
Notch Signaling Suppresses Melanoma Tumor Development in BRAF/Pten Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of Notch1- and Notch2-Null Mice
2.2. Immunohistochemistry
2.3. Immunohistochemical Analysis of TMA
2.4. Antibodies
2.5. Multiplex Chromogenic
3. Results
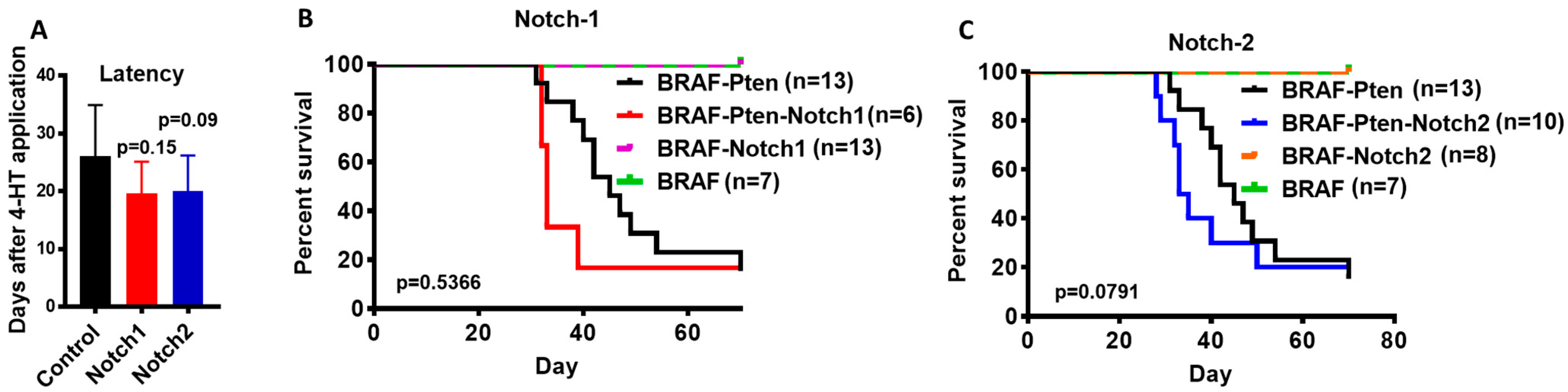
3.1. Conditional Knockout of Notch1 or Notch2 in Mouse Melanocytes Accelerates Melanoma Tumor Development and Growth
3.2. Notch1 or Notch2 Loss Cannot Substitute for Pten Loss or Cooperate with BRAFV600E in Melanomagenesis
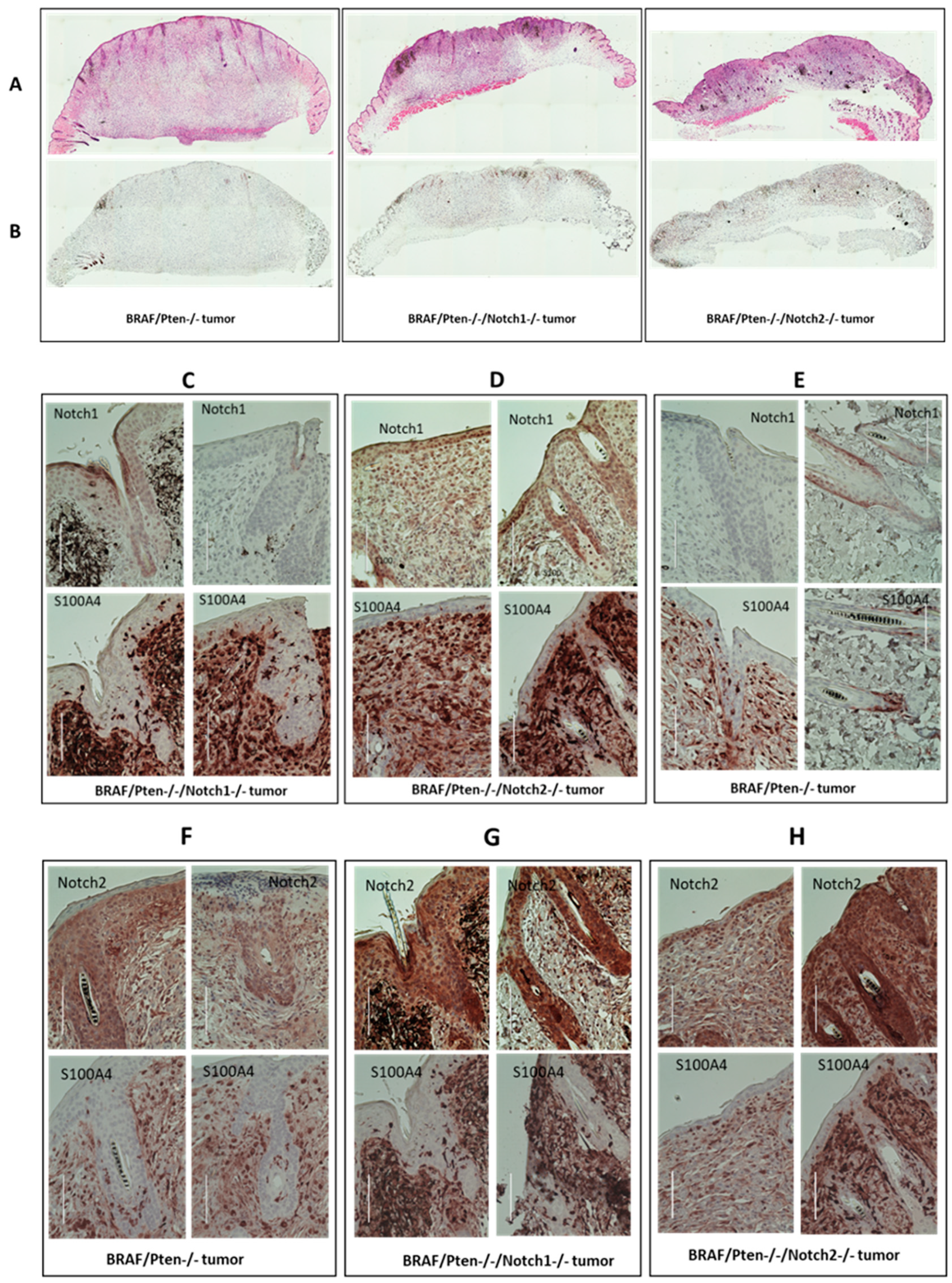
3.3. Immunohistochemical Analysis of Melanoma Tumors
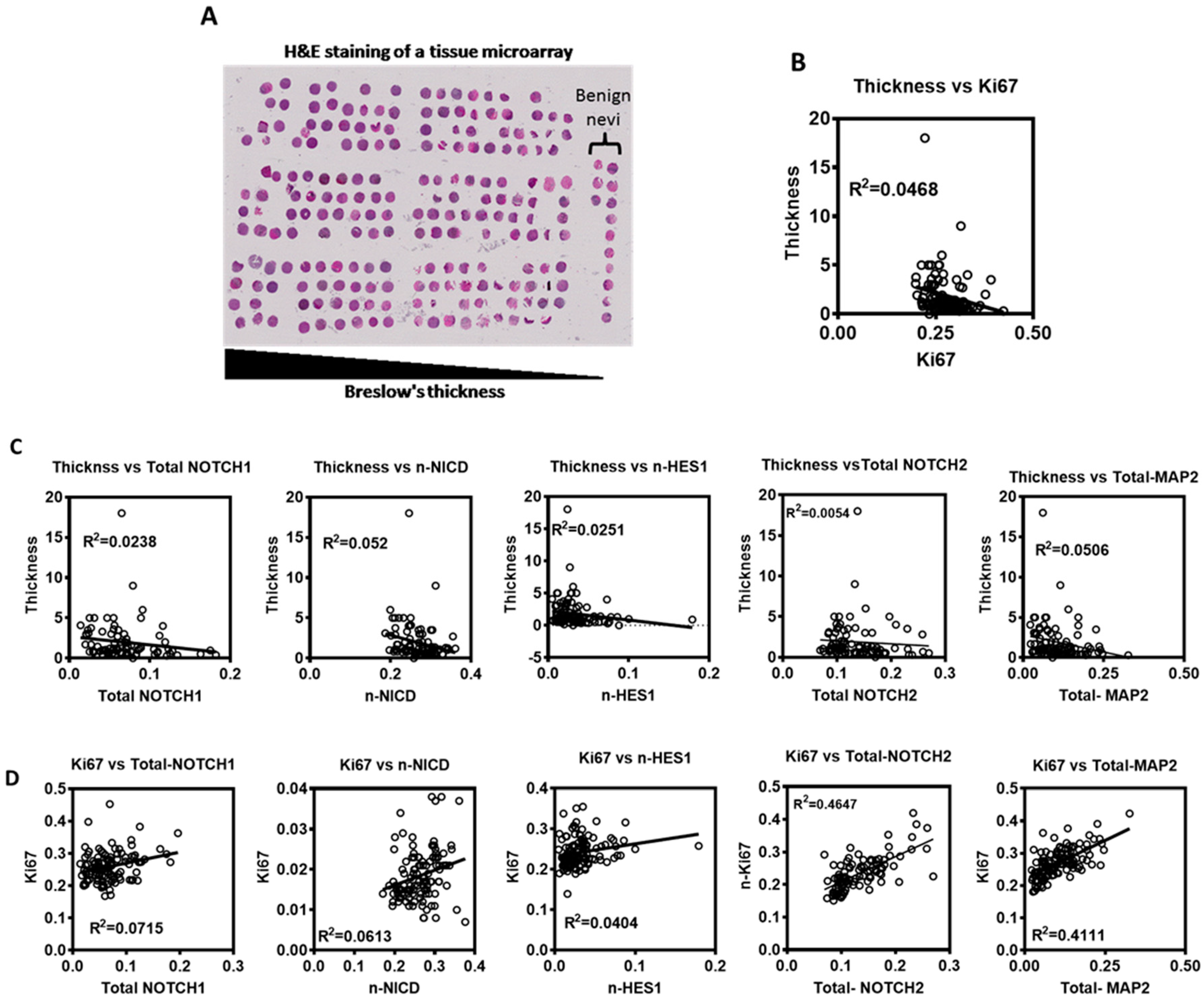
3.4. Evaluation of Notch Signaling in Human Primary Melanoma Tumors
3.5. Prognostic Value of Notch Signaling in Melanoma
4. Discussion
4.1. Notch Proteins as Melanoma Tumor Suppressors
4.2. Role of Notch1 in Melanoma Tumorigenesis and Progression
4.3. Prognostic Relevance Notch Signaling in Human Cutaneous Melanoma
4.4. Notch Signaling and Neuronal Differentiation Markers in Human Cutaneous Melanoma
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saginala, K.; Barsouk, A.; Aluru, J.S.; Rawla, P.; Barsouk, A. Epidemiology of Melanoma. Med. Sci. 2021, 9, 63. [Google Scholar] [CrossRef]
- National Cancer Institute. Cancer Stat Facts: Melanoma of The Skin. Available online: https://seer.cancer.gov/statfacts/html/melan.html (accessed on 13 December 2022).
- American Cancer Society. Key Statistics for Melanoma Skin Cancer. 2022. Available online: https://www.cancer.org/cancer/melanoma-skin-cancer/about/key-statistics.html#references (accessed on 13 December 2022).
- Liu, Z.-J.; Xiao, M.; Balint, K.; Smalley, K.S.; Brafford, P.; Qiu, R.; Pinnix, C.C.; Li, X.; Herlyn, M. Notch1 Signaling Promotes Primary Melanoma Progression by Activating Mitogen-Activated Protein Kinase/Phosphatidylinositol 3-Kinase-Akt Pathways and Up-regulating N-Cadherin Expression. Cancer Res. 2006, 66, 4182–4190. [Google Scholar] [CrossRef] [Green Version]
- Rad, E.B.; Hammerlindl, H.; Wels, C.; Popper, U.; Menon, D.R.; Breiteneder, H.; Kitzwoegerer, M.; Hafner, C.; Herlyn, M.; Bergler, H.; et al. Notch4 Signaling Induces a Mesenchymal–Epithelial–like Transition in Melanoma Cells to Suppress Malignant Behaviors. Cancer Res. 2016, 76, 1690–1697. [Google Scholar] [CrossRef] [Green Version]
- Keyghobadi, F.; Mehdipour, M.; Nekoukar, V.; Firouzi, J.; Kheimeh, A.; Lahrood, F.N.; Zavareh, V.A.; Azimi, M.; Mohammadi, M.; Sodeifi, N.; et al. Long-Term Inhibition of Notch in A-375 Melanoma Cells Enhances Tumor Growth Through the Enhancement of AXIN1, CSNK2A3, and CEBPA2 as Intermediate Genes in Wnt and Notch Pathways. Front. Oncol. 2020, 10, 531. [Google Scholar] [CrossRef]
- Wohlfeil, S.A.; Häfele, V.; Dietsch, B.; Schledzewski, K.; Winkler, M.; Zierow, J.; Leibing, T.; Mohammadi, M.M.; Heineke, J.; Sticht, C.; et al. Hepatic Endothelial Notch Activation Protects against Liver Metastasis by Regulating Endothelial-Tumor Cell Adhesion Independent of Angiocrine Signaling. Cancer Res. 2019, 79, 598–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydin, I.T.; Melamed, R.; Adams, S.J.; Castillo-Martin, M.; Demir, A.; Bryk, D.; Brunner, G.; Cordon-Cardo, C.; Osman, I.; Rabadan, R.; et al. FBXW7 Mutations in Melanoma and a New Therapeutic Paradigm. JNCI J. Natl. Cancer Inst. 2014, 106, dju107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balint, K.; Xiao, M.; Pinnix, C.C.; Soma, A.; Veres, I.; Juhasz, I.; Brown, E.J.; Capobianco, A.J.; Herlyn, M.; Liu, Z.-J. Activation of Notch1 signaling is required for β-catenin-mediated human primary melanoma progression. J. Clin. Investig. 2005, 115, 3166–3176. [Google Scholar] [CrossRef]
- Bedogni, B.; Warneke, J.A.; Nickoloff, B.J.; Giaccia, A.J.; Powell, M.B. Notch1 is an effector of Akt and hypoxia in melanoma development. J. Clin. Investig. 2008, 118, 3660–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, C.; Poliseno, L.; Segura, M.F.; Medicherla, R.; Haimovic, A.; Menendez, S.; Shang, S.; Pavlick, A.; Shao, Y.; Darvishian, F.; et al. The Novel Gamma Secretase Inhibitor RO4929097 Reduces the Tumor Initiating Potential of Melanoma. PLoS ONE 2011, 6, e25264. [Google Scholar] [CrossRef]
- Yang, Z.; Qi, Y.; Lai, N.; Zhang, J.; Chen, Z.; Liu, M.; Zhang, W.; Luo, R.; Kang, S. Notch1 signaling in melanoma cells promoted tumor-induced immunosuppression via upregulation of TGF-β1. J. Exp. Clin. Cancer Res. 2018, 37, 519. [Google Scholar] [CrossRef]
- Zhang, J.P.; Li, N.; Bai, W.Z.; Qiu, X.C.; Ma, B.A.; Zhou, Y.; Fan, Q.Y.; Shan, L.Q. Notch ligand Delta-like 1 promotes the metastasis of melanoma by enhancing tumor adhesion. Braz. J. Med. Biol. Res. 2014, 47, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, H.; Moller, M.; Cai, L.; Prokupets, R.; Yang, C.; Costa, C.; Yu, K.; Le, N.; Liu, Z.-J. Converting melanoma-associated fibroblasts into a tumor-suppressive phenotype by increasing intracellular Notch1 pathway activity. PLoS ONE 2021, 16, e0248260. [Google Scholar] [CrossRef]
- Fan, L.; Liu, H.; Zhu, G.; Singh, S.; Yu, Z.; Wang, S.; Luo, H.; Liu, S.; Xu, Y.; Ge, J.; et al. Caspase-4/11 is critical for angiogenesis by repressing Notch1 signaling via inhibiting γ-secretase activity. Br. J. Pharmacol. 2022, 179, 4809–4828. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, Q.; Duan, Q.; Tan, Y.; Sun, T.; Qi, C. NOTCH4 mutation as predictive biomarker for immunotherapy benefits in NRAS wildtype melanoma. Front. Immunol. 2022, 13, 894110. [Google Scholar] [CrossRef]
- Skarmoutsou, E.; Bevelacqua, V.; D’Amico, F.; Russo, A.; Spandidos, D.; Scalisi, A.; Malaponte, G.; Guarneri, C. FOXP3 expression is modulated by TGF-β1/NOTCH1 pathway in human melanoma. Int. J. Mol. Med. 2018, 42, 392–404. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Gao, Y.; Niu, X.; Tang, M.; Li, J.; Song, B.; Guan, X. LncRNA BASP1-AS1 interacts with YBX1 to regulate Notch transcription and drives the malignancy of melanoma. Cancer Sci. 2021, 112, 4526–4542. [Google Scholar] [CrossRef]
- Mikheil, D.M.; Prabhakar, K.; Arshad, A.; Rodriguez, C.I.; Newton, M.A.; Setaluri, V. Notch signaling activation induces cell death in MAPKi-resistant melanoma cells. Pigment. Cell Melanoma Res. 2019, 32, 528–539. [Google Scholar] [CrossRef]
- Dankort, D.; Curley, D.P.; Cartlidge, R.A.; Nelson, B.; Karnezis, A.N.; Damsky, W.E., Jr.; You, M.J.; DePinho, R.A.; McMahon, M.; Bosenberg, M. BrafV600E cooperates with Pten loss to induce metastatic melanoma. Nat. Genet. 2009, 41, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.; Mirlacher, M.; Sauter, G. Immunohistochemical Analysis of Tissue Microarrays. In Tissue Microarrays; Humana Press: Totowa, NJ, USA, 2010; Volume 664, pp. 113–126. [Google Scholar] [CrossRef]
- Hooijkaas, A.I.; Gadiot, J.; van der Valk, M.; Mooi, W.J.; Blank, C.U. Targeting BRAFV600E in an Inducible Murine Model of Melanoma. Am. J. Pathol. 2012, 181, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Villegas, S.N.; Gombos, R.; García-López, L.; Gutiérrez-Pérez, I.; García-Castillo, J.; Vallejo, D.M.; Da Ros, V.G.; Ballesta-Illán, E.; Mihály, J.; Dominguez, M. PI3K/Akt Cooperates with Oncogenic Notch by Inducing Nitric Oxide-Dependent Inflammation. Cell Rep. 2018, 22, 2541–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, M.-Y.; Yang, M.H.; Schnegg, C.I.; Hwang, S.; Ryu, B.; Alani, R.M. Notch3 signaling-mediated melanoma–endothelial crosstalk regulates melanoma stem-like cell homeostasis and niche morphogenesis. Lab. Investig. 2017, 97, 725–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltani, M.H.; Pichardo, R.; Song, Z.; Sangha, N.; Camacho, F.; Satyamoorthy, K.; Sangueza, O.P.; Setaluri, V. Microtubule-Associated Protein 2, a Marker of Neuronal Differentiation, Induces Mitotic Defects, Inhibits Growth of Melanoma Cells, and Predicts Metastatic Potential of Cutaneous Melanoma. Am. J. Pathol. 2005, 166, 1841–1850. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikheil, D.; Prabhakar, K.; Ng, T.L.; Teertam, S.; Longley, B.J.; Newton, M.A.; Setaluri, V. Notch Signaling Suppresses Melanoma Tumor Development in BRAF/Pten Mice. Cancers 2023, 15, 519. https://doi.org/10.3390/cancers15020519
Mikheil D, Prabhakar K, Ng TL, Teertam S, Longley BJ, Newton MA, Setaluri V. Notch Signaling Suppresses Melanoma Tumor Development in BRAF/Pten Mice. Cancers. 2023; 15(2):519. https://doi.org/10.3390/cancers15020519
Chicago/Turabian StyleMikheil, Dareen, Kirthana Prabhakar, Tun Lee Ng, Sireesh Teertam, B. Jack Longley, Michael A. Newton, and Vijayasaradhi Setaluri. 2023. "Notch Signaling Suppresses Melanoma Tumor Development in BRAF/Pten Mice" Cancers 15, no. 2: 519. https://doi.org/10.3390/cancers15020519
APA StyleMikheil, D., Prabhakar, K., Ng, T. L., Teertam, S., Longley, B. J., Newton, M. A., & Setaluri, V. (2023). Notch Signaling Suppresses Melanoma Tumor Development in BRAF/Pten Mice. Cancers, 15(2), 519. https://doi.org/10.3390/cancers15020519