
Cell Membrane Sialome: Sialic Acids as Therapeutic Targets and Regulators of Drug Resistance in Human Cancer Management


Abstract
:Simple Summary
Abstract
1. Introduction
2. Sialic Acids in Cancer—Smart Players in a Complex Mechanism
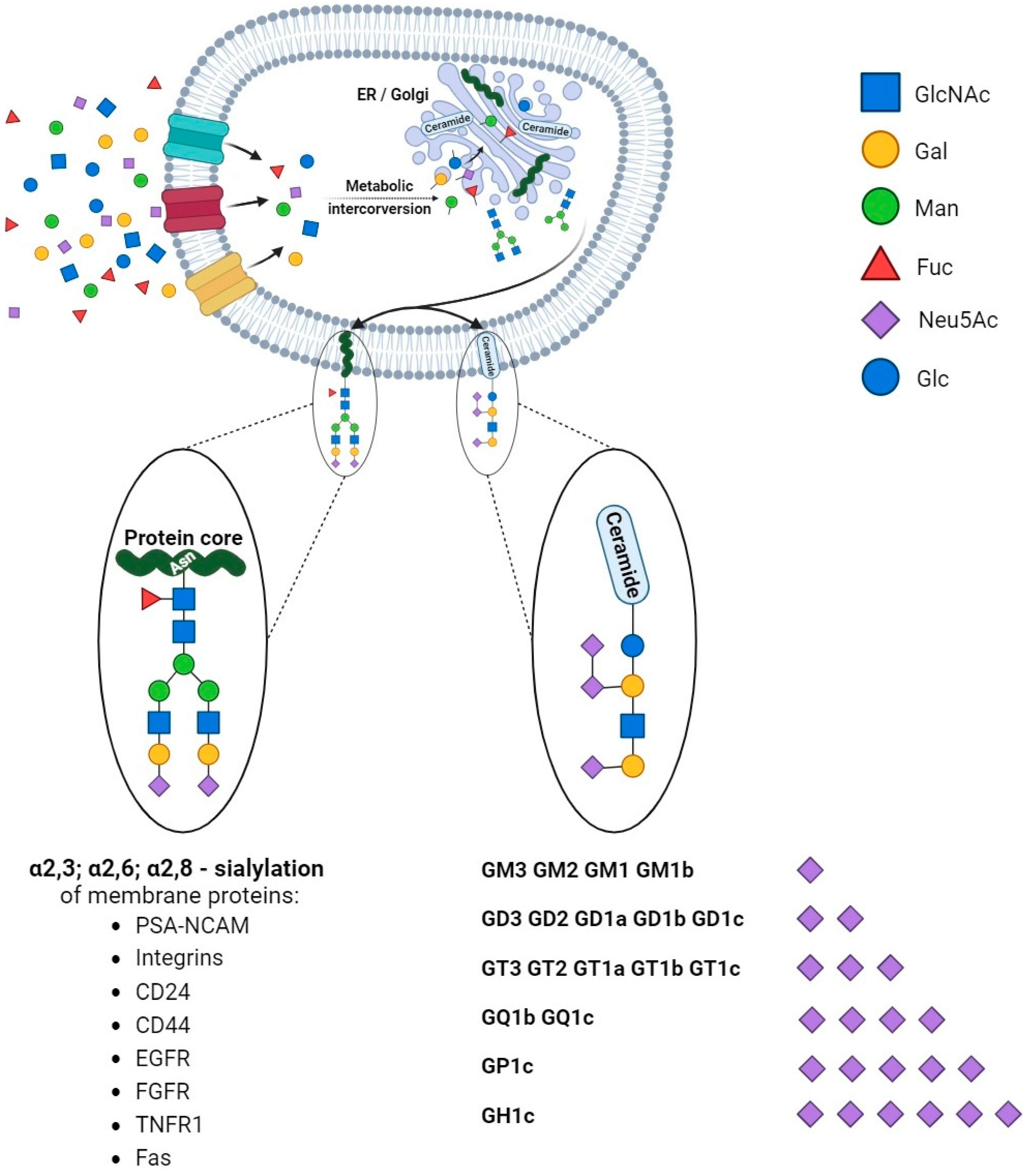
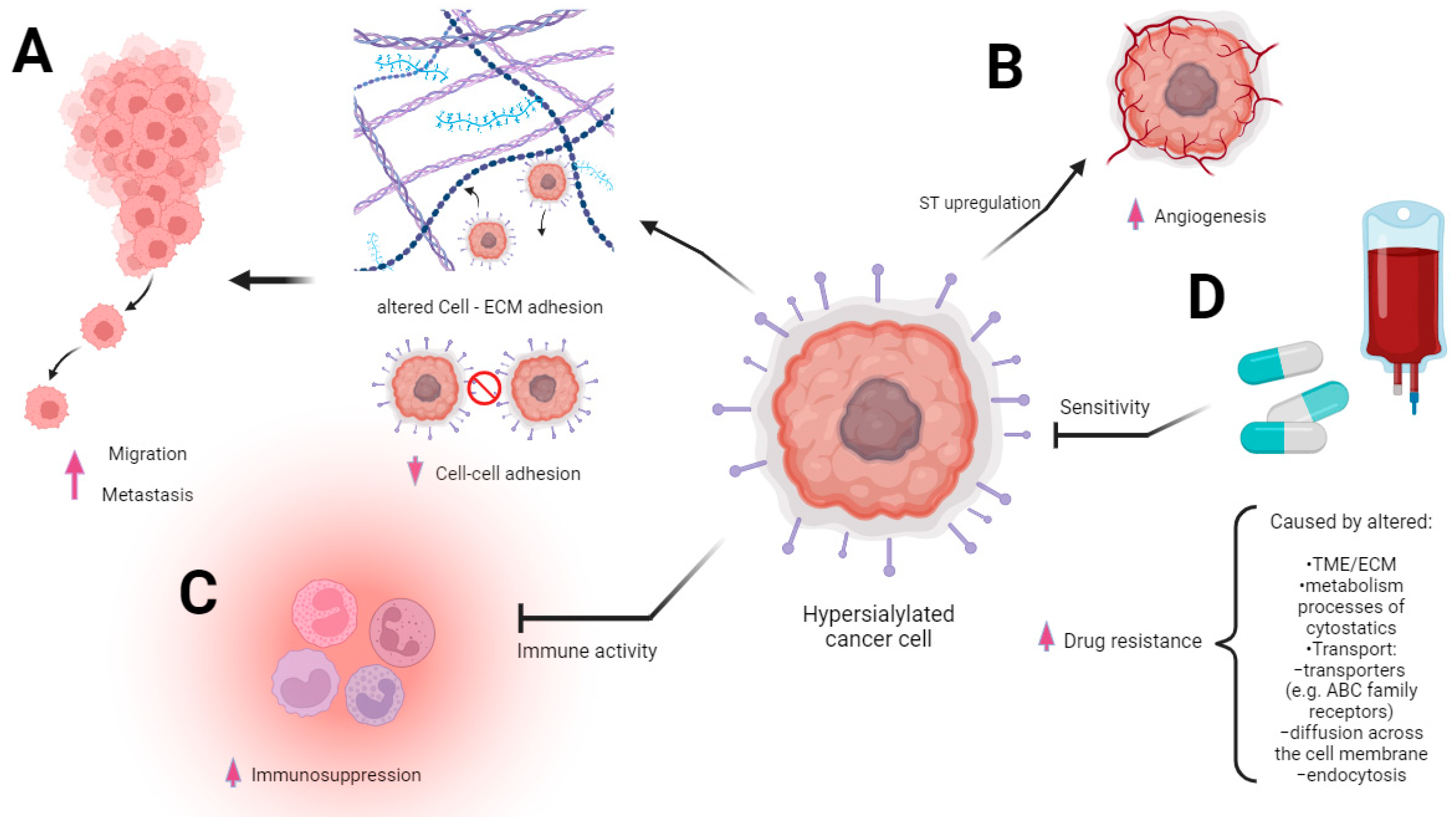
2.1. Sialic Acids and the Regulation of Cellular Functions in Cancer
2.2. Enzymatic Dysregulation of Sialylated Glycocalyx in Cancer
3. Drug Resistance in Cancer
3.1. Molecular Mechanisms of Drug Resistance in Cancer
3.2. Modulatory Effects of Sialoglycans on Drug Resistance—Related Processes
4. Sialic Acids as Targets and Cellular Indicators of Drug Action
4.1. Effects of Chemical Compounds on Sialome Machinery
4.2. Molecular and Cellular Consequences of Sialome Targeting
5. Sialic Acids as Modulators of Cellular Drug Sensitivity in Cancer
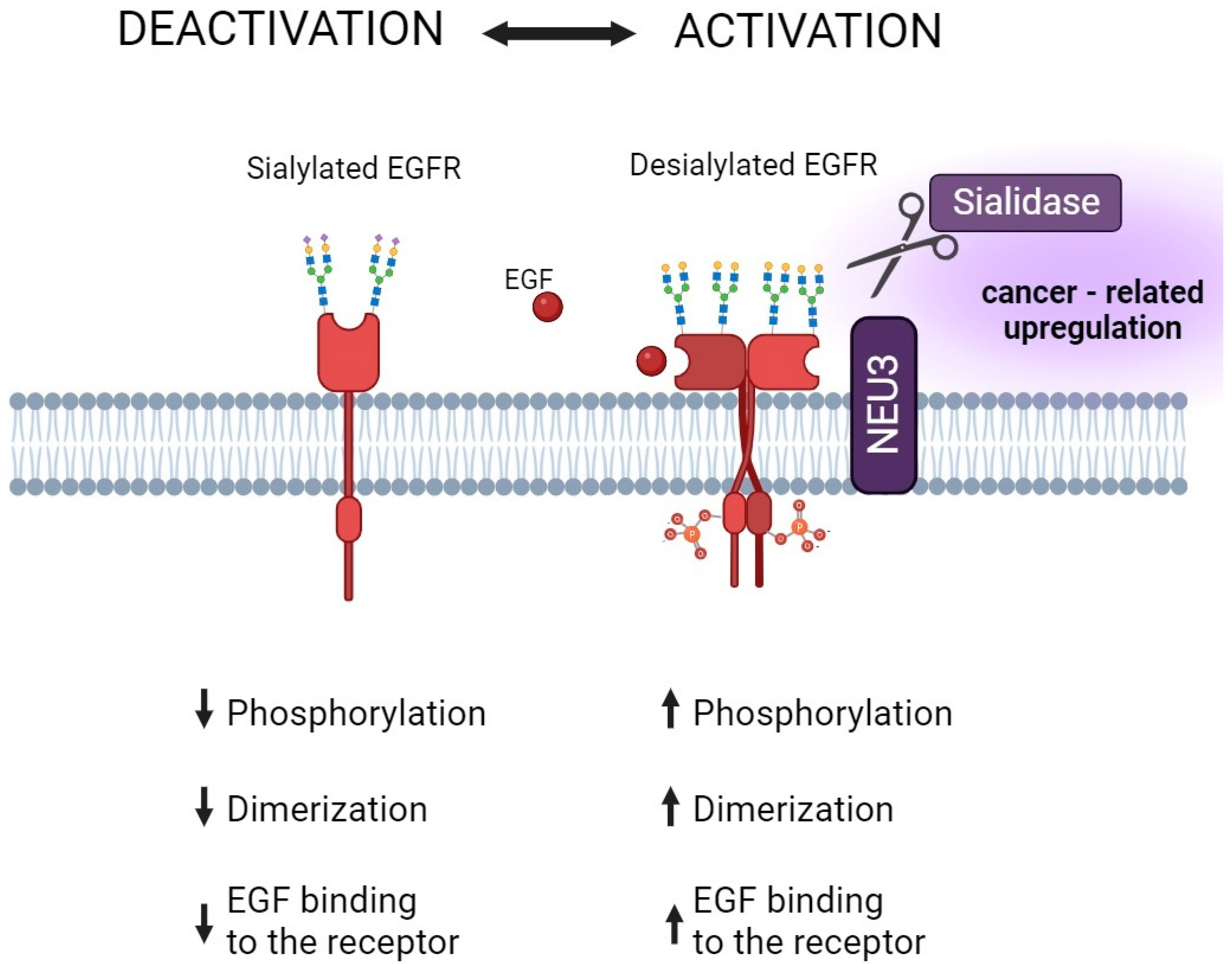
5.1. Sialylation and the Efficacy of Protein Kinase Inhibitors in Cancer Therapy
5.2. Sialylation and the Cellular Sensitivity to Cytotoxic Drugs
5.3. Sialidases as Therapeutic Target to Counter Chemoresistance
5.4. Sialic-Acid-Rich Glycocalyx and Chemoresistance in Brain Tumor Therapy
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cohen, M.; Varki, A. The sialome—Far more than the sum of its parts. Omics 2010, 14, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi, C.R.; Kiessling, L.L. Chemical glycobiology. Science 2001, 291, 2357–2364. [Google Scholar] [CrossRef] [PubMed]
- Dutta, H.; Jain, N. Post-translational modifications and their implications in cancer. Front. Oncol. 2023, 13, 1240115. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S. Sialic acid and biology of life: An introduction. In Sialic Acids and Sialoglycoconjugates in the Biology of Life, Health and Disease; Academic Press: Cambridge, MA, USA, 2020; pp. 1–61. [Google Scholar] [CrossRef]
- Ankenbauer, K.E.; Rao, T.C.; Mattheyses, A.L.; Bellis, S.L. Sialylation of EGFR by ST6GAL1 induces receptor activation and modulates trafficking dynamics. J Biol Chem. 2023, 299, 105217. [Google Scholar] [CrossRef] [PubMed]
- Fliniaux, I.; Marchand, G.; Molinaro, C.; Decloquement, M.; Martoriati, A.; Marin, M.; Bodart, J.F.; Harduin-Lepers, A.; Cailliau, K. Diversity of sialic acids and sialoglycoproteins in gametes and at fertilization. Front. Cell Dev. Biol. 2022, 10, 982931. [Google Scholar] [CrossRef]
- Dobie, C.; Skropeta, D. Insights into the role of sialylation in cancer progression and metastasis. Br. J. Cancer 2021, 124, 76–90. [Google Scholar] [CrossRef]
- Munkley, J. Aberrant Sialylation in Cancer: Therapeutic Opportunities. Cancers 2022, 14, 4248. [Google Scholar] [CrossRef]
- Pinho, S.; Reis, C. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Pally, D.; Pramanik, D.; Hussain, S.; Verma, S.; Srinivas, A.; Kumar, R.V.; Everest-Dass, A.; Bhat, R. Heterogeneity in 2,6-Linked Sialic Acids Potentiates Invasion of Breast Cancer Epithelia. ACS Cent. Sci. 2021, 7, 110–125. [Google Scholar] [CrossRef]
- Seales, E.C.; Jurado, G.A.; Brunson, B.A.; Wakefield, J.K.; Frost, A.R.; Bellis, S.L. Hypersialylation of beta1 integrins, observed in colon adenocarcinoma, may contribute to cancer progression by up-regulating cell motility. Cancer Res. 2005, 65, 4645–4652. [Google Scholar] [CrossRef]
- Kondo, K.; Harada, Y.; Nakano, M.; Suzuki, T.; Fukushige, T.; Hanzawa, K.; Yagi, H.; Takagi, K.; Mizuno, K.; Miyamoto, Y.; et al. Identification of distinct N-glycosylation patterns on extracellular vesicles from small-cell and non-small-cell lung cancer cells. J. Biol. Chem. 2022, 298, 101950. [Google Scholar] [CrossRef]
- Yamamoto, H.; Oviedo, A.; Sweeley, C.; Saito, T.; Moskal, J.R. Alpha2,6-sialylation of cell-surface N-glycans inhibits glioma formation in vivo. Cancer Res. 2001, 61, 6822–6829. [Google Scholar] [PubMed]
- Büll, C.; Stoel, M.A.; den Brok, M.H.; Adema, G.J. Sialic acids sweeten a tumor’s life. Cancer Res. 2014, 74, 3199–3204. [Google Scholar] [CrossRef] [PubMed]
- Ghirardello, M.; Shyam, R.; Galan, M.C. Reengineering of cancer cell surface charges can modulate cell migration. Chem. Commun. 2022, 58, 5522–5525. [Google Scholar] [CrossRef] [PubMed]
- Vattepu, R.; Sneed, S.L.; Anthony, R.M. Sialylation as an Important Regulator of Antibody Function. Front. Immunol. 2022, 13, 818736. [Google Scholar] [CrossRef]
- Mindler, K.; Ostertag, E.; Stehle, T. The polyfunctional polysialic acid: A structural view. Carbohydr. Res. 2021, 507, 108376. [Google Scholar] [CrossRef]
- Suzuki, M.; Suzuki, M.; Nakayama, J.; Suzuki, A.; Angata, K.; Chen, S.; Sakai, K.; Hagihara, K.; Yamaguchi, Y.; Fukuda, M. Polysialic acid facilitates tumor invasion by glioma cells. Glycobiology 2005, 15, 887–894. [Google Scholar] [CrossRef]
- Fujimoto, I.; Bruses, J.L.; Rutishauser, U. Regulation of cell adhesion by polysialic acid. Effects on cadherin, immunoglobulin cell adhesion molecule, and integrin function and independence from neural cell adhesion molecule binding or signaling activity. J. Biol. Chem. 2001, 276, 31745–31751. [Google Scholar] [CrossRef]
- Gascon, E.; Vutskits, L.; Kiss, J.Z. Polysialic acid–neural cell adhesion molecule in brain plasticity: From synapses to integration of new neurons. Brain Res. Rev. 2007, 56, 101–118. [Google Scholar] [CrossRef]
- Yang, P.; Yin, X.; Rutishauser, U. Intercellular space is affected by the polysialic acid content of NCAM. J. Cell Biol. 1992, 116, 1487–1496. [Google Scholar] [CrossRef]
- Hoffman, S.; Edelman, G.M. Kinetics of homophilic binding by embryonic and adult forms of the neural cell adhesion molecule. Proc. Natl. Acad. Sci. USA 1983, 80, 5762–5766. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Elkashef, S.M.; Allison, S.J.; Sadiq, M.; Basheer, H.A.; Ribeiro Morais, G.; Loadman, P.M.; Pors, K.; Falconer, R.A. Polysialic acid sustains cancer cell survival and migratory capacity in a hypoxic environment. Sci. Rep. 2016, 6, 33026. [Google Scholar] [CrossRef] [PubMed]
- Borsig, L. Selectins in cancer immunity. Glycobiology 2018, 28, 648–655. [Google Scholar] [CrossRef]
- Liang, J.X.; Liang, Y.; Gao, W. Clinicopathological and prognostic significance of sialyl Lewis X overexpression in patients with cancer: A meta-analysis. Onco Targets Ther. 2016, 9, 3113–3125. [Google Scholar] [PubMed]
- Almaraz, R.T.; Tian, Y.; Bhattarcharya, R.; Tan, E.; Chen, S.H.; Dallas, M.R.; Chen, L.; Zhang, Z.; Zhang, H.; Konstantopoulos, K.; et al. Metabolic flux increases glycoprotein sialylation: Implications for cell adhesion and cancer metastasis. Mol. Cell. Proteom. 2012, 11, 017558. [Google Scholar] [CrossRef]
- Miyagi, T.; Yamaguchi, K. Mammalian sialidases: Physiological and pathological roles in cellular functions. Glycobiology 2012, 22, 880–896. [Google Scholar] [CrossRef]
- Miyagi, T.; Takahashi, K.; Hata, K.; Shiozaki, K.; Yamaguchi, K. Sialidase significance for cancer progression. Glycoconj. J. 2012, 29, 567–577. [Google Scholar] [CrossRef]
- Gc, S.; Bellis, S.L.; Hjelmeland, A.B. ST6Gal1: Oncogenic signaling pathways and targets. Front. Mol. Biosci. 2022, 9, 962908. [Google Scholar] [CrossRef]
- Xu, C.; Wang, S.; Wu, Y.; Sun, X.; Yang, D.; Wang, S. Recent advances in understanding the roles of sialyltransferases in tumor angiogenesis and metastasis. Glycoconj. J. 2021, 38, 119–127. [Google Scholar] [CrossRef]
- Dorsett, K.A.; Marciel, M.P.; Hwang, J.; Ankenbauer, K.E.; Bhalerao, N.; Bellis, S.L. Regulation of ST6GAL1 sialyltransferase expression in cancer cells. Glycobiology 2021, 31, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Moreno, M.; Visser, E.A.; Hodgson, K.; Hipgrave Ederveen, A.L.; Bastian, K.; Goode, E.A.; Öztürk, Ö.; Pijnenborg, J.F.A.; Eerden, N.; Moons, S.J.; et al. Targeting aberrant sialylation and fucosylation in prostate cancer cells using potent metabolic inhibitors. Glycobiology 2023, cwad085. [Google Scholar] [CrossRef]
- Wichert, B.; Milde-Langosch, K.; Galatenko, V.; Schmalfeldt, B.; Oliveira-Ferrer, L. Prognostic role of the sialyltransferase ST6GAL1 in ovarian cancer. Glycobiology 2018, 28, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, H.; Ushijima, T. Accumulation of genetic and epigenetic alterations in normal cells and cancer risk. NPJ Precis. Onc. 2019, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Greville, G.; McCann, A.; Rudd, P.M.; Saldova, R. Epigenetic regulation of glycosylation and the impact on chemo-resistance in breast and ovarian cancer. Epigenetics 2016, 11, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, A.; Relvas-Santos, M.; Azevedo, R.; Santos, L.L.; Ferreira, J.A. Protein Glycosylation and Tumor Microenvironment Alterations Driving Cancer Hallmarks. Front. Oncol. 2019, 9, 380. [Google Scholar] [CrossRef]
- Li, C.; Teixeira, A.F.; Zhu, H.J.; Ten Dijke, P. Cancer associated-fibroblast-derived exosomes in cancer progression. Mol. Cancer 2021, 20, 154. [Google Scholar] [CrossRef]
- Barallobre-Barreiro, J.; Baig, F.; Fava, M.; Yin, X.; Mayr, M. Glycoproteomics of the Extracellular Matrix: A Method for Intact Glycopeptide Analysis Using Mass Spectrometry. J. Vis. Exp. 2017, 122, 55674. [Google Scholar] [CrossRef]
- Walker, C.; Mojares, E.; Del Río Hernández, A. Role of Extracellular Matrix in Development and Cancer Progression. Int. J. Mol. Sci. 2018, 19, 3028. [Google Scholar] [CrossRef]
- Bassagañas, S.; Pérez-Garay, M.; Peracaula, R. Cell surface sialic acid modulates extracellular matrix adhesion and migration in pancreatic adenocarcinoma cells. Pancreas 2014, 43, 109–117. [Google Scholar] [CrossRef]
- Coulson-Thomas, Y.M.; Gesteira, T.F.; Norton, A.L.; Kao, W.W.; Nader, H.B.; Coulson-Thomas, V.J. The role of proteoglycans in the reactive stroma on tumor growth and progression. Histol. Histopathol. 2015, 30, 33–41. [Google Scholar] [PubMed]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.; Waller, C.; Timpl, R.; Schirrmacher, V. Surface sialic acid reduces attachment of metastatic tumour cells to collagen type IV and fibronectin. Nature 1982, 300, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, O.; Abe, M.; Hashimoto, Y. Sialylation and glycosylation modulate cell adhesion and invasion to extracellular matrix in human malignant lymphoma: Dependency on integrin and the Rho GTPase family. Int. J. Oncol. 2015, 47, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, D.; Pally, D.; Saini, D.K.; Bhat, R.; Ghosh, A. Nanomotors Sense Local Physicochemical Heterogeneities in Tumor Microenvironments. Angew. Chem. Int. Ed. 2020, 59, 23690–23696. [Google Scholar] [CrossRef]
- Yu, S.; Fan, J.; Liu, L.; Zhang, L.; Wang, S.; Zhang, J. Caveolin-1 up-regulates integrin α2,6-sialylation to promote integrin α5β1-dependent hepatocarcinoma cell adhesion. FEBS Lett. 2013, 587, 782–787. [Google Scholar] [CrossRef]
- Su, C.Y.; Li, J.Q.; Zhang, L.L.; Wang, H.; Wang, F.H.; Tao, Y.W.; Wang, Y.Q.; Guo, Q.R.; Li, J.J.; Liu, Y.; et al. The Biological Functions and Clinical Applications of Integrins in Cancers. Front. Pharmacol. 2020, 11, 579068. [Google Scholar] [CrossRef]
- Yuan, Y.; Wu, L.; Shen, S.; Wu, S.; Burdick, M.M. Effect of alpha 2,6 sialylation on integrin-mediated adhesion of breast cancer cells to fibronectin and collagen IV. Life Sci. 2016, 149, 138–145. [Google Scholar] [CrossRef]
- Seales, E.C.; Jurado, G.A.; Singhal, A.; Bellis, S.L. Ras oncogene directs expression of a differentially sialylated, functionally altered beta1 integrin. Oncogene 2003, 22, 7137–7145. [Google Scholar] [CrossRef]
- Shaikh, F.M.; Seales, E.C.; Clem, W.C.; Hennessy, K.M.; Zhuo, Y.; Bellis, S.L. Tumor cell migration and invasion are regulated by expression of variant integrin glycoforms. Exp. Cell Res. 2008, 314, 2941–2950. [Google Scholar] [CrossRef]
- Mao, X.; Xu, J.; Wang, W.; Liang, C.; Hua, J.; Liu, J.; Zhang, B.; Meng, Q.; Yu, X.; Shi, S. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: New findings and future perspectives. Mol. Cancer 2021, 20, 131. [Google Scholar] [CrossRef]
- Sasano, T.; Gonzalez-Delgado, R.; Muñoz, N.M.; Carlos-Alcade, W.; Cho, M.S.; Sheth, R.A.; Sood, A.K.; Afshar-Kharghan, V. Podoplanin promotes tumor growth, platelet aggregation, and venous thrombosis in murine models of ovarian cancer. J. Thromb. Haemost. 2022, 20, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Oki, H.; Hozumi, Y.; Liu, X.; Ogasawara, S.; Takagi, M.; Goto, K.; Kato, Y. Monoclonal Antibody LpMab-9 Recognizes O-glycosylated N-Terminus of Human Podoplanin. Monoclon. Antib. Immunodiagn. Immunother. 2015, 34, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Kato, Y.; Kameyama, A.; Ito, H.; Kuno, A.; Hirabayashi, J.; Kubota, T.; Amano, K.; Chiba, Y.; Hasegawa, Y.; et al. Functional glycosylation of human podoplanin: Glycan structure of platelet aggregation-inducing factor. FEBS Lett. 2007, 581, 331–336. [Google Scholar] [CrossRef] [PubMed]
- van Houtum, E.J.H.; Büll, C.; Cornelissen, L.A.M.; Adema, G.J. Siglec Signaling in the Tumor Microenvironment. Front. Immunol. 2021, 12, 790317. [Google Scholar] [CrossRef]
- Gianchecchi, E.; Arena, A.; Fierabracci, A. Sialic Acid-Siglec Axis in Human Immune Regulation, Involvement in Autoimmunity and Cancer and Potential Therapeutic Treatments. Int. J. Mol. Sci. 2021, 22, 5774. [Google Scholar] [CrossRef]
- Crocker, P.; Paulson, J.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef]
- Jiang, K.Y.; Qi, L.L.; Kang, F.B.; Wang, L. The intriguing roles of Siglec family members in the tumor microenvironment. Biomark. Res. 2022, 10, 22. [Google Scholar] [CrossRef]
- Hudak, J.; Canham, S.; Bertozzi, C. Glycocalyx engineering reveals a Siglec-based mechanism for NK cell immunoevasion. Nat. Chem. Biol. 2014, 10, 69–75. [Google Scholar] [CrossRef]
- Assaraf, Y.G.; Brozovic, A.; Gonçalves, A.C.; Jurkovicova, D.; Linē, A.; Machuqueiro, M.; Saponara, S.; Sarmento-Ribeiro, A.B.; Xavier, C.P.R.; Vasconcelos, M.H. The multi-factorial nature of clinical multidrug resistance in cancer. Drug Resist. Updates 2019, 46, 100645. [Google Scholar] [CrossRef]
- Kodan, A.; Futamata, R.; Kimura, Y.; Kioka, N.; Nakatsu, T.; Kato, H.; Ueda, K. ABCB1/MDR1/P-gp employs an ATP-dependent twist-and-squeeze mechanism to export hydrophobic drugs. FEBS Lett. 2021, 595, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Tanigawara, Y. Role of P-Glycoprotein in Drug Disposition. Ther. Drug Monit. 2000, 22, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Pilotto Heming, C.; Muriithi, W.; Wanjiku Macharia, L.; Niemeyer Filho, P.; Moura-Neto, V.; Aran, V. P-glycoprotein and cancer: What do we currently know? Heliyon 2022, 8, e11171. [Google Scholar] [CrossRef]
- Fu, D. Where is it and How Does it Get There—Intracellular Localization and Traffic of P-glycoprotein. Front. Oncol. 2013, 3, 321. [Google Scholar] [CrossRef] [PubMed]
- Hanke, N.T.; Imler, E.; Marron, M.T.; Seligmann, B.E.; Garland, L.L.; Baker, A.F. Characterization of carfilzomib-resistant non-small cell lung cancer cell lines. J. Cancer Res. Clin. Oncol. 2018, 144, 1317–1327. [Google Scholar] [CrossRef]
- Bakos, É.; Homolya, L. Portrait of multifaceted transporter, the multidrug resistance-associated protein 1 (MRP1/ABCC1). Pflug. Arch—Eur. J. Physiol. 2007, 453, 621–641. [Google Scholar] [CrossRef]
- Kartal-Yandim, M.; Adan-Gokbulut, A.; Baran, Y. Molecular mechanisms of drug resistance and its reversal in cancer. Crit. Rev. Biotechnol. 2016, 36, 716–726. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Tew, K.D. Glutathione-associated enzymes in anticancer drug resistance. Cancer Res. 1994, 54, 4313–4320. [Google Scholar] [CrossRef]
- Triller, N.; Korosec, P.; Kern, I.; Kosnik, M.; Debeljak, A. Multidrug resistance in small cell lung cancer: Expression of P-glycoprotein, multidrug resistance protein 1 and lung resistance protein in chemo-naive patients and in relapsed disease. Lung Cancer 2006, 54, 235–240. [Google Scholar] [CrossRef]
- Gottesman, M.M. Mechanisms of cancer drug resistance. Annu. Rev. Med. 2002, 53, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhou, H.; Liu, G.; Wu, J.; Yuan, Y.; Shang, A. Tumor Microenvironment: Lactic Acid Promotes Tumor Development. J. Immunol. Res. 2022, 2022, 3119375. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Choi, S.Y.C.; Niu, X.; Kang, N.; Xue, H.; Killam, J.; Wang, Y. Lactic Acid and an Acidic Tumor Microenvironment suppress Anticancer Immunity. Int. J. Mol. Sci. 2020, 21, 8363. [Google Scholar] [CrossRef]
- Liu, R.; Chen, Y.; Liu, G.; Li, C.; Song, Y.; Cao, Z.; Li, W.; Hu, J.; Lu, C.; Liu, Y. PI3K/AKT pathway as a key link modulates the multidrug resistance of cancers. Cell Death Dis. 2020, 11, 797. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, W.; Zhou, H.; Li, H.; Wang, N.; Miao, X.; Jia, L. α-2,8-sialyltransferase is involved in the development of multidrug resistance via PI3K/Akt pathway in human chronic myeloid leukemia. IUBMB Life 2015, 67, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, X.; Meng, F.; Huang, T.; Wang, S.; Zheng, Z.; Zheng, G.; Li, W.; Zhang, J.; Liu, Y. α2,6-Sialylation promotes hepatocellular carcinoma cells migration and invasion via enhancement of nSmase2-mediated exosomal miRNA sorting. J. Physiol. Biochem. 2023, 79, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Wei, A.; Fan, B.; Zhao, Y.; Zhang, H.; Wang, L.; Yu, X.; Yuan, Q.; Yang, D.; Wang, S. ST6Gal-I overexpression facilitates prostate cancer progression via the PI3K/Akt/GSK-3β/β-catenin signaling pathway. Oncotarget 2016, 7, 65374–65388. [Google Scholar] [CrossRef]
- Nath, S.; Mandal, C.; Chatterjee, U.; Mandal, C. Association of cytosolic sialidase Neu2 with plasma membrane enhances Fas-mediated apoptosis by impairing PI3K-Akt/mTOR-mediated pathway in pancreatic cancer cells. Cell Death Dis. 2018, 9, 210. [Google Scholar] [CrossRef]
- Jurj, A.; Ionescu, C.; Berindan-Neagoe, I.; Braicu, C. The extracellular matrix alteration, implication in modulation of drug resistance mechanism: Friends or foes? J. Exp. Clin. Cancer Res. 2022, 41, 276. [Google Scholar] [CrossRef]
- Henke, E.; Nandigama, R.; Ergün, S. Extracellular Matrix in the Tumor Microenvironment and Its Impact on Cancer Therapy. Front. Mol. Biosci. 2020, 6, 160. [Google Scholar] [CrossRef]
- Saw, P.E.; Chen, J.; Song, E. Targeting CAFs to overcome anticancer therapeutic resistance. Trends Cancer 2022, 8, 527–555. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef] [PubMed]
- De, P.; Aske, J.; Sulaiman, R.; Dey, N. Bête Noire of Chemotherapy and Targeted Therapy: CAF-Mediated Resistance. Cancers 2022, 14, 1519. [Google Scholar] [CrossRef] [PubMed]
- Mehraj, U.; Dar, A.H.; Wani, N.A.; Mir, M.A. Tumor microenvironment promotes breast cancer chemoresistance. Cancer Chemother. Pharmacol. 2021, 87, 147–158. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P. The interaction of anticancer therapies with tumor-associated macrophages. J. Exp. Med. 2015, 212, 435–445. [Google Scholar] [CrossRef]
- Olson, O.C.; Kim, H.; Quail, D.F.; Foley, E.A.; Joyce, J.A. Tumor-Associated Macrophages Suppress the Cytotoxic Activity of Antimitotic Agents. Cell Rep. 2017, 19, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Yao, S.; Hu, Y.; Feng, Y.; Li, M.; Bian, Z.; Zhang, J.; Qin, Y.; Qi, X.; Zhou, L.; et al. The Immune-microenvironment Confers Chemoresistance of Colorectal Cancer through Macrophage-Derived IL6. Clin. Cancer Res. 2017, 23, 7375–7387. [Google Scholar] [CrossRef]
- Feng, D.D.; Zhang, H.; Zhang, P.; Zheng, Y.S.; Zhang, X.J.; Han, B.W.; Luo, X.Q.; Xu, L.; Zhou, H.; Qu, L.H.; et al. Down-regulated miR-331-5p and miR-27a are associated with chemotherapy resistance and relapse in leukaemia. J. Cell. Mol. Med. 2011, 15, 2164–2175. [Google Scholar] [CrossRef]
- Yang, H.; Kong, W.; He, L.; Zhao, J.J.; O’Donnell, J.D.; Wang, J.; Wenham, R.M.; Coppola, D.; Kruk, P.A.; Nicosia, S.V.; et al. MicroRNA expression profiling in human ovarian cancer: miR-214 induces cell survival and cisplatin resistance by targeting PTEN. Cancer Res. 2008, 68, 425–433. [Google Scholar] [CrossRef]
- Hu, C.; Xia, R.; Zhang, X.; Li, T.; Ye, Y.; Li, G.; He, R.; Li, Z.; Lin, Q.; Zheng, S.; et al. circFARP1 enables cancer-associated fibroblasts to promote gemcitabine resistance in pancreatic cancer via the LIF/STAT3 axis. Mol. Cancer 2022, 21, 24. [Google Scholar] [CrossRef]
- Yang, J.; Liu, S.; Du, L.; Jiang, S. A new role of neuraminidase (NA) in the influenza virus life cycle: Implication for developing NA inhibitors with novel mechanism of action. Rev. Med. Virol. 2016, 26, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, T.; Yamamoto, K. Sialidase NEU3 and its pathological significance. Glycoconj. J. 2022, 39, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, E.P.; Luzina, I.G.; Atamas, S.P. Mammalian Neuraminidases in Immune-Mediated Diseases: Mucins and Beyond. Front. Immunol. 2022, 13, 883079. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.Q.; Ma, G.; Liu, J.F.; Cai, Y.Y.; Zhang, J.Y.; Wei, T.T.; Pan, A.; Jiang, S.; Xiao, Y.; Xiao, P.; et al. Neuraminidase 1 is a driver of experimental cardiac hypertrophy. Eur. Heart J. 2021, 42, 3770–3782. [Google Scholar] [CrossRef]
- de Oliveira, J.T.; Santos, A.L.; Gomes, C.; Barros, R.; Ribeiro, C.; Mendes, N.; de Matos, A.J.; Vasconcelos, M.H.; Oliveira, M.J.; Reis, C.A.; et al. Anti-Influenza Neuraminidase Inhibitor Oseltamivir Phosphate Induces Canine Mammary Cancer Cell Aggressiveness. PLoS ONE 2015, 10, e0121590. [Google Scholar] [CrossRef]
- Berghuis, A.Y.; Pijnenborg, J.F.A.; Boltje, T.J.; Pijnenborg, J.M.A. Sialic acids in gynecological cancer development and progression: Impact on diagnosis and treatment. Int. J. Cancer 2022, 150, 678–687. [Google Scholar] [CrossRef]
- Heise, T.; Pijnenborg, J.F.A.; Büll, C.; van Hilten, N.; Kers-Rebel, E.D.; Balneger, N.; Elferink, H.; Adema, G.J.; Boltje, T.J. Potent Metabolic Sialylation Inhibitors Based on C-5-Modified Fluorinated Sialic Acids. J. Med. Chem. 2019, 62, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Rillahan, C.D.; Antonopoulos, A.; Lefort, C.T.; Sonon, R.; Azadi, P.; Ley, K.; Dell, A.; Haslam, S.M.; Paulson, J.C. Global metabolic inhibitors of sialyl- and fucosyltransferases remodel the glycome. Nat. Chem. Biol. 2012, 8, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Büll, C.; Boltje, T.J.; Wassink, M.; de Graaf, A.M.; van Delft, F.L.; den Brok, M.H.; Adema, G.J. Targeting aberrant sialylation in cancer cells using a fluorinated sialic acid analog impairs adhesion, migration, and in vivo tumor growth. Mol. Cancer Ther. 2013, 12, 1935–1946. [Google Scholar] [CrossRef] [PubMed]
- Büll, C.; Boltje, T.J.; van Dinther, E.A.; Peters, T.; de Graaf, A.M.; Leusen, J.H.; Kreutz, M.; Figdor, C.G.; den Brok, M.H.; Adema, G.J. Targeted delivery of a sialic acid-blocking glycomimetic to cancer cells inhibits metastatic spread. ACS Nano 2015, 9, 733–745. [Google Scholar] [CrossRef]
- Marquina, G.; Waki, H.; Fernandez, L.E.; Kon, K.; Carr, A.; Valiente, O.; Perez, R.; Ando, S. Gangliosides expressed in human breast cancer. Cancer Res. 1996, 56, 5165–5171. [Google Scholar] [PubMed]
- Casadesús, A.V.; Fernández-Marrero, Y.; Clavell, M.; Gómez, J.A.; Hernández, T.; Moreno, E.; López-Requena, A. A shift from N-glycolyl- to N-acetyl-sialic acid in the GM3 ganglioside impairs tumor development in mouse lymphocytic leukemia cells. Glycoconj. J. 2013, 7, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Labrada, M.; Dorvignit, D.; Hevia, G.; Rodríguez-Zhurbenko, N.; Hernández, A.M.; Vázquez, A.M.; Fernández, L.E. GM3(Neu5Gc) ganglioside. An evolution fixed neoantigen for cancer immunotherapy. Semin. Oncol. 2018, 45, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Segatori, V.I.; Gulino, C.A.; Capobianco, C.S.; Rojo, S.; Ferreira, G.M.; Cuello, H.A.; Gabri, M.R.; Alonso, D.A. Abstract 6648: Racotumomab and PD-1 blockade combination exhibits an additive antitumor effect in a non-small cell lung cancer model. Cancer Res. 2020, 80 (Suppl. S16), 6648. [Google Scholar] [CrossRef]
- Uskent, N.; Ayla, S.; Molinas Mandel, N.; Ozkan, M.; Teomete, M.; Baloglu, H.; Aydıncer, C.; Yergok, H.; Dogan, E.; Berk, B.; et al. Prognostic Significance of Tumor Tissue NeuGcGM3 Ganglioside Expression in Patients Receiving Racotumomab Immunotherapy. J. Oncol. 2020, 2020, 1360431. [Google Scholar] [CrossRef]
- Daly, J.; Carlsten, M.; O’Dwyer, M. Sugar Free: Novel Immunotherapeutic Approaches Targeting Siglecs and Sialic Acids to Enhance Natural Killer Cell Cytotoxicity Against Cancer. Front. Immunol. 2019, 10, 1047. [Google Scholar] [CrossRef]
- Perez, S.J.L.P.; Fu, C.W.; Li, W.S. Sialyltransferase Inhibitors for the Treatment of Cancer Metastasis: Current Challenges and Future Perspectives. Molecules 2021, 26, 5673. [Google Scholar] [CrossRef]
- Hindenburg, A.A.; Taub, R.N.; Grant, S.; Chang, G.; Baker, M.A. Effects of Pyrimidine Antagonists on Sialic Acid Regeneration in HL-60 Cells1. Cancer Res. 1985, 45, 3048–3052. [Google Scholar]
- Gińdzieńska-Sieśkiewicz, E.; Radziejewska, I.; Domysławska, I.; Klimiuk, P.A.; Sulik, A.; Rojewska, J.; Gabryel-Porowska, H.; Sierakowski, S. Changes of glycosylation of IgG in rheumatoid arthritis patients treated with methotrexate. Adv. Med. Sci. 2016, 61, 193–197. [Google Scholar] [CrossRef]
- Deng, X.; Liu, X.; Zhang, Y.; Ke, D.; Yan, R.; Wang, Q.; Tian, X.; Li, M.; Zeng, X.; Hu, C. Changes of serum IgG glycosylation patterns in rheumatoid arthritis. Clin. Proteom. 2023, 20, 7. [Google Scholar] [CrossRef]
- Lundström, S.L.; Hensvold, A.H.; Rutishauser, D.; Klareskog, L.; Ytterberg, A.J.; Zubarev, R.A.; Catrina, A.I. IgG Fc galactosylation predicts response to methotrexate in early rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 182. [Google Scholar] [CrossRef]
- Qorri, B.; Harless, W.; Szewczuk, M.R. Novel Molecular Mechanism of Aspirin and Celecoxib Targeting Mammalian Neuraminidase-1 Impedes Epidermal Growth Factor Receptor Signaling Axis and Induces Apoptosis in Pancreatic Cancer Cells. Drug Des. Dev. Ther. 2020, 14, 4149–4167. [Google Scholar] [CrossRef] [PubMed]
- Qorri, B.; Mokhtari, R.B.; Harless, W.W.; Szewczuk, M.R. Repositioning of Old Drugs for Novel Cancer Therapies: Continuous Therapeutic Perfusion of Aspirin and Oseltamivir Phosphate with Gemcitabine Treatment Disables Tumor Progression, Chemoresistance, and Metastases. Cancers 2022, 14, 3595. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.U.; Malik, S.A.; Bashir, M.; Khan, R.U.; Idrees, M. Serum sialic acid changes in type 2 diabetic patients on metformin or rosiglitazone treatment. J. Clin. Pharm. Ther. 2010, 35, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein glycosylation in cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef]
- Lee, M.; Lee, H.J.; Bae, S.; Lee, Y.S. Protein sialylation by sialyltransferase involves radiation resistance. Mol. Cancer Res. 2008, 6, 1316–1325. [Google Scholar] [CrossRef]
- Nag, S.; Mandal, A.; Joshi, A.; Jain, N.; Srivastava, R.S.; Singh, S.; Khattri, A. Sialyltransferases and Neuraminidases: Potential Targets for Cancer Treatment. Diseases 2022, 10, 114. [Google Scholar] [CrossRef]
- Weber, M.; Modemann, S.; Schipper, P.; Trauer, H.; Franke, H.; Illes, P.; Geiger, K.D.; Hengstler, J.G.; Kleemann, W.J. Increased polysialic acid neural cell adhesion molecule expression in human hippocampus of heroin addicts. Neuroscience 2006, 138, 1215–1223. [Google Scholar] [CrossRef]
- Shetlar, M.R. Brain glycoconjugates in alcohol and narcotic research. Neurobehav. Toxicol. Teratol. 1985, 7, 113–118. [Google Scholar]
- Azuine, M.A.; Patel, S.J.; Lakshman, M.R. Effects of chronic ethanol administration on the activities and relative synthetic rates of myelin and synaptosomal plasma membrane-associated sialidase in the rat brain. Neurochem. Int. 2006, 48, 67–74. [Google Scholar] [CrossRef]
- Boyzo, A.; Ayala, J.; Gutiérrez, R.; Hernández-R, J. Neuraminidase activity in different regions of the seizing epileptic and non-epileptic brain. Brain Res. 2003, 964, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Wielgat, P.; Walesiuk, A.; Braszko, J.J. Effects of chronic stress and corticosterone on sialidase activity in the rat hippocampus. Behav. Brain Res. 2011, 222, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Smithson, M.; Irwin, R.; Williams, G.; Alexander, K.L.; Smythies, L.E.; Nearing, M.; McLeod, M.C.; Al Diffalha, S.; Bellis, S.L.; Hardiman, K.M. Sialyltransferase ST6GAL-1 mediates resistance to chemoradiation in rectal cancer. J. Biol. Chem. 2022, 298, 101594. [Google Scholar] [CrossRef] [PubMed]
- Park, J.J.; Yi, J.Y.; Jin, Y.B.; Lee, Y.J.; Lee, J.S.; Lee, Y.S.; Ko, Y.G.; Lee, M. Sialylation of epidermal growth factor receptor regulates receptor activity and chemosensitivity to gefitinib in colon cancer cells. Biochem. Pharmacol. 2012, 83, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Britain, C.M.; Holdbrooks, A.T.; Anderson, J.C.; Willey, C.D.; Bellis, S.L. Sialylation of EGFR by the ST6Gal-I sialyltransferase promotes EGFR activation and resistance to gefitinib-mediated cell death. J. Ovar. Res. 2018, 11, 12. [Google Scholar] [CrossRef]
- Yen, H.Y.; Liu, Y.C.; Chen, N.Y.; Tsai, C.F.; Wang, Y.T.; Chen, Y.J.; Hsu, T.L.; Yang, P.C.; Wong, C.H. Effect of sialylation on EGFR phosphorylation and resistance to tyrosine kinase inhibition. Proc. Natl. Acad. Sci. USA 2015, 112, 6955–6960. [Google Scholar] [CrossRef]
- Liu, N.; Zhu, M.; Linhai, Y.; Song, Y.; Gui, X.; Tan, G.; Li, J.; Liu, Y.; Deng, Z.; Chen, X.; et al. Increasing HER2 α2,6 sialylation facilitates gastric cancer progression and resistance via the Akt and ERK pathways. Oncol. Rep. 2018, 40, 2997–3005. [Google Scholar] [CrossRef]
- Ou, L.; He, X.; Liu, N.; Song, Y.; Li, J.; Gao, L.; Huang, X.; Deng, Z.; Wang, X.; Lin, S. Sialylation of FGFR1 by ST6Gal-I overexpression contributes to ovarian cancer cell migration and chemoresistance. Mol. Med. Rep. 2020, 21, 1449–1460. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, J.; Ruan, Y.; Sun, L.; Xu, C.; Jiang, H. Sialyltransferase ST3GAL1 promotes cell migration, invasion, and TGF-β1-induced EMT and confers paclitaxel resistance in ovarian cancer. Cell Death Dis. 2018, 9, 1102. [Google Scholar] [CrossRef]
- Huang, S.; Day, T.W.; Choi, M.R.; Safa, A.R. Human beta-galactoside alpha-2,3-sialyltransferase (ST3Gal III) attenuated Taxol-induced apoptosis in ovarian cancer cells by downregulating caspase-8 activity. Mol. Cell Biochem. 2009, 331, 81–88. [Google Scholar] [CrossRef]
- Schultz, M.J.; Swindall, A.F.; Wright, J.W.; Sztul, E.S.; Landen, C.N.; Bellis, S.L. ST6Gal-I sialyltransferase confers cisplatin resistance in ovarian tumor cells. J. Ovar. Res. 2013, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Imaeda, Y.; Umemoto, S.; Kobayashi, K.; Suzuki, H.; Okamoto, T. Cimetidine increases survival of colorectal cancer patients with high levels of sialyl Lewis-X and sialyl Lewis-A epitope expression on tumour cells. Br. J. Cancer 2002, 86, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Wattanavises, S.; Silsirivanit, A.; Sawanyawisuth, K.; Cha’on, U.; Waraasawapati, S.; Saentaweesuk, W.; Luang, S.; Chalermwat, C.; Wongkham, C.; Wongkham, S. Increase of MAL-II Binding Alpha2,3-Sialylated Glycan Is Associated with 5-FU Resistance and Short Survival of Cholangiocarcinoma Patients. Medicina 2019, 55, 761. [Google Scholar] [CrossRef]
- Li, Y.; Luo, S.; Dong, W.; Song, X.; Zhou, H.; Zhao, L.; Jia, L. Alpha-2, 3-sialyltransferases regulate the multidrug resistance of chronic myeloid leukemia through miR-4701-5p targeting ST3GAL1. Lab. Investig. 2016, 96, 731–740. [Google Scholar] [CrossRef]
- Mei, J.; Lin, W.; Li, S.; Tang, Y.; Ye, Z.; Lu, L.; Wen, Y.; Kan, A.; Zou, J.; Yu, C.; et al. Long noncoding RNA TINCR facilitates hepatocellular carcinoma progression and dampens chemosensitivity to oxaliplatin by regulating the miR-195-3p/ST6GAL1/NF-κB pathway. J. Exp. Clin. Cancer Res. 2022, 41, 5. [Google Scholar] [CrossRef]
- Zhao, L.; Li, Y.; Song, X.; Zhou, H.; Li, N.; Miao, Y.; Jia, L. Upregulation of miR-181c inhibits chemoresistance by targeting ST8SIA4 in chronic myelocytic leukemia. Oncotarget 2016, 7, 60074–60086. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Y.; Zhao, L.; Pan, Y.; Shan, Y.; Li, Y.; Jia, L. Upregulation of microRNA-135b and microRNA-182 promotes chemoresistance of colorectal cancer by targeting ST6GALNAC2 via PI3K/AKT pathway. Mol. Carcinog. 2017, 56, 2669–2680. [Google Scholar] [CrossRef]
- Haxho, F.; Neufeld, R.J.; Szewczuk, M.R. Neuraminidase-1: A novel therapeutic target in multistage tumorigenesis. Oncotarget 2016, 7, 40860–40881. [Google Scholar] [CrossRef]
- O’Shea, L.K.; Abdulkhalek, S.; Allison, S.; Neufeld, R.J.; Szewczuk, M.R. Therapeutic targeting of Neu1 sialidase with oseltamivir phosphate (Tamiflu®) disables cancer cell survival in human pancreatic cancer with acquired chemoresistance. Onco Targets Ther. 2014, 7, 117–134. [Google Scholar]
- Moore, M.L.; Chi, M.H.; Zhou, W.; Goleniewska, K.; O’Neal, J.F.; Higginbotham, J.N.; Peebles, R.S., Jr. Cutting Edge: Oseltamivir decreases T cell GM1 expression and inhibits clearance of respiratory syncytial virus: Potential role of endogenous sialidase in antiviral immunity. J. Immunol. 2007, 178, 2651–2654. [Google Scholar] [CrossRef]
- Hama, R.; Bennett, C.L. The mechanisms of sudden-onset type adverse reactions to oseltamivir. Acta Neurol. Scand. 2017, 135, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Benvindo, C.; Leijs, A.A.; Thaler, M.; Tas, A.; Arbiser, J.L.; Snijder, E.J.; van Hemert, M.J. Honokiol Inhibits SARS-CoV-2 Replication in Cell Culture at a Post-Entry Step. Microbiol. Spectr. 2023, 11, e0327322. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Jiang, X.; Zhao, M.; Su, K.; Tang, M.; Hong, F.; Ye, N.; Zhang, R.; Li, N.; Wang, L.; et al. Identification of the target protein and molecular mechanism of honokiol in anti-inflammatory action. Phytomedicine 2023, 109, 154617. [Google Scholar] [CrossRef] [PubMed]
- Amorati, R.; Zotova, J.; Baschieri, A.; Valgimigli, L. Antioxidant Activity of Magnolol and Honokiol: Kinetic and Mechanistic Investigations of Their Reaction with Peroxyl Radicals. J. Org. Chem. 2015, 80, 10651–10659. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, F.; Qiao, Y.; Chen, T.; Fan, L.; Shen, X.; Yu, D.; Huang, Y.; Wei, M. Honokiol inhibits interleukin-induced angiogenesis in the NSCLC microenvironment through the NF-κB signaling pathway. Chem. Biol. Interact. 2023, 370, 110295. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, F.; Ali, S.; Kumar, V.; Elasbali, A.M.; Alhassan, H.H.; Alharethi, S.H.; Islam, A.; Hassan, M.I. Pharmacological features, health benefits and clinical implications of honokiol. J. Biomol. Struct. Dyn. 2023, 41, 7511–7533. [Google Scholar] [CrossRef]
- Thulasiraman, P.; Kerr, K.; McAlister, K.; Hardisty, S.; Wistner, A.; McCullough, I. Neuraminidase 1 regulates proliferation, apoptosis and the expression of Cadherins in mammary carcinoma cells. Mol. Cell. Biochem. 2019, 462, 207–215. [Google Scholar] [CrossRef]
- Chio, C.C.; Tai, Y.; Mohanraj, M.; Liu, S.H.; Yang, S.T.; Chen, R.M. Honokiol enhances temozolomide-induced apoptotic insults to malignant glioma cells via an intrinsic mitochondrion-dependent pathway. Phytomedicine 2018, 49, 41–51. [Google Scholar] [CrossRef]
- Ji, N.; Jiang, L.; Deng, P.; Xu, H.; Chen, F.; Liu, J.; Li, J.; Liao, G.; Zeng, X.; Lin, Y.; et al. Synergistic effect of honokiol and 5-fluorouracil on apoptosis of oral squamous cell carcinoma cells. J. Oral Pathol. Med. 2017, 46, 201–207. [Google Scholar] [CrossRef]
- Yi, X.; Lou, L.; Wang, J.; Xiong, J.; Zhou, S. Honokiol antagonizes doxorubicin resistance in human breast cancer via miR-188-5p/FBXW7/c-Myc pathway. Cancer Chemother. Pharmacol. 2021, 87, 647–656. [Google Scholar] [CrossRef]
- Wang, C.; Cai, Z.; Huang, Y.; Liu, X.; Liu, X.; Chen, F.; Li, W. Honokiol in glioblastoma recurrence: A case report. Front. Neurol. 2023, 14, 1172860. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.P.; Liao, M.; Dai, C.; Chen, J.F.; Yang, C.J.; Liu, M.; Chen, Z.G.; Yao, M.C. Sanguisorba officinalis L synergistically enhanced 5-fluorouracil cytotoxicity in colorectal cancer cells by promoting a reactive oxygen species-mediated, mitochondria-caspase-dependent apoptotic pathway. Sci. Rep. 2016, 6, 34245. [Google Scholar] [CrossRef]
- Liu, J.; Zu, M.; Chen, K.; Gao, L.; Min, H.; Zhuo, W.; Chen, W.; Liu, A. Screening of neuraminidase inhibitory activities of some medicinal plants traditionally used in Lingnan Chinese medicines. BMC Complement. Altern. Med. 2018, 18, 102. [Google Scholar] [CrossRef]
- Wang, Z.; Loo, W.T.; Wang, N.; Chow, L.W.; Wang, D.; Han, F.; Zheng, X.; Chen, J.P. Effect of Sanguisorba officinalis L on breast cancer growth and angiogenesis. Expert. Opin. Ther. Targets 2012, 16 (Suppl. S1), 79–89. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qian, C.; Zhang, W.; Shi, M.; Chen, X.; Wang, Y.; Lin, F. Ginkgetin suppresses ovarian cancer growth through inhibition of JAK2/STAT3 and MAPKs signaling pathways. Phytomedicine 2023, 116, 154846. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Jung, S.N.; Yun, J.; Lee, C.W.; Choi, J.; Lee, Y.J.; Han, D.C.; Kwon, B.M. Ginkgetin inhibits the growth of DU-145 prostate cancer cells through inhibition of signal transducer and activator of transcription 3 activity. Cancer Sci. 2015, 106, 413–420. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Z.; Zhang, G.; Wang, T.; Ma, Y.; Guo, W. Down-regulation of PADI2 prevents proliferation and epithelial-mesenchymal transition in ovarian cancer through inhibiting JAK2/STAT3 pathway in vitro and in vivo, alone or in combination with Olaparib. J. Transl. Med. 2020, 18, 357. [Google Scholar] [CrossRef]
- Lou, J.S.; Zhao, L.P.; Huang, Z.H.; Chen, X.Y.; Xu, J.T.; Tai, W.C.; Tsim, K.W.K.; Chen, Y.T.; Xie, T. Ginkgetin derived from Ginkgo biloba leaves enhances the therapeutic effect of cisplatin via ferroptosis-mediated disruption of the Nrf2/HO-1 axis in EGFR wild-type non-small-cell lung cancer. Phytomedicine 2021, 80, 153370. [Google Scholar] [CrossRef]
- Miki, K.; Nagai, T.; Suzuki, K.; Tsujimura, R.; Koyama, K.; Kinoshita, K.; Furuhata, K.; Yamada, H.; Takahashi, K. Anti-influenza virus activity of biflavonoids. Bioorg. Med. Chem. Lett. 2007, 17, 772–775. [Google Scholar] [CrossRef]
- Mykhailenko, O.; Petrikaitė, V.; Korinek, M.; El-Shazly, M.; Chen, B.H.; Yen, C.H.; Hsieh, C.F.; Bezruk, I.; Dabrišiūtė, A.; Ivanauskas, L.; et al. Bio-guided bioactive profiling and HPLC-DAD fingerprinting of Ukrainian saffron (Crocus sativus stigmas): Moving from correlation toward causation. BMC Complement. Med. Ther. 2021, 21, 203. [Google Scholar] [CrossRef]
- Huang, Y.; Guo, X.; Wang, Z.; Yin, C.; Chen, M.; Xie, J.; Li, N.; Tu, Z.; Li, J.; Cao, J.; et al. High-resolution neuraminidase inhibition profiling of Arnebia euchroma (Royle) I.M. Johnst. based on HR-MS and target isolation: An example study of anti-infectious constituents in traditional Chinese medicine. J. Ethnopharmacol. 2023, 319, 117074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Chen, Q.Q.; Li, J.; Zhang, L.; Qi, L.W. Neuraminidase 1 and its Inhibitors from Chinese Herbal Medicines: An Emerging Role for Cardiovascular Diseases. Am. J. Chin. Med. 2021, 49, 843–862. [Google Scholar] [CrossRef] [PubMed]
- Chavas, L.M.; Kato, R.; Suzuki, N.; von Itzstein, M.; Mann, M.C.; Thomson, R.J.; Dyason, J.C.; McKimm-Breschkin, J.; Fusi, P.; Tringali, C.; et al. Complexity in influenza virus targeted drug design: Interaction with human sialidases. J. Med. Chem. 2010, 53, 2998–3002. [Google Scholar] [CrossRef]
- Liu, S.Q.; Xu, C.Y.; Qin, M.B.; Tan, L.; Zhuge, C.F.; Mao, Y.B.; Lai, M.Y.; Huang, J.A. Ginkgo biloba extract enhances chemotherapy sensitivity and reverses chemoresistance through suppression of the KSR1-mediated ERK1/2 pathway in gastric cancer cells. Oncol. Rep. 2015, 33, 2871–2882. [Google Scholar] [CrossRef]
- Yue, J.; Zhang, S.; Zheng, B.; Raza, F.; Luo, Z.; Li, X.; Zhang, Y.; Nie, Q.; Qiu, M. Efficacy and Mechanism of Active Fractions in Fruit of Amomum villosum Lour. for Gastric Cancer. J. Cancer 2021, 12, 5991–5998. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Shu, Y.; Hu, X.; Akram, W.; Wang, J.; Dong, S.; Luo, B.; Zhang, J.; Hu, S.; Li, X.; et al. An Optimized Two-Herb Chinese Food as Medicine Formula Reduces Cisplatin-Induced Nephrotoxicity in the Treatment of Lung Cancer in Mice. Front. Pharmacol. 2022, 13, 827901. [Google Scholar] [CrossRef]
- Dagsuyu, E.; Koroglu, P.; Gul, I.B.; Bulan, O.K.; Yanardag, R. Oxidative brain and cerebellum injury in diabetes and prostate cancer model: Protective effect of metformin. J. Biochem. Mol. Toxicol. 2023, 37, e23440. [Google Scholar] [CrossRef]
- Wang, D.; Bie, L.; Su, Y.; Xu, H.; Zhang, F.; Su, Y.; Zhang, B. Effect of lithocholic acid on biologically active α,β-unsaturated aldehydes induced by H2O2 in glioma mitochondria for use in glioma treatment. Int. J. Mol. Med. 2018, 41, 3195–3202. [Google Scholar] [CrossRef]
- Trah, J.; Arand, J.; Oh, J.; Pagerols-Raluy, L.; Trochimiuk, M.; Appl, B.; Heidelbach, H.; Vincent, D.; Saleem, M.A.; Reinshagen, K.; et al. Lithocholic bile acid induces apoptosis in human nephroblastoma cells: A non-selective treatment option. Sci. Rep. 2020, 10, 20349. [Google Scholar] [CrossRef]
- Schnaar, R.L.; Gerardy-Schahn, R.; Hildebrandt, H. Sialic acids in the brain: Gangliosides and polysialic acid in nervous system development, stability, disease, and regeneration. Physiol. Rev. 2014, 94, 461–518. [Google Scholar] [CrossRef]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Schnaar, R.L. Glycolipid-mediated cell-cell recognition in inflammation and nerve regeneration. Arch. Biochem. Biophys. 2004, 426, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Mikami, T.; Kashiwagi, M.; Tsuchihashi, K.; Daino, T.; Akino, T.; Gasa, S. Further characterization of equine brain gangliosides: The presence of GM3 having N-glycolyl neuraminic acid in the central nervous system. J. Biochem. 1998, 123, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Fabris, D.; Rožman, M.; Sajko, T.; Vukelić, Ž. Aberrant ganglioside composition in glioblastoma multiforme and peritumoral tissue: A mass spectrometry characterization. Biochimie 2017, 137, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Sarbu, M.; Petrica, L.; Clemmer, D.E.; Vukelić, Ž.; Zamfir, A.D. Gangliosides of human glioblastoma multiforme: A comprehensive mapping and structural analysis by ion mobility tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2021, 32, 1249–1257. [Google Scholar] [CrossRef]
- El Malki, K.; Wehling, P.; Alt, F.; Sandhoff, R.; Zahnreich, S.; Ustjanzew, A.; Wilzius, C.; Brockmann, M.A.; Wingerter, A.; Russo, A.; et al. Glucosylceramide Synthase Inhibitors Induce Ceramide Accumulation and Sensitize H3K27 Mutant Diffuse Midline Glioma to Irradiation. Int. J. Mol. Sci. 2023, 24, 9905. [Google Scholar] [CrossRef]
- Inda, M.M.; Bonavia, R.; Seoane, J. Glioblastoma multiforme: A look inside its heterogeneous nature. Cancers 2014, 6, 226–239. [Google Scholar] [CrossRef]
- Becker, A.P.; Sells, B.E.; Haque, S.J.; Chakravarti, A. Tumor Heterogeneity in Glioblastomas: From Light Microscopy to Molecular Pathology. Cancers 2021, 13, 761. [Google Scholar] [CrossRef]
- Wielgat, P.; Wawrusiewicz-Kurylonek, N.; Czarnomysy, R.; Rogowski, K.; Bielawski, K.; Car, H. The Paired Siglecs in Brain Tumours Therapy: The Immunomodulatory Effect of Dexamethasone and Temozolomide in Human Glioma In Vitro Model. Int. J. Mol. Sci. 2021, 22, 1791. [Google Scholar] [CrossRef]
- Hsu, C.C.; Lin, T.W.; Chang, W.W.; Wu, C.Y.; Lo, W.H.; Wang, P.H.; Tsai, Y.C. Soyasaponin-I-modified invasive behavior of cancer by changing cell surface sialic acids. Gynecol. Oncol. 2005, 96, 415–422. [Google Scholar] [CrossRef]
- Montgomery, A.; Szabo, R.; Skropeta, D.; Yu, H. Computational characterisation of the interactions between human ST6Gal I and transition-state analogue inhibitors: Insights for inhibitor design. J. Mol. Recognit. 2016, 29, 210–222. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Target | Biochemical Effect | Other Effects | Cancer Type | |
|---|---|---|---|---|---|
| Neuraminidase Inhibitors | Oseltamivir Zanamivir | Inhibit: ►Virus neuraminidase ►Human sialidase NEU1 NEU2 | Altered sialylation pattern | Reversal of EMT; increased drug sensitivity | Lung cancer (oseltamivir) [166] |
| Honokiol (HNK) | Regulate the expression of Neu1 Suppress Neu1 protein expression | Anti-tumor, anti-inflammatory, antioxidant, and antiangiogenic activity, enhancing the proapoptotic effects of drugs | Breast cancer (MCF, MDA) [143,144,145,146,147,148,149,150,151,152] | ||
| Sanguisorba officinalis | Neuraminidase activity | Sanguisorba officinalis + 5FU → synergistically enhancing cytotoxicity | Colorectal cancer [153] | ||
| Ginkgo biloba (ginkgetin) | Viral sialidases | Anti-tumor activity, inhibited tumor growth, enhancing chemotherapy sensitivity, and reversing chemoresistance | Ovarian cancer cell lines, prostate cancer cells (DU-145), and gastric cancer cells [156,157,158,159,160,165] | ||
| Crocus sativus | Neuraminidase | Cytotoxic activity on cancer cells | Melanoma cells (IGR39), triple-negative breast cancer(MDA-MB-231), and glioblastoma cell lines (U-87) [161] | ||
| Amomum villosum, Melaphis chinensis (Galla Chinensis), Caryophylli Flos, Lonicera Japonica, Yupingfeng San, Huanglian Jiedu | Neuraminidase and probably others | Inhibitory effects on tumor growth, synergizing the antitumor effect of DDP (Lonicera Japonica) | Various types of cancer, gastric cancer, and lung cancer [166,167] | ||
| Drug | Target | Biochemical Effect | Other Effects | Cancer Type | |
|---|---|---|---|---|---|
| Antimetabolites | 3-dezauridine (competitive inhibitor of CTP synthetase) | Limiting resialylation without affecting de novo synthesis | Decreased sialic acid expression on cell surface | Diverse and substance-dependent | Various types of cancers [109] |
| Acivicin | Limiting resialylation and de novo sialic acid synthesis delay | ||||
| 1-beta-D-Arabinofuranosylcytosine | Inhibited both de novo sialic acid synthesis and membrane resialylation | ||||
| Racotumomab | NGc-containing gangliosides trigger the immune system against tumor antigen NGcGM3(mimics NGc gangliosides) | Decrease in sialic acid levels | Additive antitumor effect | Non-small-cell lung cancer model [105,106] | |
| Aspirin and celecoxib | Inhibit Neu-1 activity | Decrease in sialic acid levels | Inducing apoptosis and necrosis | Pancreatic cancer cells [113,114] | |
| Metformin and rosiglitazone | N/A | Higher serum sialic acid concentrations | Anti-tumor activity | Prostate cancer model [168] | |
| Soyasaponin I | ST3Gal-IV | Depress mRNA expression of ST3Gal-IV and attenuate α2,3-sialylation on the cell surface | Inhibit the migration ability of cancer cells, enhance cell adhesion to extracellular matrix proteins, and reduce metastasis | Breast cancer cells and lung cancer (mice) [108,181] | |
| Lithocholic acid and its derivatives | α2,3-sialyltransferase (ST3Gal-I) | ↓ sialic acid levels | Induces apoptosis and reduces cancer cell proliferation, aggressiveness, and the metastatic potential of primary tumors | Glioblastoma [168], Ewing sarcoma [169] and breast cancer [168,169,170,171] | |
| Transition-state analogs | Sialyltransferase, mimics the substrate CMP-Neu5Ac | The highest inhibitory activity and lower level of sialic acid expression on the cell surface | Potential new antimetastatic agents | Various types of cancers [182] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jastrząb, P.; Narejko, K.; Car, H.; Wielgat, P. Cell Membrane Sialome: Sialic Acids as Therapeutic Targets and Regulators of Drug Resistance in Human Cancer Management. Cancers 2023, 15, 5103. https://doi.org/10.3390/cancers15205103
Jastrząb P, Narejko K, Car H, Wielgat P. Cell Membrane Sialome: Sialic Acids as Therapeutic Targets and Regulators of Drug Resistance in Human Cancer Management. Cancers. 2023; 15(20):5103. https://doi.org/10.3390/cancers15205103
Chicago/Turabian StyleJastrząb, Patrycja, Karolina Narejko, Halina Car, and Przemyslaw Wielgat. 2023. "Cell Membrane Sialome: Sialic Acids as Therapeutic Targets and Regulators of Drug Resistance in Human Cancer Management" Cancers 15, no. 20: 5103. https://doi.org/10.3390/cancers15205103
APA StyleJastrząb, P., Narejko, K., Car, H., & Wielgat, P. (2023). Cell Membrane Sialome: Sialic Acids as Therapeutic Targets and Regulators of Drug Resistance in Human Cancer Management. Cancers, 15(20), 5103. https://doi.org/10.3390/cancers15205103