

Turning Tertiary Lymphoid Structures (TLS) into Hot Spots: Values of TLS in Gastrointestinal Tumors
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
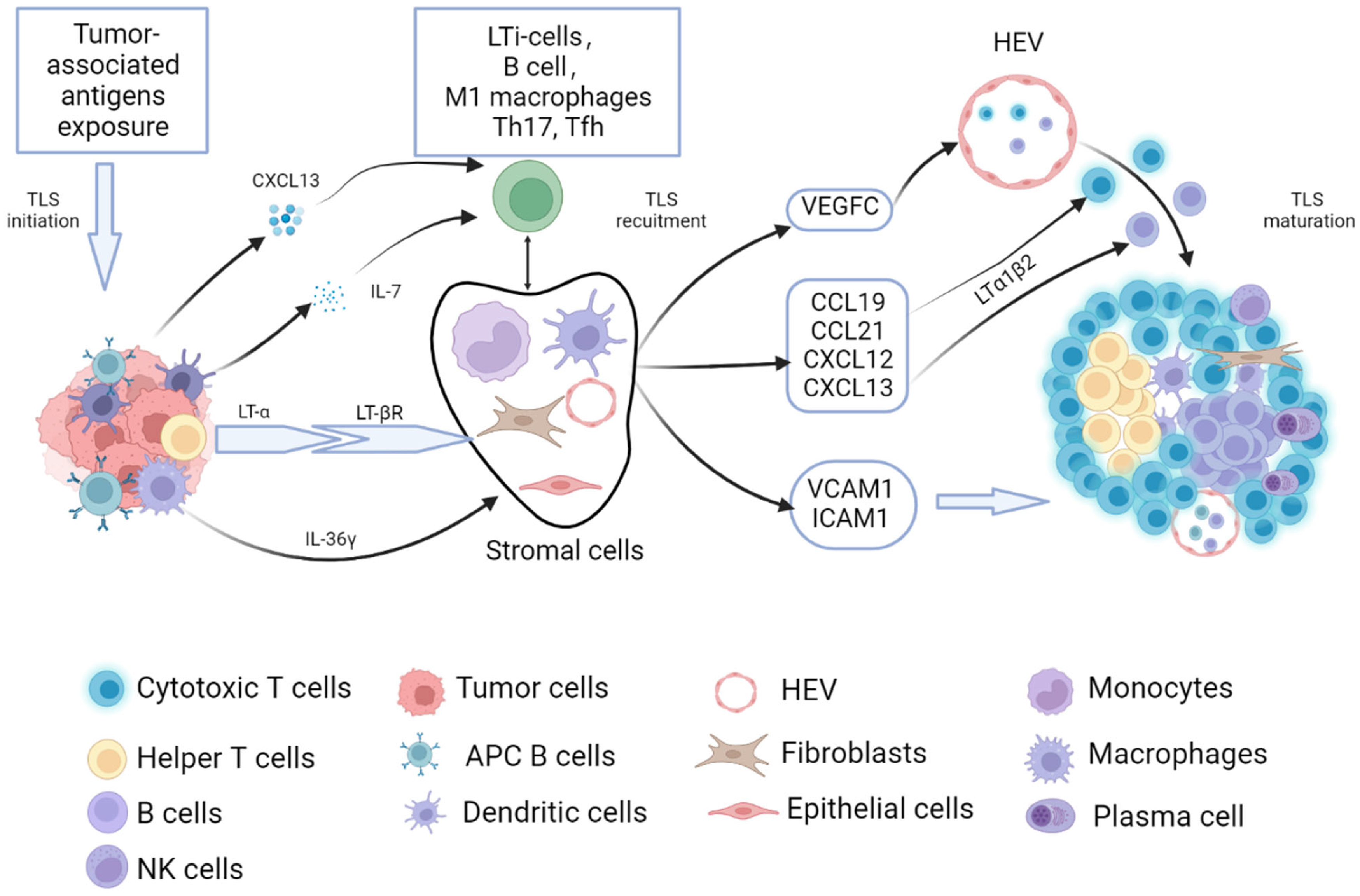
2. Process of Tumor-Related TLSs Formation and Maturation
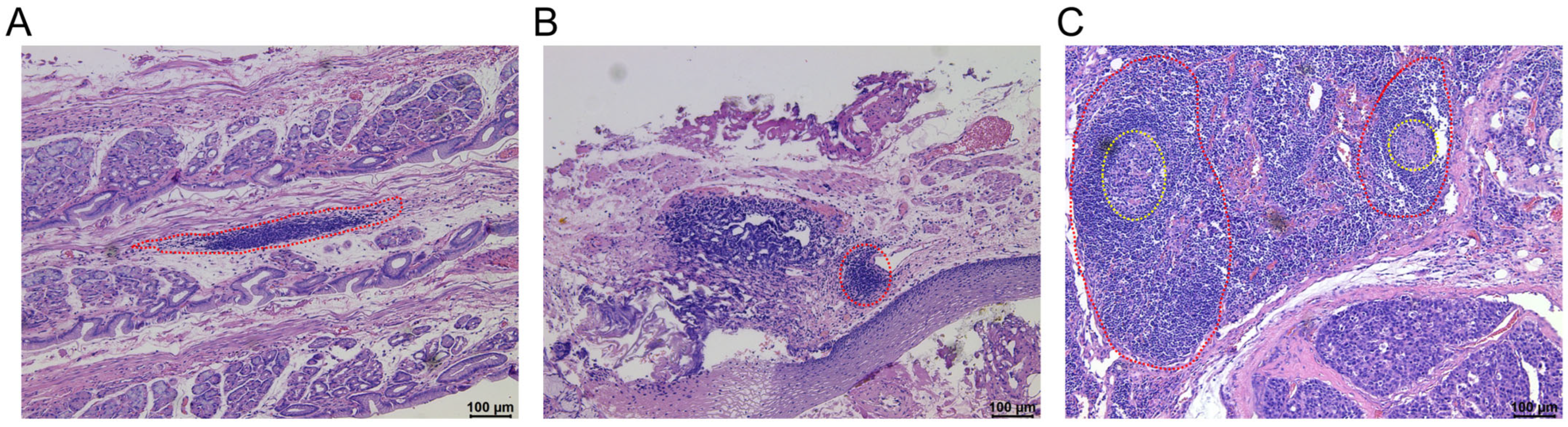
3. Recognition of the Characteristics of Tumor-Related TLSs in Gastrointestinal Tumors
4. Anti-Tumor Mechanisms and Influencing Factors of the Tumor-Associated TLSs
4.1. Anti-Tumor Mechanisms of the Tumor-Associated TLSs
4.2. Influencing Factors of the Tumor-Associated TLSs
5. Prognostic and Predictive Potential of TLSs in Gastrointestinal Tumors
5.1. Relationship between Cellular Components of TLSs and Prognosis
5.2. Relationship between the Spatial Distribution of TLSs and the Prognosis
5.3. Prognostic Role of TLSs in Combination with Other Indicators
5.4. Prediction Values of TLSs in Immune Therapy
6. Therapeutic Strategies for Inducing TLS Formation in Gastrointestinal Tumors
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dieu-Nosjean, M.C.; Goc, J.; Giraldo, N.A.; Sautes-Fridman, C.; Fridman, W.H. Tertiary lymphoid structures in cancer and beyond. Trends Immunol. 2014, 35, 571–580. [Google Scholar] [CrossRef]
- Kratz, A.; Campos-Neto, A.; Hanson, M.S.; Ruddle, N.H. Chronic inflammation caused by lymphotoxin is lymphoid neogenesis. J. Exp. Med. 1996, 183, 1461–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pipi, E.; Nayar, S.; Gardner, D.H.; Colafrancesco, S.; Smith, C.; Barone, F. Tertiary Lymphoid Structures: Autoimmunity Goes Local. Front. Immunol. 2018, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Aloisi, F.; Pujol-Borrell, R. Lymphoid neogenesis in chronic inflammatory diseases. Nat. Rev. Immunol. 2006, 6, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Nasr, I.W.; Reel, M.; Oberbarnscheidt, M.H.; Mounzer, R.H.; Baddoura, F.K.; Ruddle, N.H.; Lakkis, F.G. Tertiary lymphoid tissues generate effector and memory T cells that lead to allograft rejection. Am. J. Transplant. 2007, 7, 1071–1079. [Google Scholar] [CrossRef]
- Dieu-Nosjean, M.C.; Giraldo, N.A.; Kaplon, H.; Germain, C.; Fridman, W.H.; Sautes-Fridman, C. Tertiary lymphoid structures, drivers of the anti-tumor responses in human cancers. Immunol. Rev. 2016, 271, 260–275. [Google Scholar] [CrossRef]
- Sautes-Fridman, C.; Lawand, M.; Giraldo, N.A.; Kaplon, H.; Germain, C.; Fridman, W.H.; Dieu-Nosjean, M.C. Tertiary Lymphoid Structures in Cancers: Prognostic Value, Regulation, and Manipulation for Therapeutic Intervention. Front. Immunol. 2016, 7, 407. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global Burden of 5 Major Types of Gastrointestinal Cancer. Gastroenterology 2020, 159, 335–349. [Google Scholar] [CrossRef]
- Hennequin, A.; Derangere, V.; Boidot, R.; Apetoh, L.; Vincent, J.; Orry, D.; Fraisse, J.; Causeret, S.; Martin, F.; Arnould, L.; et al. Tumor infiltration by Tbet+ effector T cells and CD20+ B cells is associated with survival in gastric cancer patients. Oncoimmunology 2016, 5, e1054598. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Ito, M.; Ohmura, H.; Hanamura, F.; Nakano, M.; Tsuchihashi, K.; Nagai, S.; Ariyama, H.; Kusaba, H.; Yamamoto, H.; et al. Helper T cell-dominant tertiary lymphoid structures are associated with disease relapse of advanced colorectal cancer. Oncoimmunology 2020, 9, 1724763. [Google Scholar] [CrossRef]
- Lin, Q.; Tao, P.; Wang, J.; Ma, L.; Jiang, Q.; Li, J.; Zhang, G.; Liu, J.; Zhang, Y.; Hou, Y.; et al. Tumor-associated tertiary lymphoid structure predicts postoperative outcomes in patients with primary gastrointestinal stromal tumors. Oncoimmunology 2020, 9, 1747339. [Google Scholar] [CrossRef] [Green Version]
- Sautes-Fridman, C.; Petitprez, F.; Calderaro, J.; Fridman, W.H. Tertiary lymphoid structures in the era of cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 307–325. [Google Scholar] [CrossRef] [PubMed]
- van de Pavert, S.A.; Mebius, R.E. New insights into the development of lymphoid tissues. Nat. Rev. Immunol. 2010, 10, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Denton, A.E.; Innocentin, S.; Carr, E.J.; Bradford, B.M.; Lafouresse, F.; Mabbott, N.A.; Morbe, U.; Ludewig, B.; Groom, J.R.; Good-Jacobson, K.L.; et al. Type I interferon induces CXCL13 to support ectopic germinal center formation. J. Exp. Med. 2019, 216, 621–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtado, G.C.; Marinkovic, T.; Martin, A.P.; Garin, A.; Hoch, B.; Hubner, W.; Chen, B.K.; Genden, E.; Skobe, M.; Lira, S.A. Lymphotoxin beta receptor signaling is required for inflammatory lymphangiogenesis in the thyroid. Proc. Natl. Acad. Sci. USA 2007, 104, 5026–5031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overacre-Delgoffe, A.E.; Bumgarner, H.J.; Cillo, A.R.; Burr, A.H.P.; Tometich, J.T.; Bhattacharjee, A.; Bruno, T.C.; Vignali, D.A.A.; Hand, T.W. Microbiota-specific T follicular helper cells drive tertiary lymphoid structures and anti-tumor immunity against colorectal cancer. Immunity 2021, 54, 2812–2824. [Google Scholar] [CrossRef]
- Luther, S.A.; Bidgol, A.; Hargreaves, D.C.; Schmidt, A.; Xu, Y.; Paniyadi, J.; Matloubian, M.; Cyster, J.G. Differing activities of homeostatic chemokines CCL19, CCL21, and CXCL12 in lymphocyte and dendritic cell recruitment and lymphoid neogenesis. J. Immunol. 2002, 169, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Fleige, H.; Ravens, S.; Moschovakis, G.L.; Bolter, J.; Willenzon, S.; Sutter, G.; Haussler, S.; Kalinke, U.; Prinz, I.; Forster, R. IL-17-induced CXCL12 recruits B cells and induces follicle formation in BALT in the absence of differentiated FDCs. J. Exp. Med. 2014, 211, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Feng, Z.; Luo, J.; He, Z.; Liu, J.; Wu, J.; Rong, P. Tertiary Lymphoid Structures in Cancer: The Double-Edged Sword Role in Antitumor Immunity and Potential Therapeutic Induction Strategies. Front. Immunol. 2021, 12, 689270. [Google Scholar] [CrossRef]
- Bridgewood, C.; Stacey, M.; Alase, A.; Lagos, D.; Graham, A.; Wittmann, M. IL-36gamma has proinflammatory effects on human endothelial cells. Exp. Dermatol. 2017, 26, 402–408. [Google Scholar] [CrossRef]
- Weinstein, A.M.; Giraldo, N.A.; Petitprez, F.; Julie, C.; Lacroix, L.; Peschaud, F.; Emile, J.F.; Marisa, L.; Fridman, W.H.; Storkus, W.J.; et al. Association of IL-36gamma with tertiary lymphoid structures and inflammatory immune infiltrates in human colorectal cancer. Cancer Immunol. Immunother. 2019, 68, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Spratt, J.S., Jr.; Spjut, H.J. Prevalence and prognosis of individual clinical and pathologic variables associated with colorectal carcinoma. Cancer 1967, 20, 1976–1985. [Google Scholar] [CrossRef] [PubMed]
- Jass, J.R.; Love, S.B.; Northover, J.M. A new prognostic classification of rectal cancer. Lancet 1987, 1, 1303–1306. [Google Scholar] [CrossRef]
- Picker, L.J.; Butcher, E.C. Physiological and molecular mechanisms of lymphocyte homing. Annu. Rev. Immunol. 1992, 10, 561–591. [Google Scholar] [CrossRef] [PubMed]
- Ladanyi, A.; Kiss, J.; Somlai, B.; Gilde, K.; Fejos, Z.; Mohos, A.; Gaudi, I.; Timar, J. Density of DC-LAMP(+) mature dendritic cells in combination with activated T lymphocytes infiltrating primary cutaneous melanoma is a strong independent prognostic factor. Cancer Immunol. Immunother. 2007, 56, 1459–1469. [Google Scholar] [CrossRef] [PubMed]
- Dieu-Nosjean, M.C.; Antoine, M.; Danel, C.; Heudes, D.; Wislez, M.; Poulot, V.; Rabbe, N.; Laurans, L.; Tartour, E.; de Chaisemartin, L.; et al. Long-term survival for patients with non-small-cell lung cancer with intratumoral lymphoid structures. J. Clin. Oncol. 2008, 26, 4410–4417. [Google Scholar] [CrossRef] [Green Version]
- Finkin, S.; Yuan, D.; Stein, I.; Taniguchi, K.; Weber, A.; Unger, K.; Browning, J.L.; Goossens, N.; Nakagawa, S.; Gunasekaran, G.; et al. Ectopic lymphoid structures function as microniches for tumor progenitor cells in hepatocellular carcinoma. Nat. Immunol. 2015, 16, 1235–1244. [Google Scholar] [CrossRef]
- Jiang, Q.; Tian, C.; Wu, H.; Min, L.; Chen, H.; Chen, L.; Liu, F.; Sun, Y. Tertiary lymphoid structure patterns predicted anti-PD1 therapeutic responses in gastric cancer. Chin. J. Cancer Res. 2022, 34, 365–382. [Google Scholar] [CrossRef]
- Wang, Q.; Shen, X.; An, R.; Bai, J.; Dong, J.; Cai, H.; Zhu, H.; Zhong, W.; Chen, W.; Liu, A.; et al. Peritumoral tertiary lymphoid structure and tumor stroma percentage predict the prognosis of patients with non-metastatic colorectal cancer. Front. Immunol. 2022, 13, 962056. [Google Scholar] [CrossRef]
- Galon, J.; Costes, A.; Sanchez-Cabo, F.; Kirilovsky, A.; Mlecnik, B.; Lagorce-Pages, C.; Tosolini, M.; Camus, M.; Berger, A.; Wind, P.; et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 2006, 313, 1960–1964. [Google Scholar] [CrossRef]
- Zhang, W.H.; Wang, W.Q.; Han, X.; Gao, H.L.; Xu, S.S.; Li, S.; Li, T.J.; Xu, H.X.; Li, H.; Ye, L.Y.; et al. Infiltrating pattern and prognostic value of tertiary lymphoid structures in resected non-functional pancreatic neuroendocrine tumors. J. Immunother. Cancer 2020, 8, e001188. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Li, P.; Cheng, H.; Zhao, X.; Dong, M.; Zhang, Y.; Zhao, P.; Chen, J.; Shao, C. Prognostic Value of Tumor-Infiltrating Lymphocytes and Tertiary Lymphoid Structures in Epstein-Barr Virus-Associated and -Negative Gastric Carcinoma. Front. Immunol. 2021, 12, 692859. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, D.; He, W.; Chen, T.; Yan, Z.; Gao, X.; Chen, L.; Zheng, X.; Xu, B.; Lu, B.; et al. CD8(+) T cells located in tertiary lymphoid structures are associated with improved prognosis in patients with gastric cancer. Oncol. Lett. 2020, 20, 2655–2664. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Kim, Y.M.; Kim, S.; Lee, W.S.; Kong, S.J.; Yang, H.; Kang, B.; Cheon, J.; Shin, S.J.; Kim, C.; et al. High endothelial venule is a surrogate biomarker for T-cell inflamed tumor microenvironment and prognosis in gastric cancer. J. Immunother. Cancer 2021, 9, e003353. [Google Scholar] [CrossRef] [PubMed]
- Meylan, M.; Petitprez, F.; Becht, E.; Bougouin, A.; Pupier, G.; Calvez, A.; Giglioli, I.; Verkarre, V.; Lacroix, G.; Verneau, J.; et al. Tertiary lymphoid structures generate and propagate anti-tumor antibody-producing plasma cells in renal cell cancer. Immunity 2022, 55, 527–541 e525. [Google Scholar] [CrossRef] [PubMed]
- Sakimura, C.; Tanaka, H.; Okuno, T.; Hiramatsu, S.; Muguruma, K.; Hirakawa, K.; Wanibuchi, H.; Ohira, M. B cells in tertiary lymphoid structures are associated with favorable prognosis in gastric cancer. J. Surg. Res. 2017, 215, 74–82. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Chandel, N.S. Cancer metabolism: Looking forward. Nat. Rev. Cancer 2021, 21, 669–680. [Google Scholar] [CrossRef]
- Yamakoshi, Y.; Tanaka, H.; Sakimura, C.; Deguchi, S.; Mori, T.; Tamura, T.; Toyokawa, T.; Muguruma, K.; Hirakawa, K.; Ohira, M. Immunological potential of tertiary lymphoid structures surrounding the primary tumor in gastric cancer. Int. J. Oncol. 2020, 57, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Buisseret, L.; Desmedt, C.; Garaud, S.; Fornili, M.; Wang, X.; Van den Eyden, G.; de Wind, A.; Duquenne, S.; Boisson, A.; Naveaux, C.; et al. Reliability of tumor-infiltrating lymphocyte and tertiary lymphoid structure assessment in human breast cancer. Mod. Pathol. 2017, 30, 1204–1212. [Google Scholar] [CrossRef] [Green Version]
- Coppola, D.; Nebozhyn, M.; Khalil, F.; Dai, H.; Yeatman, T.; Loboda, A.; Mule, J.J. Unique ectopic lymph node-like structures present in human primary colorectal carcinoma are identified by immune gene array profiling. Am. J. Pathol. 2011, 179, 37–45. [Google Scholar] [CrossRef]
- Jia, L.; Wang, T.; Zhao, Y.; Zhang, S.; Ba, T.; Kuai, X.; Wang, B.; Zhang, N.; Zhao, W.; Yang, Z.; et al. Single-cell profiling of infiltrating B cells and tertiary lymphoid structures in the TME of gastric adenocarcinomas. Oncoimmunology 2021, 10, 1969767. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, T.N.; Thommen, D.S. Tertiary lymphoid structures in cancer. Science 2022, 375, eabf9419. [Google Scholar] [CrossRef] [PubMed]
- Motallebzadeh, R.; Bolton, E.M.; Pettigrew, G.J. Lymphoid tissue formation in allografts: Innocent until proven guilty. Transplantation 2008, 85, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Drayton, D.L.; Liao, S.; Mounzer, R.H.; Ruddle, N.H. Lymphoid organ development: From ontogeny to neogenesis. Nat. Immunol. 2006, 7, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, T.; Berghoff, A.S.; Glogner, C.; Glueck, O.; Rajky, O.; Traxler, D.; Birner, P.; Preusser, M.; Klepetko, W.; Hoetzenecker, K. Tumor-infiltrating lymphocyte subsets and tertiary lymphoid structures in pulmonary metastases from colorectal cancer. Clin. Exp. Metastasis 2016, 33, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ma, Y.; Xu, Y. Follicular regulatory T cells infiltrated the ovarian carcinoma and resulted in CD8 T cell dysfunction dependent on IL-10 pathway. Int. Immunopharmacol. 2019, 68, 81–87. [Google Scholar] [CrossRef]
- Bento, D.C.; Jones, E.; Junaid, S.; Tull, J.; Williams, G.T.; Godkin, A.; Ager, A.; Gallimore, A. High endothelial venules are rare in colorectal cancers but accumulate in extra-tumoral areas with disease progression. Oncoimmunology 2015, 4, e974374. [Google Scholar] [CrossRef]
- Chen, L.; Oke, T.; Siegel, N.; Cojocaru, G.; Tam, A.J.; Blosser, R.L.; Swailes, J.; Ligon, J.A.; Lebid, A.; Morris, C.; et al. The Immunosuppressive Niche of Soft-Tissue Sarcomas is Sustained by Tumor-Associated Macrophages and Characterized by Intratumoral Tertiary Lymphoid Structures. Clin. Cancer Res. 2020, 26, 4018–4030. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wu, J.; Xu, X.; Wang, H.; Zhang, C.; Yin, S.; He, Y. Peritumoral TIGIT(+)CD20(+) B cell infiltration indicates poor prognosis but favorable adjuvant chemotherapeutic response in gastric cancer. Int. Immunopharmacol. 2022, 108, 108735. [Google Scholar] [CrossRef]
- Brand, A.; Singer, K.; Koehl, G.E.; Kolitzus, M.; Schoenhammer, G.; Thiel, A.; Matos, C.; Bruss, C.; Klobuch, S.; Peter, K.; et al. LDHA-Associated Lactic Acid Production Blunts Tumor Immunosurveillance by T and NK Cells. Cell Metab. 2016, 24, 657–671. [Google Scholar] [CrossRef]
- Fischer, K.; Hoffmann, P.; Voelkl, S.; Meidenbauer, N.; Ammer, J.; Edinger, M.; Gottfried, E.; Schwarz, S.; Rothe, G.; Hoves, S.; et al. Inhibitory effect of tumor cell-derived lactic acid on human T cells. Blood 2007, 109, 3812–3819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posch, F.; Silina, K.; Leibl, S.; Mundlein, A.; Moch, H.; Siebenhuner, A.; Samaras, P.; Riedl, J.; Stotz, M.; Szkandera, J.; et al. Maturation of tertiary lymphoid structures and recurrence of stage II and III colorectal cancer. Oncoimmunology 2018, 7, e1378844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.S.; Huang, W.B.; Zhang, Y.H.; Chen, J.; Li, J.; Fu, H.F.; Wei, Z.W.; He, Y.L. The association of immune cell infiltration and prognostic value of tertiary lymphoid structures in gastric cancer. Neoplasma 2022, 69, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, J.; Petitprez, F.; Becht, E.; Laurent, A.; Hirsch, T.Z.; Rousseau, B.; Luciani, A.; Amaddeo, G.; Derman, J.; Charpy, C.; et al. Intra-tumoral tertiary lymphoid structures are associated with a low risk of early recurrence of hepatocellular carcinoma. J. Hepatol. 2019, 70, 58–65. [Google Scholar] [CrossRef]
- Ding, G.Y.; Ma, J.Q.; Yun, J.P.; Chen, X.; Ling, Y.; Zhang, S.; Shi, J.Y.; Chang, Y.Q.; Ji, Y.; Wang, X.Y.; et al. Distribution and density of tertiary lymphoid structures predict clinical outcome in intrahepatic cholangiocarcinoma. J. Hepatol. 2022, 76, 608–618. [Google Scholar] [CrossRef]
- Sofopoulos, M.; Fortis, S.P.; Vaxevanis, C.K.; Sotiriadou, N.N.; Arnogiannaki, N.; Ardavanis, A.; Vlachodimitropoulos, D.; Perez, S.A.; Baxevanis, C.N. The prognostic significance of peritumoral tertiary lymphoid structures in breast cancer. Cancer Immunol. Immunother. 2019, 68, 1733–1745. [Google Scholar] [CrossRef]
- Yamakoshi, Y.; Tanaka, H.; Sakimura, C.; Mori, T.; Deguchi, S.; Yoshii, M.; Tamura, T.; Toyokawa, T.; Lee, S.; Muguruma, K.; et al. Association between the preoperative neutrophil-to-lymphocyte ratio and tertiary lymphoid structures surrounding tumor in gastric cancer. Mol. Clin. Oncol. 2021, 14, 76. [Google Scholar] [CrossRef]
- Kawazoe, A.; Yamaguchi, K.; Yasui, H.; Negoro, Y.; Azuma, M.; Amagai, K.; Hara, H.; Baba, H.; Tsuda, M.; Hosaka, H.; et al. Safety and efficacy of pembrolizumab in combination with S-1 plus oxaliplatin as a first-line treatment in patients with advanced gastric/gastroesophageal junction cancer: Cohort 1 data from the KEYNOTE-659 phase IIb study. Eur. J. Cancer 2020, 129, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Boku, N.; Ryu, M.H.; Kato, K.; Chung, H.C.; Minashi, K.; Lee, K.W.; Cho, H.; Kang, W.K.; Komatsu, Y.; Tsuda, M.; et al. Safety and efficacy of nivolumab in combination with S-1/capecitabine plus oxaliplatin in patients with previously untreated, unresectable, advanced, or recurrent gastric/gastroesophageal junction cancer: Interim results of a randomized, phase II trial (ATTRACTION-4). Ann. Oncol. 2019, 30, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Fukuoka, S.; Hara, H.; Takahashi, N.; Kojima, T.; Kawazoe, A.; Asayama, M.; Yoshii, T.; Kotani, D.; Tamura, H.; Mikamoto, Y.; et al. Regorafenib Plus Nivolumab in Patients With Advanced Gastric or Colorectal Cancer: An Open-Label, Dose-Escalation, and Dose-Expansion Phase Ib Trial (REGONIVO, EPOC1603). J. Clin. Oncol. 2020, 38, 2053–2061. [Google Scholar] [CrossRef]
- Singh, A.S.; Hecht, J.R.; Rosen, L.; Wainberg, Z.A.; Wang, X.; Douek, M.; Hagopian, A.; Andes, R.; Sauer, L.; Brackert, S.R.; et al. A Randomized Phase II Study of Nivolumab Monotherapy or Nivolumab Combined with Ipilimumab in Patients with Advanced Gastrointestinal Stromal Tumors. Clin. Cancer Res. 2022, 28, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.P.; Othus, M.; Chae, Y.K.; Giles, F.J.; Hansel, D.E.; Singh, P.P.; Fontaine, A.; Shah, M.H.; Kasi, A.; Baghdadi, T.A.; et al. A Phase II Basket Trial of Dual Anti-CTLA-4 and Anti-PD-1 Blockade in Rare Tumors (DART SWOG 1609) in Patients with Nonpancreatic Neuroendocrine Tumors. Clin. Cancer Res. 2020, 26, 2290–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabernero, J.; Cutsem, E.V.; Bang, Y.-J.; Fuchs, C.S.; Wyrwicz, L.; Lee, K.W.; Kudaba, I.; Garrido, M.; Chung, H.C.; Salguero, H.R.C.; et al. Pembrolizumab with or without chemotherapy versus chemotherapy for advanced gastric or gastroesophageal junction (G/GEJ) adenocarcinoma: The phase III KEYNOTE-062 study. J. Clin. Oncol. 2019, 37, LBA4007. [Google Scholar] [CrossRef]
- Bang, Y.J.; Kang, Y.K.; Catenacci, D.V.; Muro, K.; Fuchs, C.S.; Geva, R.; Hara, H.; Golan, T.; Garrido, M.; Jalal, S.I.; et al. Pembrolizumab alone or in combination with chemotherapy as first-line therapy for patients with advanced gastric or gastroesophageal junction adenocarcinoma: Results from the phase II nonrandomized KEYNOTE-059 study. Gastric Cancer 2019, 22, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Uram, J.N.; Wang, H.; Bartlett, B.R.; Kemberling, H.; Eyring, A.D.; Skora, A.D.; Luber, B.S.; Azad, N.S.; Laheru, D.; et al. PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. N. Engl. J. Med. 2015, 372, 2509–2520. [Google Scholar] [CrossRef] [Green Version]
- Overman, M.J.; McDermott, R.; Leach, J.L.; Lonardi, S.; Lenz, H.J.; Morse, M.A.; Desai, J.; Hill, A.; Axelson, M.; Moss, R.A.; et al. Nivolumab in patients with metastatic DNA mismatch repair-deficient or microsatellite instability-high colorectal cancer (CheckMate 142): An open-label, multicentre, phase 2 study. Lancet Oncol. 2017, 18, 1182–1191. [Google Scholar] [CrossRef]
- Catenacci, D.V.T.; Kang, Y.K.; Park, H.; Uronis, H.E.; Lee, K.W.; Ng, M.C.H.; Enzinger, P.C.; Park, S.H.; Gold, P.J.; Lacy, J.; et al. Margetuximab plus pembrolizumab in patients with previously treated, HER2-positive gastro-oesophageal adenocarcinoma (CP-MGAH22-05): A single-arm, phase 1b-2 trial. Lancet Oncol. 2020, 21, 1066–1076. [Google Scholar] [CrossRef]
- Kawazoe, A.; Fukuoka, S.; Nakamura, Y.; Kuboki, Y.; Wakabayashi, M.; Nomura, S.; Mikamoto, Y.; Shima, H.; Fujishiro, N.; Higuchi, T.; et al. Lenvatinib plus pembrolizumab in patients with advanced gastric cancer in the first-line or second-line setting (EPOC1706): An open-label, single-arm, phase 2 trial. Lancet Oncol. 2020, 21, 1057–1065. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, P.; Zhang, Y.; Li, Z.; Gong, J.; Li, J.; Li, J.; Li, Y.; Zhang, X.; Lu, Z.; et al. Efficacy, Safety, and Biomarkers of Toripalimab in Patients with Recurrent or Metastatic Neuroendocrine Neoplasms: A Multiple-Center Phase Ib Trial. Clin. Cancer Res. 2020, 26, 2337–2345. [Google Scholar] [CrossRef] [Green Version]
- Overman, M.J.; Lonardi, S.; Wong, K.Y.M.; Lenz, H.J.; Gelsomino, F.; Aglietta, M.; Morse, M.A.; Van Cutsem, E.; McDermott, R.; Hill, A.; et al. Durable Clinical Benefit With Nivolumab Plus Ipilimumab in DNA Mismatch Repair-Deficient/Microsatellite Instability-High Metastatic Colorectal Cancer. J. Clin. Oncol. 2018, 36, 773–779. [Google Scholar] [CrossRef]
- D’Angelo, S.P.; Shoushtari, A.N.; Keohan, M.L.; Dickson, M.A.; Gounder, M.M.; Chi, P.; Loo, J.K.; Gaffney, L.; Schneider, L.; Patel, Z.; et al. Combined KIT and CTLA-4 Blockade in Patients with Refractory GIST and Other Advanced Sarcomas: A Phase Ib Study of Dasatinib plus Ipilimumab. Clin. Cancer Res. 2017, 23, 2972–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solinas, C.; Garaud, S.; De Silva, P.; Boisson, A.; Van den Eynden, G.; de Wind, A.; Risso, P.; Rodrigues Vitoria, J.; Richard, F.; Migliori, E.; et al. Immune Checkpoint Molecules on Tumor-Infiltrating Lymphocytes and Their Association with Tertiary Lymphoid Structures in Human Breast Cancer. Front. Immunol. 2017, 8, 1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G.; et al. B cells and tertiary lymphoid structures promote immunotherapy response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.G.; Yu, L.; Gao, H.; Balic, J.J.; West, A.; Oshima, H.; McLeod, L.; Oshima, M.; Gallimore, A.; D’Costa, K.; et al. Hyperactive gp130/STAT3-driven gastric tumourigenesis promotes submucosal tertiary lymphoid structure development. Int. J. Cancer 2018, 143, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, A.M.; Chen, L.; Brzana, E.A.; Patil, P.R.; Taylor, J.L.; Fabian, K.L.; Wallace, C.T.; Jones, S.D.; Watkins, S.C.; Lu, B.; et al. Tbet and IL-36gamma cooperate in therapeutic DC-mediated promotion of ectopic lymphoid organogenesis in the tumor microenvironment. Oncoimmunology 2017, 6, e1322238. [Google Scholar] [CrossRef] [Green Version]
- Adachi, K.; Kano, Y.; Nagai, T.; Okuyama, N.; Sakoda, Y.; Tamada, K. IL-7 and CCL19 expression in CAR-T cells improves immune cell infiltration and CAR-T cell survival in the tumor. Nat. Biotechnol. 2018, 36, 346–351. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, H.; Zhou, Q.; Ren, X. Insights into tertiary lymphoid structures in the solid tumor microenvironment: Anti-tumor mechanism, functional regulation, and immunotherapeutic strategies. Cancer Biol. Med. 2021, 18, 981. [Google Scholar] [CrossRef]
- Cottrell, T.R.; Thompson, E.D.; Forde, P.M.; Stein, J.E.; Duffield, A.S.; Anagnostou, V.; Rekhtman, N.; Anders, R.A.; Cuda, J.D.; Illei, P.B.; et al. Pathologic features of response to neoadjuvant anti-PD-1 in resected non-small-cell lung carcinoma: A proposal for quantitative immune-related pathologic response criteria (irPRC). Ann. Oncol. 2018, 29, 1853–1860. [Google Scholar] [CrossRef]
- Maldonado, L.; Teague, J.E.; Morrow, M.P.; Jotova, I.; Wu, T.C.; Wang, C.; Desmarais, C.; Boyer, J.D.; Tycko, B.; Robins, H.S.; et al. Intramuscular therapeutic vaccination targeting HPV16 induces T cell responses that localize in mucosal lesions. Sci. Transl. Med. 2014, 6, 221ra213. [Google Scholar] [CrossRef] [Green Version]
- Lutz, E.R.; Wu, A.A.; Bigelow, E.; Sharma, R.; Mo, G.; Soares, K.; Solt, S.; Dorman, A.; Wamwea, A.; Yager, A.; et al. Immunotherapy converts nonimmunogenic pancreatic tumors into immunogenic foci of immune regulation. Cancer Immunol. Res. 2014, 2, 616–631. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Watanabe, T. Gel-Trapped Lymphorganogenic Chemokines Trigger Artificial Tertiary Lymphoid Organs and Mount Adaptive Immune Responses In Vivo. Front. Immunol. 2016, 7, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morcrette, G.; Hirsch, T.Z.; Badour, E.; Pilet, J.; Caruso, S.; Calderaro, J.; Martin, Y.; Imbeaud, S.; Letouze, E.; Rebouissou, S.; et al. APC germline hepatoblastomas demonstrate cisplatin-induced intratumor tertiary lymphoid structures. Oncoimmunology 2019, 8, e1583547. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Characteristic Detecting Markers | Subpopulation Markers | Prognoses | Reference | ||
|---|---|---|---|---|---|
| Gastric cancer | B cell zone | CD20+ B cells | Favorable | [9,32,33,35,36,37] | |
| GC B cells (Bcl-6+ CD20+) | Favorable | [35] | |||
| FDCs (CD21+) | No evaluation | [32,33,35] | |||
| Macrophage (CD68+) | Unfavorable | [37] | |||
| T cell zone | CD3+ T cells | Favorable | [32,35,37] | ||
| Help T cells (CD4+) | No evaluation | [33,35] | |||
| Th1 (CD4+ T-bet+) | Favorable | [9] | |||
| Tregs (CD4+ FOXP3+) | Unfavorable | [9] | |||
| Cytotoxic T cells (CD8+) | Favorable | [32,35,37] | |||
| Memory T cells (CD45RO+) | No evaluation | ||||
| HEVs | PNAd or MECA79 | PNAd or MECA79 | Favorable | [33,35] | |
| Colorectal cancer | B cell zone | CD20+ B cells | Tfh cells (CD3+ CD8- Bcl-6+) | No significance | [10] |
| FDCs (FDC+) | No significance | [10,29] | |||
| GC B cells (Bcl-6+ CD20+) | No significance | [10] | |||
| Macrophage (CD68+) | Unfavorable | [10,29] | |||
| T cell zone | CD3+ T cells | CTL (CD8+) | No significance | [10] | |
| Th1 (CD3+ T-bet+) | No significance | [10] | |||
| Th2 (CD3+ GATA3+) | Unfavorable | [10] | |||
| Th17(CD3+ ROR-γT+) | No significance | [10] | |||
| Tregs (CD3+ FOXP3+) | Unfavorable | [10,38] | |||
| Memory T cells (CD45RO+) | No evaluation | [29] | |||
| Other immune cells | NCR1+ NK cells | No significance | [29] | ||
| CD15+ TAN cells | No significance | [29] | |||
| HEVs | PNAd or MECA79 | PNAd or MECA79 | Unfavorable | [27] | |
| Gastrointestinal Stromal Tumors (GIST) | B cell zone | CD20+ B cells | Naive B cells (Bn) (CD20+ CD27− IgM+) | No evaluation | [11] |
| IgM+ memory B cells (IgM+ Bm) (CD20+ CD27+ IgM+) | No evaluation | [11] | |||
| CD27− isotypeswitched memory B cells (CD27− Sw Bm) (CD20+ CD27− IgM−) | No evaluation | [11] | |||
| CD27+ isotype-switched memory B cells (CD27+ Sw Bm) (CD20+ CD27+ IgM−) | No evaluation | [11] | |||
| Plasma cells (PCs) (CD20-CD24−CD27hiCD38hi) | No evaluation | [11] | |||
| T cell zone | CD3+ T cells | CTL (CD8+) | No significance | [11] | |
| Th1 (CD4+ T-bet+) | No significance | [11] | |||
| Th2 (CD4+ GATA3+) | No significance | [11] | |||
| Th17(CD4+ ROR-γT+) | No significance | [11] | |||
| Tregs: (CD4+ FOXP3+) | Unfavorable | [11] | |||
| Other immune cells | Tissue-resident memory T cells | CD103+ Trm | No evaluation | [11] | |
| HEVs | PNAd or MECA79 | PNAd or MECA79 | No detected |
| Tumor Type | Medication Plan | Key Outcome | Effectiveness | Reference | |
|---|---|---|---|---|---|
| Gastric cancer and Gastroesophageal junction cancer | GC | Pembrolizumab + lenvatinib | ORR 69% | Positive | [68] |
| GC/GEJC | Pembrolizumab + SOX | ORR 72.2% | Positive | [58] | |
| GEJC | Margetuximab + pembrolizumab | ORR 18% | No significance | [67] | |
| GC/GEJC | Pembrolizumab | ORR 25.8% | No significance | [64] | |
| GC/GEJC | Pembrolizumab | ORR 14.5% | No significance | [63] | |
| GC/GEJC | Nivolumab + CapeOX | ORR 76.5% | Positive | [59] | |
| Colorectal cancer | MSI-H mCRC | Pembrolizumab | ORR 54% | Positive | [65] |
| MSS mCRC | Pembrolizumab | ORR 0% | Failed | [65] | |
| Advanced CRC | Nivolumab | ORR 32% | No significance | [66,70] | |
| Advanced CRC | Nivolumab+ Ipilimumab | ORR 55% | Positive | [66,70] | |
| MSS CRC | Regorafenib+ Nivolumab | ORR 36% | No significance | [60] | |
| Gastrointestinal Stromal Tumors (GIST) | Advanced GIST | Nivolumab+Ipilimumab | CR 6.7% | No significance | [61] |
| Advanced GIST | Dasatinib+Ipilimumab | ORR 53.8% | Positive | [71] | |
| Gastrointestinal neuroendocrine neoplasms (GE-NEN) | Recurrent or Metastatic GNENs | Toripalimab | ORR 20% | No significance | [69] |
| NENs (GNENs47%) | Ipilimumab +nivolumab | ORR 25% | No significance | [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, D.; Yu, H.; Wang, X.; Mao, Y.; Liang, M.; Lu, X.; Shen, X.; Guan, W. Turning Tertiary Lymphoid Structures (TLS) into Hot Spots: Values of TLS in Gastrointestinal Tumors. Cancers 2023, 15, 367. https://doi.org/10.3390/cancers15020367
Cai D, Yu H, Wang X, Mao Y, Liang M, Lu X, Shen X, Guan W. Turning Tertiary Lymphoid Structures (TLS) into Hot Spots: Values of TLS in Gastrointestinal Tumors. Cancers. 2023; 15(2):367. https://doi.org/10.3390/cancers15020367
Chicago/Turabian StyleCai, Daming, Heng Yu, Xingzhou Wang, Yonghuan Mao, Mengjie Liang, Xiaofeng Lu, Xiaofei Shen, and Wenxian Guan. 2023. "Turning Tertiary Lymphoid Structures (TLS) into Hot Spots: Values of TLS in Gastrointestinal Tumors" Cancers 15, no. 2: 367. https://doi.org/10.3390/cancers15020367
APA StyleCai, D., Yu, H., Wang, X., Mao, Y., Liang, M., Lu, X., Shen, X., & Guan, W. (2023). Turning Tertiary Lymphoid Structures (TLS) into Hot Spots: Values of TLS in Gastrointestinal Tumors. Cancers, 15(2), 367. https://doi.org/10.3390/cancers15020367