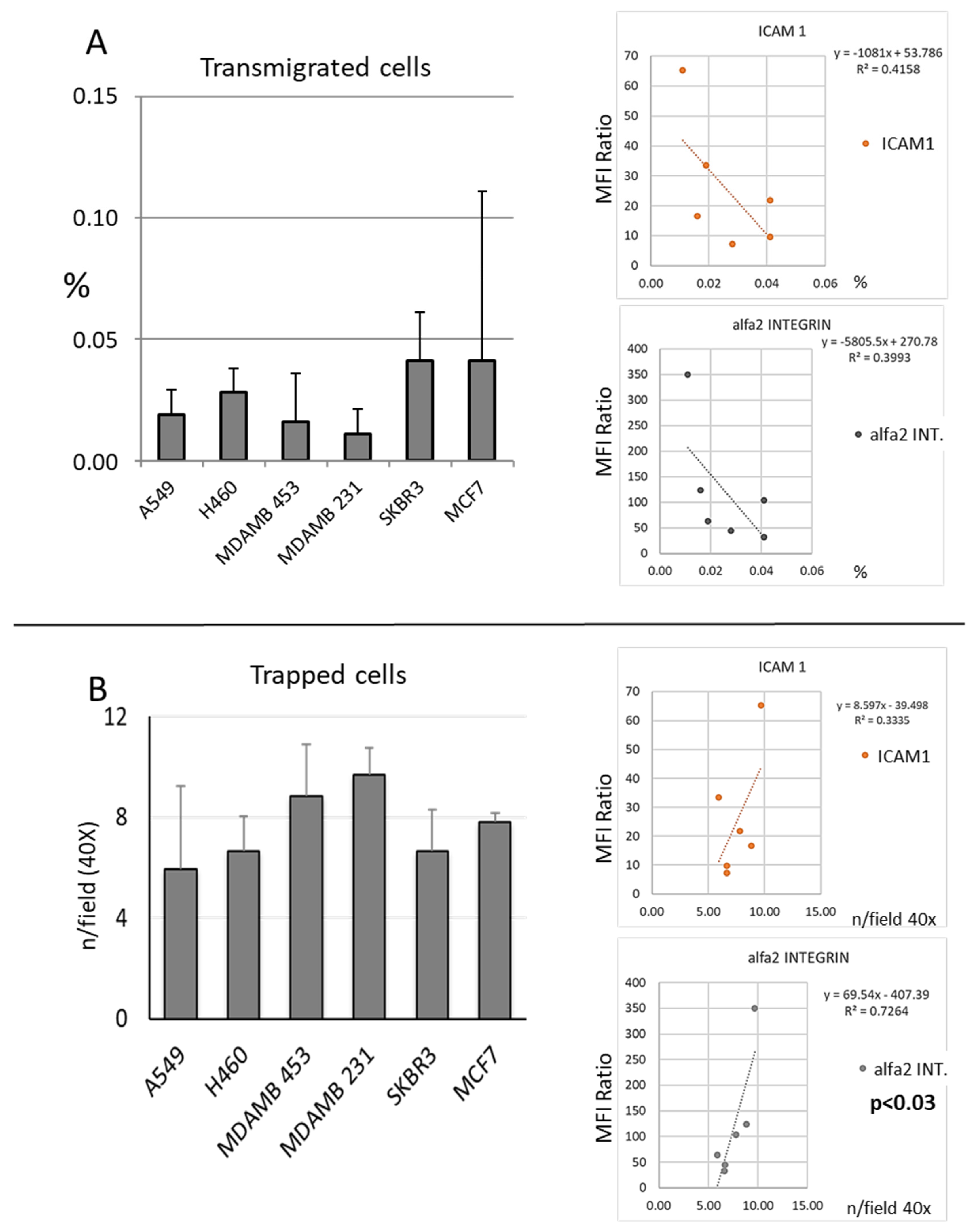
3.1. Migration
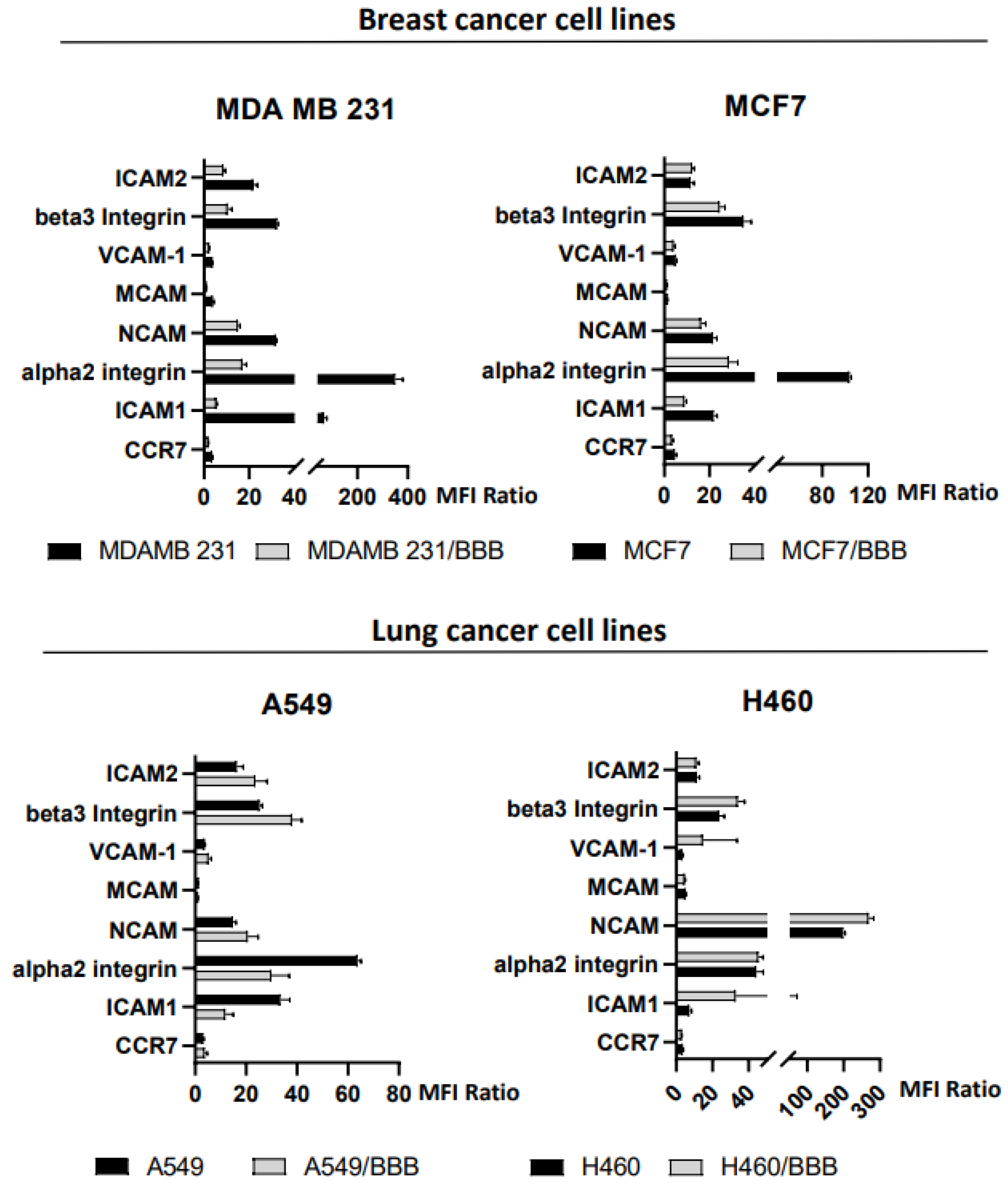
3.2. Expression of Adhesion Molecules
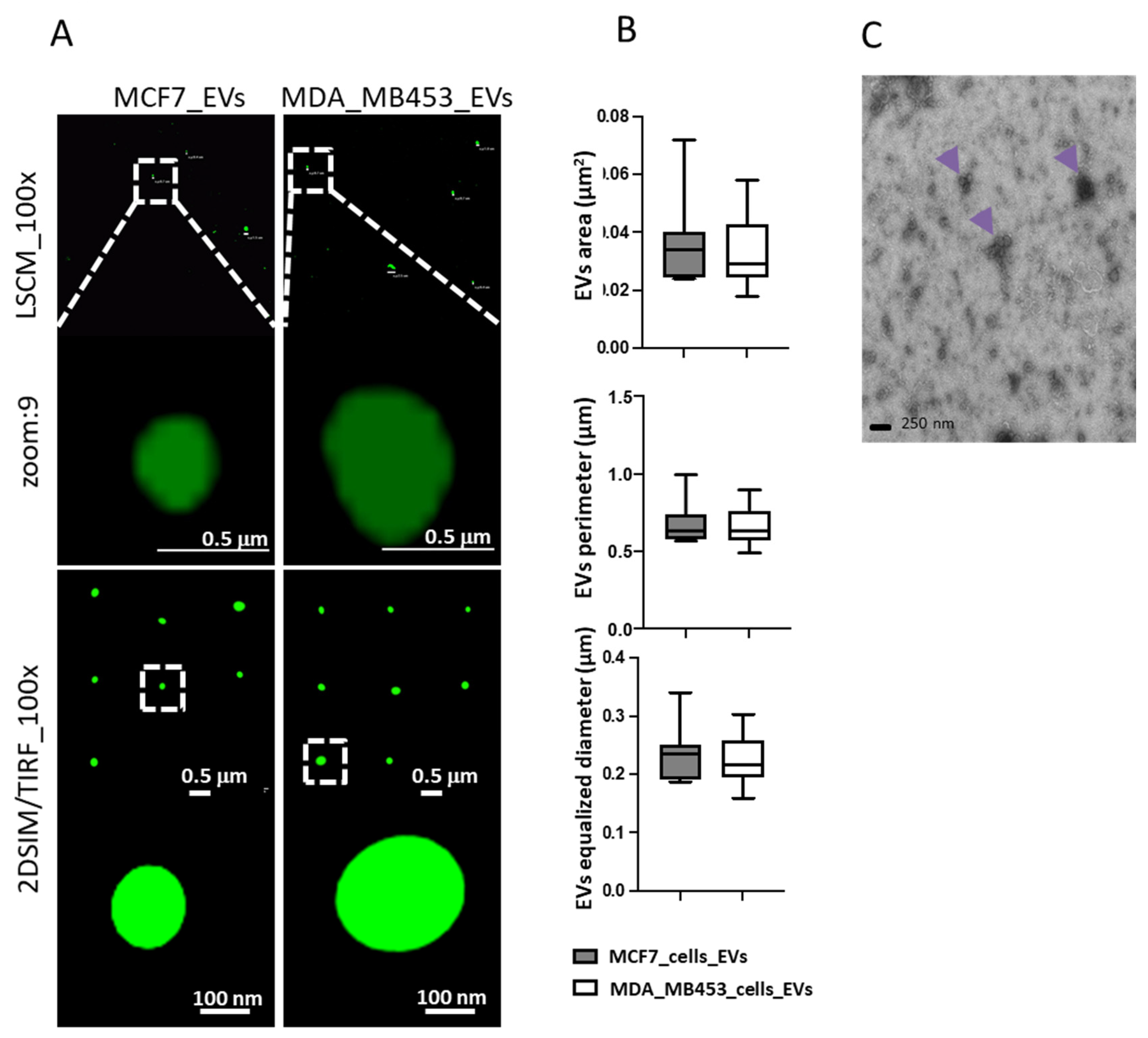
3.3. Characterization of EVs Produced by Tumor Cell Lines
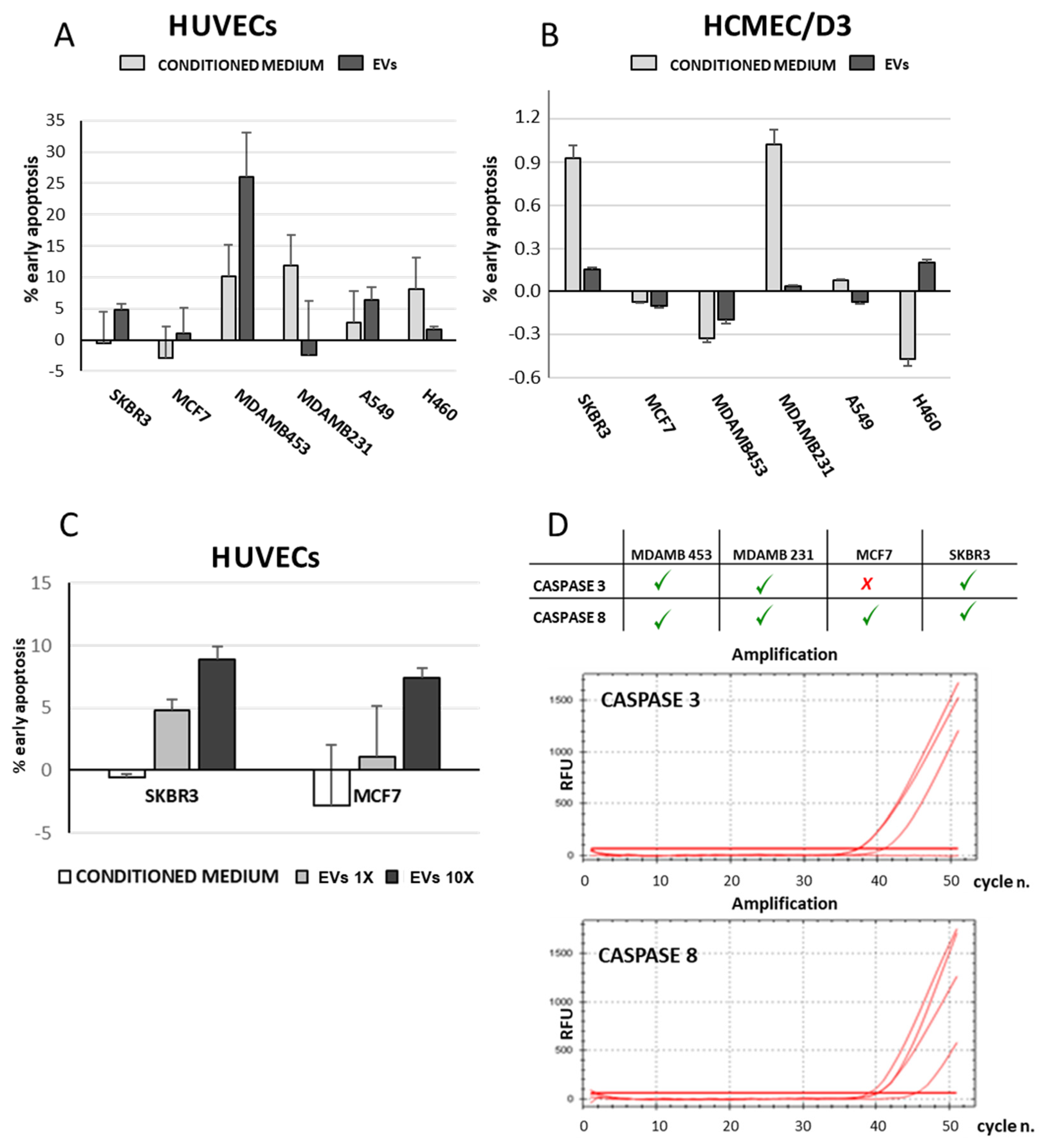
3.4. Isolated EVs Induced Apoptosis of Endothelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaspar, L.; Scott, C.; Rotman, M.; Asbell, S.; Phillips, T.; Wasserman, T.; McKenna, W.G.; Byhardt, R. Recursive partitioning analysis (RPA) of prognostic factors in three Radiation Therapy Oncology Group (RTOG) brain metastases trials. Int. J. Radiat. Oncol. Biol. Phys. 1997, 37, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Sperduto, P.W.; Kased, N.; Roberge, D.; Xu, Z.; Shanley, R.; Luo, X.; Sneed, P.K.; Chao, S.T.; Weil, R.J.; Suh, J.; et al. Summary report on the graded prognostic assessment: An accurate and facile diagnosis-specific tool to estimate survival for patients with brain metastases. J. Clin. Oncol. 2012, 30, 419–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperduto, P.W.; Yang, T.J.; Beal, K.; Pan, H.; Brown, P.D.; Bangdiwala, A.; Shanley, R.; Yeh, N.; Gaspar, L.E.; Braunstein, S.; et al. Estimating Survival in Patients With Lung Cancer and Brain Metastases: An Update of the Graded Prognostic Assessment for Lung Cancer Using Molecular Markers (Lung-molGPA). JAMA Oncol. 2017, 3, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Lamba, N.; Wen, P.Y.; Aizer, A.A. Epidemiology of brain metastases and leptomeningeal disease. Neuro Oncol. 2021, 23, 1447–1456. [Google Scholar] [CrossRef]
- Yamamoto, M.; Serizawa, T.; Shuto, T.; Akabane, A.; Higuchi, Y.; Kawagishi, J.; Yamanaka, K.; Sato, Y.; Jokura, H.; Yomo, S.; et al. Stereotactic radiosurgery for patients with multiple brain metastases (JLGK0901): A multi-institutional prospective observational study. Lancet Oncol. 2014, 15, 387–395. [Google Scholar] [CrossRef]
- Brastianos, P.K.; Carter, S.L.; Santagata, S.; Cahill, D.P.; Taylor-Weiner, A.; Jones, R.T.; Van Allen, E.M.; Lawrence, M.S.; Horowitz, P.M.; Cibulskis, K.; et al. Genomic Characterization of Brain Metastases Reveals Branched Evolution and Potential Therapeutic Targets. Cancer Discov. 2015, 5, 1164–1177. [Google Scholar] [CrossRef] [Green Version]
- Berghoff, A.S.; Brastianos, P.K. Toward Precision Medicine in Brain Metastases. Semin. Neurol. 2018, 38, 95–103. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Huang, T.; Wang, Y.; Song, M.M.; Song, T.; Long, G.; Zhang, X.; Li, X.; Zhang, L. Cerebrospinal fluid circulating tumor DNA depicts profiling of brain metastasis in NSCLC. Mol. Oncol. 2022, 17, 810–824. [Google Scholar] [CrossRef]
- Reymond, N.; d’Água, B.B.; Ridley, A.J. Crossing the endothelial barrier during metastasis. Nat. Rev. Cancer 2013, 13, 858–870. [Google Scholar] [CrossRef]
- Claesson-Welsh, L.; Dejana, E.; McDonald, D.M. Permeability of the Endothelial Barrier: Identifying and Reconciling Controversies. Trends Mol. Med. 2021, 27, 314–331. [Google Scholar] [CrossRef]
- Arvanitis, C.D.; Ferraro, G.B.; Jain, R.K. The blood-brain barrier and blood-tumour barrier in brain tumours and metastases. Nat. Rev. Cancer 2020, 20, 26–41. [Google Scholar] [CrossRef]
- Carretero-González, A.; Hergueta-Redondo, M.; Sánchez-Redondo, S.; Ximénez-Embún, P.; Manso Sánchez, L.; Gil, E.C.; Castellano, D.; de Velasco, G.; Peinado, H. Characterization of plasma circulating small extracellular vesicles in patients with metastatic solid tumors and newly diagnosed brain metastasis. Oncoimmunology 2022, 11, 2067944. [Google Scholar] [CrossRef]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Popper, H.H. Progression and metastasis of lung cancer. Cancer Metastasis Rev. 2016, 35, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J. Clin. Investig. 1973, 52, 2745–2756. [Google Scholar] [CrossRef]
- Rizzo, A.; Vasco, C.; Girgenti, V.; Fugnanesi, V.; Calatozzolo, C.; Canazza, A.; Salmaggi, A.; Rivoltini, L.; Morbin, M.; Ciusani, E. Melanoma cells homing to the brain: An in vitro model. Biomed Res. Int. 2015, 2015, 476069. [Google Scholar] [CrossRef] [Green Version]
- Panachan, J.; Rojsirikulchai, N.; Pongsakul, N.; Khowawisetsut, L.; Pongphitcha, P.; Siriboonpiputtana, T.; Chareonsirisuthigul, T.; Phornsarayuth, P.; Klinkulab, N.; Jinawath, N.; et al. Extracellular Vesicle-Based Method for Detecting. Cancers 2022, 14, 2627. [Google Scholar] [CrossRef]
- Gállego Pérez-Larraya, J.; Hildebrand, J. Brain metastases. Handb. Clin. Neurol. 2014, 121, 1143–1157. [Google Scholar]
- Achrol, A.S.; Rennert, R.C.; Anders, C.; Soffietti, R.; Ahluwalia, M.S.; Nayak, L.; Peters, S.; Arvold, N.D.; Harsh, G.R.; Steeg, P.S.; et al. Brain metastases. Nat. Rev. Dis. Prim. 2019, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Komorowski, A.S.; Warner, E.; MacKay, H.J.; Sahgal, A.; Pritchard, K.I.; Jerzak, K.J. Incidence of Brain Metastases in Nonmetastatic and Metastatic Breast Cancer: Is There a Role for Screening? Clin. Breast Cancer 2020, 20, e54–e64. [Google Scholar] [CrossRef]
- Franchino, F.; Rudà, R.; Soffietti, R. Mechanisms and Therapy for Cancer Metastasis to the Brain. Front. Oncol. 2018, 8, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Perez, C.; Planas-Rigol, E.; Trincado, J.L.; Bonfill-Teixidor, E.; Arias, A.; Marchese, D.; Moutinho, C.; Serna, G.; Pedrosa, L.; Iurlaro, R.; et al. Immune cell profiling of the cerebrospinal fluid enables the characterization of the brain metastasis microenvironment. Nat. Commun. 2021, 12, 1503. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ricarte, F.; Mayor, R.; Martínez-Sáez, E.; Rubio-Pérez, C.; Pineda, E.; Cordero, E.; Cicuéndez, M.; Poca, M.A.; López-Bigas, N.; Cajal, S.R.Y.; et al. Molecular Diagnosis of Diffuse Gliomas through Sequencing of Cell-Free Circulating Tumor DNA from Cerebrospinal Fluid. Clin. Cancer Res. 2018, 24, 2812–2819. [Google Scholar] [CrossRef] [Green Version]
- Münsterberg, J.; Loreth, D.; Brylka, L.; Werner, S.; Karbanova, J.; Gandrass, M.; Schneegans, S.; Besler, K.; Hamester, F.; Robador, J.R.; et al. ALCAM contributes to brain metastasis formation in non-small-cell lung cancer through interaction with the vascular endothelium. Neuro Oncol. 2020, 22, 955–966. [Google Scholar] [CrossRef]
- Moritz, M.N.O.; Merkel, A.R.; Feldman, E.G.; Selistre-de-Araujo, H.S.; Rhoades Sterling, J.A. Biphasic α2β1 Integrin Expression in Breast Cancer Metastasis to Bone. Int. J. Mol. Sci. 2020, 22, 6906. [Google Scholar] [CrossRef]
- Ramirez, N.E.; Zhang, Z.; Madamanchi, A.; Boyd, K.L.; O’Rear, L.D.; Nashabi, A.; Li, Z.; Dupont, W.D.; Zijlstra, A.; Zutter, M.M. The α₂β₁ integrin is a metastasis suppressor in mouse models and human cancer. J. Clin. Investig. 2011, 121, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Wang, Y.; Zhang, Z.; Qin, R.; Peng, Y.; Tang, W.; Xi, Y.; Tian, G.; Zhang, Y. Roles of intercellular cell adhesion molecule-1 (ICAM-1) in colorectal cancer: Expression, functions, prognosis, tumorigenesis, polymorphisms and therapeutic implications. Front. Oncol. 2022, 12, 1052672. [Google Scholar] [CrossRef]
- Şener, B.B.; Yiğit, D.; Bayraç, A.T.; Bayraç, C. Inhibition of cell migration and invasion by ICAM-1 binding DNA aptamers. Anal. Biochem. 2021, 628, 114262. [Google Scholar] [CrossRef]
- Ghislin, S.; Obino, D.; Middendorp, S.; Boggetto, N.; Alcaide-Loridan, C.; Deshayes, F. LFA-1 and ICAM-1 expression induced during melanoma-endothelial cell co-culture favors the transendothelial migration of melanoma cell lines in vitro. BMC Cancer 2012, 12, 455. [Google Scholar] [CrossRef]
- Khan, I.; Steeg, P.S. Keeping brain metastases dormant. Nat. Cancer 2022, 3, 3–5. [Google Scholar] [CrossRef]
- Wu, D.; Deng, S.; Li, L.; Liu, T.; Zhang, T.; Li, J.; Yu, Y.; Xu, Y. TGF-β1-mediated exosomal lnc-MMP2-2 increases blood-brain barrier permeability via the miRNA-1207-5p/EPB41L5 axis to promote non-small cell lung cancer brain metastasis. Cell Death Dis. 2021, 12, 721. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, J.; Zhu, H.; Min, J.; Feng, Y.; Zhang, H. Biological characteristics of a specific brain metastatic cell line derived from human lung adenocarcinoma. Med. Oncol. 2010, 27, 708–714. [Google Scholar] [CrossRef]
- Fazakas, C.; Wilhelm, I.; Nagyoszi, P.; Farkas, A.E.; Haskó, J.; Molnár, J.; Bauer, H.; Bauer, H.C.; Ayaydin, F.; Dung, N.T.; et al. Transmigration of melanoma cells through the blood-brain barrier: Role of endothelial tight junctions and melanoma-released serine proteases. PLoS ONE 2011, 6, e20758. [Google Scholar] [CrossRef] [Green Version]
- Wrobel, J.K.; Toborek, M. Blood-brain Barrier Remodeling during Brain Metastasis Formation. Mol. Med. 2016, 22, 32–40. [Google Scholar] [CrossRef]
- András, I.E.; Toborek, M. Extracellular vesicles of the blood-brain barrier. Tissue Barriers 2016, 4, e1131804. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, M.T.; Leaver, H.A. Brain endothelial cell death: Modes, signaling pathways, and relevance to neural development, homeostasis, and disease. Mol. Neurobiol. 2010, 42, 52–63. [Google Scholar] [CrossRef]
- Wosik, K.; Biernacki, K.; Khouzam, M.P.; Prat, A. Death receptor expression and function at the human blood brain barrier. J. Neurol. Sci. 2007, 259, 53–60. [Google Scholar] [CrossRef]
- Tămaș, F.; Bălașa, R.; Manu, D.; Gyorki, G.; Chinezu, R.; Tămaș, C.; Bălașa, A. The Importance of Small Extracellular Vesicles in the Cerebral Metastatic Process. Int. J. Mol. Sci. 2022, 23, 1449. [Google Scholar] [CrossRef]
- Teuwen, L.A.; Geldhof, V.; Pasut, A.; Carmeliet, P. COVID-19: The vasculature unleashed. Nat. Rev. Immunol. 2020, 20, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, H.; Tachikawa, M.; Yagi, Y.; Umetsu, M.; Nurdin, A.; Miyauchi, E.; Watanabe, M.; Uchida, Y.; Terasaki, T. Cluster of Differentiation 46 Is the Major Receptor in Human Blood-Brain Barrier Endothelial Cells for Uptake of Exosomes Derived from Brain-Metastatic Melanoma Cells (SK-Mel-28). Mol. Pharm. 2019, 16, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, K.; Massagué, J. Targeting metastatic cancer. Nat. Med. 2021, 27, 34–44. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EMMPRIN | ICAM 2 | Beta 3 Integrin | VCAM | Alpha 2 Integrin | ICAM 1 | CCR7 | NCAM | |
|---|---|---|---|---|---|---|---|---|
| A549 | + | N.T. | N.T. | + | N.T. | + | + | + |
| MCF7 | + | − | + | + | + | + | + | + |
| MDAMB 453 | + | − | + | − | + | + | − | − |
| MDAMB 231 | + | − | N.T. | − | N.T. | − | − | − |
| H460 | + | + | N.T. | + | N.T. | + | + | − |
| SKBR3 | + | N.T. | N.T. | + | N.T. | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasco, C.; Rizzo, A.; Cordiglieri, C.; Corsini, E.; Maderna, E.; Ciusani, E.; Salmaggi, A. The Role of Adhesion Molecules and Extracellular Vesicles in an In Vitro Model of the Blood–Brain Barrier for Metastatic Disease. Cancers 2023, 15, 3045. https://doi.org/10.3390/cancers15113045
Vasco C, Rizzo A, Cordiglieri C, Corsini E, Maderna E, Ciusani E, Salmaggi A. The Role of Adhesion Molecules and Extracellular Vesicles in an In Vitro Model of the Blood–Brain Barrier for Metastatic Disease. Cancers. 2023; 15(11):3045. https://doi.org/10.3390/cancers15113045
Chicago/Turabian StyleVasco, Chiara, Ambra Rizzo, Chiara Cordiglieri, Elena Corsini, Emanuela Maderna, Emilio Ciusani, and Andrea Salmaggi. 2023. "The Role of Adhesion Molecules and Extracellular Vesicles in an In Vitro Model of the Blood–Brain Barrier for Metastatic Disease" Cancers 15, no. 11: 3045. https://doi.org/10.3390/cancers15113045
APA StyleVasco, C., Rizzo, A., Cordiglieri, C., Corsini, E., Maderna, E., Ciusani, E., & Salmaggi, A. (2023). The Role of Adhesion Molecules and Extracellular Vesicles in an In Vitro Model of the Blood–Brain Barrier for Metastatic Disease. Cancers, 15(11), 3045. https://doi.org/10.3390/cancers15113045