

Prevention and Treatment of HPV-Induced Skin Tumors


{kind=link}
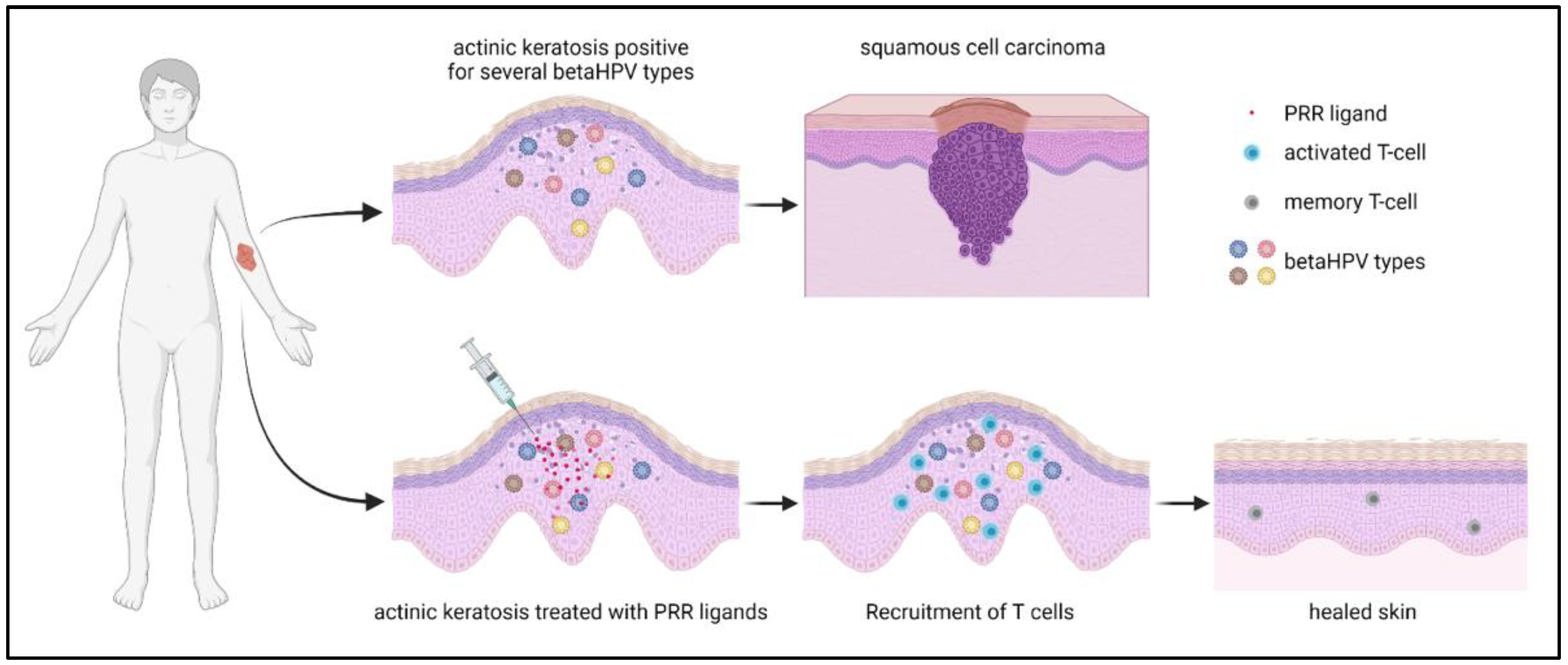{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Economic Burden of cSCC
3. The Impact of Immune Suppression
4. Area of Concern: What Are the “High-Risk” Cutaneous HPV Types?
5. Area of Concern: When Should a Prophylactic Vaccine Be Administered?

6. Prophylactic Vaccine Strategies in Clinical Trials
7. T-Cell Mediated Approaches against HPV-Induced Lesions
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kricker, A.; Weber, M.; Sitas, F.; Banks, E.; Rahman, B.; Goumas, C.; Kabir, A.; Hodgkinson, V.S.; van Kemenade, C.H.; Waterboer, T.; et al. Early Life UV and Risk of Basal and Squamous Cell Carcinoma in New South Wales, Australia. Photochem. Photobiol. 2017, 93, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Van Doorslaer, K.; Li, Z.; Xirasagar, S.; Maes, P.; Kaminsky, D.; Liou, D.; Sun, Q.; Kaur, R.; Huyen, Y.; McBride, A.A. The Papillomavirus Episteme: A major update to the papillomavirus sequence database. Nucleic. Acids Res. 2017, 45, D499–D506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Fera, A.N.; Warburton, A.; Coursey, T.L.; Khurana, S.; McBride, A.A. Persistent Human Papillomavirus Infection. Viruses 2021, 13, 321. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25 (Suppl. 1), 2–23. [Google Scholar] [CrossRef] [Green Version]
- Schiffman, M.; Doorbar, J.; Wentzensen, N.; de Sanjose, S.; Fakhry, C.; Monk, B.J.; Stanley, M.A.; Franceschi, S. Carcinogenic human papillomavirus infection. Nat. Rev. Dis. Primers 2016, 2, 16086. [Google Scholar] [CrossRef]
- Palefsky, J.M.; Lensing, S.Y.; Belzer, M.; Lee, J.; Gaur, A.H.; Mayer, K.; Futterman, D.; Stier, E.A.; Paul, M.E.; Chiao, E.Y.; et al. High prevalence of anal high-grade squamous intraepithelial lesions, and prevention through human papillomavirus vaccination, in young men who have sex with men living with HIV. Clin. Infect. Dis. 2021, 73, 1388–1396. [Google Scholar] [CrossRef]
- Hübbers, C.U.; Akgül, B. HPV and cancer of the oral cavity. Virulence 2015, 6, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Hess, J. Predictive Factors for Outcome and Quality of Life in HPV-Positive and HPV-Negative HNSCC. Recent Results Cancer Res. 2017, 206, 233–242. [Google Scholar] [CrossRef]
- Wurlitzer, M.; Mockelmann, N.; Kriegs, M.; Vens, M.; Omidi, M.; Hoffer, K.; Bargen, C.V.; Moller-Koop, C.; Witt, M.; Droste, C.; et al. Mass Spectrometric Comparison of HPV-Positive and HPV-Negative Oropharyngeal Cancer. Cancers 2020, 12, 1531. [Google Scholar] [CrossRef]
- De Jong, S.J.; Imahorn, E.; Itin, P.; Uitto, J.; Orth, G.; Jouanguy, E.; Casanova, J.L.; Burger, B. Epidermodysplasia Verruciformis: Inborn Errors of Immunity to Human Beta-Papillomaviruses. Front. Microbiol. 2018, 9, 1222. [Google Scholar] [CrossRef]
- Venuti, A.; Lohse, S.; Tommasino, M.; Smola, S. Cross-talk of cutaneous beta human papillomaviruses and the immune system: Determinants of disease penetrance. Philos Trans. R Soc. Lond B Biol. Sci. 2019, 374, 20180287. [Google Scholar] [CrossRef] [Green Version]
- Smola, S. Human Papillomaviruses and Skin Cancer. Adv. Exp. Med. Biol. 2020, 1268, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Howley, P.M.; Pfister, H.J. Beta genus papillomaviruses and skin cancer. Virology 2015, 479–480, 290–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tampa, M.; Mitran, C.I.; Mitran, M.I.; Nicolae, I.; Dumitru, A.; Matei, C.; Manolescu, L.; Popa, G.L.; Caruntu, C.; Georgescu, S.R. The Role of Beta HPV Types and HPV-Associated Inflammatory Processes in Cutaneous Squamous Cell Carcinoma. J. Immunol. Res. 2020, 2020, 5701639. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, W.R.; He, Q.; Rady, P.L.; Hughes, T.K.; Neto, C.F.; Rivitti, E.A.; Tyring, S.K. HPV typing in Brazilian patients with epidermodysplasia verruciformis: High prevalence of EV-HPV 25. J. Cutan Med. Surg. 2004, 8, 110–115. [Google Scholar] [CrossRef]
- Dell’Oste, V.; Azzimonti, B.; De Andrea, M.; Mondini, M.; Zavattaro, E.; Leigheb, G.; Weissenborn, S.J.; Pfister, H.; Michael, K.M.; Waterboer, T.; et al. High beta-HPV DNA loads and strong seroreactivity are present in epidermodysplasia verruciformis. J. Invest. Dermatol. 2009, 129, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Bouwes Bavinck, J.N.; Feltkamp, M.C.W.; Green, A.C.; Fiocco, M.; Euvrard, S.; Harwood, C.A.; Nasir, S.; Thomson, J.; Proby, C.M.; Naldi, L.; et al. Human papillomavirus and posttransplantation cutaneous squamous cell carcinoma: A multicenter, prospective cohort study. Am. J. Transplant. 2018, 18, 1220–1230. [Google Scholar] [CrossRef] [Green Version]
- Hasche, D.; Vinzon, S.E.; Rosl, F. Cutaneous Papillomaviruses and Non-melanoma Skin Cancer: Causal Agents or Innocent Bystanders? Front. Microbiol. 2018, 9, 874. [Google Scholar] [CrossRef] [Green Version]
- Rollison, D.E.; Viarisio, D.; Amorrortu, R.P.; Gheit, T.; Tommasino, M. An emerging issue in oncogenic virology: The role of beta HPV types in development of cutaneous squamous cell carcinoma. J. Virol. 2019, 93, e01003-18. [Google Scholar] [CrossRef] [Green Version]
- Tommasino, M. HPV and skin carcinogenesis. Papillomavirus Res. 2019, 7, 129–131. [Google Scholar] [CrossRef]
- Hufbauer, M.; Akgül, B. Molecular Mechanisms of Human Papillomavirus Induced Skin Carcinogenesis. Viruses 2017, 9, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendel, S.O.; Wallace, N.A. Loss of Genome Fidelity: Beta HPVs and the DNA Damage Response. Front. Microbiol. 2017, 8, 2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gheit, T. Mucosal and Cutaneous Human Papillomavirus Infections and Cancer Biology. Front. Oncol. 2019, 9, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Que, S.K.T.; Zwald, F.O.; Schmults, C.D. Cutaneous squamous cell carcinoma: Incidence, risk factors, diagnosis, and staging. J. Am. Acad. Dermatol. 2018, 78, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Augustin, J.; Kis, A.; Sorbe, C.; Schafer, I.; Augustin, M. Epidemiology of skin cancer in the German population: Impact of socioeconomic and geographic factors. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1906–1913. [Google Scholar] [CrossRef]
- Augustin, A.; Glaeske, G. Hautkrebsreport 2019; Universität Bremen: Bremen, Germany, 2019; Available online: https://www.tk.de/resource/blob/2061220/6fc0df4b35fa9776c1699ad71a0c4542/hautkrebsreport-2019-data.pdf (accessed on 17 February 2023).
- Tokez, S.; Hollestein, L.; Louwman, M.; Nijsten, T.; Wakkee, M. Incidence of Multiple vs First Cutaneous Squamous Cell Carcinoma on a Nationwide Scale and Estimation of Future Incidences of Cutaneous Squamous Cell Carcinoma. JAMA Dermatol. 2020, 156, 1300–1306. [Google Scholar] [CrossRef]
- Tang, E.; Fung, K.; Chan, A.W. Incidence and mortality rates of keratinocyte carcinoma from 1998–2017: A population-based study of sex differences in Ontario, Canada. CMAJ 2021, 193, E1516–E1524. [Google Scholar] [CrossRef]
- Umezono, Y.; Sato, Y.; Noto, M.; Yamada, K.; Noguchi, N.; Hasunuma, N.; Osada, S.I.; Manabe, M. Incidence rate of cutaneous squamous cell carcinoma is rapidly increasing in Akita Prefecture: Urgent alert for super-aged society. J. Dermatol. 2019, 46, 259–262. [Google Scholar] [CrossRef]
- Gordon, L.G.; Elliott, T.M.; Wright, C.Y.; Deghaye, N.; Visser, W. Modelling the healthcare costs of skin cancer in South Africa. BMC Health Serv. Res. 2016, 16, 113. [Google Scholar] [CrossRef] [Green Version]
- Imarc. Non-Melanoma Skin Cancer Treatment Market: Global Industry Trends, Share, Size, Growth, Opportunity and Forecast 2022–2027. 2021. Available online: https://www.imarcgroup.com/non-melanoma-skin-cancer-treatment-market (accessed on 17 February 2023).
- Mofidi, A.; Tompa, E.; Spencer, J.; Kalcevich, C.; Peters, C.E.; Kim, J.; Song, C.; Mortazavi, S.B.; Demers, P.A. The economic burden of occupational non-melanoma skin cancer due to solar radiation. J. Occup. Environ. Hyg. 2018, 15, 481–491. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, R.; Knusel, K.D.; Ezaldein, H.H.; Bordeaux, J.S.; Scott, J.F. Characteristics of Patients Hospitalized for Cutaneous Squamous Cell Carcinoma. Dermatol. Surg. 2020, 46, 742–746. [Google Scholar] [CrossRef] [PubMed]
- Marcellusi, A.; Bini, C.; Peris, K.; Ascierto, P.A.; Mennini, F.S. Cost of illness of cutaneous squamous cell carcinoma (CSCC). Glob. Reg. Health Technol. Assess 2020, 7, 148–153. [Google Scholar] [CrossRef]
- Aguilar-Bernier, M.; Gonzalez-Carrascosa, M.; Padilla-Espana, L.; Rivas-Ruiz, F.; Jimenez-Puente, A.; de Troya-Martin, M. Five-year economic evaluation of non-melanoma skin cancer surgery at the Costa del Sol Hospital (2006–2010). J. Eur. Acad. Dermatol. Venereol. 2014, 28, 320–326. [Google Scholar] [CrossRef]
- Fransen, M.; Karahalios, A.; Sharma, N.; English, D.R.; Giles, G.G.; Sinclair, R.D. Non-melanoma skin cancer in Australia. Med. J Aust. 2012, 197, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Euvrard, S.; Kanitakis, J.; Claudy, A. Skin cancers after organ transplantation. N. Engl. J. Med. 2003, 348, 1681–1691. [Google Scholar] [CrossRef] [Green Version]
- Spurgeon, M.E.; Lambert, P.F. Mus musculus Papillomavirus 1: A New Frontier in Animal Models of Papillomavirus Pathogenesis. J. Virol. 2020, 94, e00002-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickley, J.D.; Messerschmidt, J.L.; Awad, M.E.; Li, T.; Hasegawa, T.; Ha, D.T.; Nabeta, H.W.; Bevins, P.A.; Ngo, K.H.; Asgari, M.M.; et al. Immunity to commensal papillomaviruses protects against skin cancer. Nature 2019, 575, 519–522. [Google Scholar] [CrossRef]
- Lambert, P.F.; Munger, K.; Rosl, F.; Hasche, D.; Tommasino, M. Beta human papillomaviruses and skin cancer. Nature 2020, 588, E20–E21. [Google Scholar] [CrossRef]
- Rangwala, S.; Tsai, K.Y. Roles of the immune system in skin cancer. Br. J. Dermatol. 2011, 165, 953–965. [Google Scholar] [CrossRef] [Green Version]
- Neale, R.E.; Weissenborn, S.; Abeni, D.; Bavinck, J.N.; Euvrard, S.; Feltkamp, M.C.; Green, A.C.; Harwood, C.; de Koning, M.; Naldi, L.; et al. Human papillomavirus load in eyebrow hair follicles and risk of cutaneous squamous cell carcinoma. Cancer Epidemiol. Biomarkers Prev. 2013, 22, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Antonsson, A.; Erfurt, C.; Hazard, K.; Holmgren, V.; Simon, M.; Kataoka, A.; Hossain, S.; Hakangard, C.; Hansson, B.G. Prevalence and type spectrum of human papillomaviruses in healthy skin samples collected in three continents. J. Gen. Virol. 2003, 84, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Galati, L.; Brancaccio, R.N.; Robitaille, A.; Cuenin, C.; Luzi, F.; Fiorucci, G.; Chiantore, M.V.; Marascio, N.; Matera, G.; Liberto, M.C.; et al. Detection of human papillomaviruses in paired healthy skin and actinic keratosis by next generation sequencing. Papillomavirus Res. 2020, 9, 100196. [Google Scholar] [CrossRef] [PubMed]
- Weissenborn, S.; Neale, R.E.; Waterboer, T.; Abeni, D.; Bavinck, J.N.; Green, A.C.; Harwood, C.A.; Euvrard, S.; Feltkamp, M.C.; de Koning, M.N.; et al. Beta-papillomavirus DNA loads in hair follicles of immunocompetent people and organ transplant recipients. Med. Microbiol. Immunol. 2012, 201, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Lott, D.G.; Manz, R.; Koch, C.; Lorenz, R.R. Aggressive behavior of nonmelanotic skin cancers in solid organ transplant recipients. Transplantation 2010, 90, 683–687. [Google Scholar] [CrossRef]
- Alam, M.; Ratner, D. Cutaneous squamous-cell carcinoma. N. Engl. J. Med. 2001, 344, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Weissenborn, S.J.; Nindl, I.; Purdie, K.; Harwood, C.; Proby, C.; Breuer, J.; Majewski, S.; Pfister, H.; Wieland, U. Human papillomavirus-DNA loads in actinic keratoses exceed those in non-melanoma skin cancers. J. Investig. Dermatol. 2005, 125, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hufbauer, M.; Lazic, D.; Akgül, B.; Brandsma, J.L.; Pfister, H.; Weissenborn, S.J. Enhanced human papillomavirus type 8 oncogene expression levels are crucial for skin tumorigenesis in transgenic mice. Virology 2010, 403, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Weissenborn, S.J.; De Koning, M.N.; Wieland, U.; Quint, W.G.; Pfister, H.J. Intrafamilial transmission and family-specific spectra of cutaneous betapapillomaviruses. J. Virol. 2009, 83, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Hsu, J.Y.; Chen, A.C.; Keleher, A.; McMillan, N.A.; Antonsson, A. Shared and persistent asymptomatic cutaneous human papillomavirus infections in healthy skin. J. Med. Virol. 2009, 81, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Hopfl, R.; Bens, G.; Wieland, U.; Petter, A.; Zelger, B.; Fritsch, P.; Pfister, H. Human papillomavirus DNA in non-melanoma skin cancers of a renal transplant recipient: Detection of a new sequence related to epidermodysplasia verruciformis associated types. J. Invest. Dermatol. 1997, 108, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Bens, G.; Wieland, U.; Hofmann, A.; Hopfl, R.; Pfister, H. Detection of new human papillomavirus sequences in skin lesions of a renal transplant recipient and characterization of one complete genome related to epidermodysplasia verruciformis-associated types. J. Gen. Virol. 1998, 79 Pt 4, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Villiers, E.M.; Lavergne, D.; McLaren, K.; Benton, E.C. Prevailing papillomavirus types in non-melanoma carcinomas of the skin in renal allograft recipients. Int. J. Cancer. 1997, 73, 356–361. [Google Scholar] [CrossRef]
- Iannacone, M.R.; Gheit, T.; Pfister, H.; Giuliano, A.R.; Messina, J.L.; Fenske, N.A.; Cherpelis, B.S.; Sondak, V.K.; Roetzheim, R.G.; Silling, S.; et al. Case-control study of genus-beta human papillomaviruses in plucked eyebrow hairs and cutaneous squamous cell carcinoma. Int. J. Cancer 2014, 134, 2231–2244. [Google Scholar] [CrossRef] [Green Version]
- Bolatti, E.M.; Hosnjak, L.; Chouhy, D.; Re-Louhau, M.F.; Casal, P.E.; Bottai, H.; Kocjan, B.J.; Stella, E.J.; Gorosito, M.D.; Sanchez, A.; et al. High prevalence of Gammapapillomaviruses (Gamma-PVs) in pre-malignant cutaneous lesions of immunocompetent individuals using a new broad-spectrum primer system, and identification of HPV210, a novel Gamma-PV type. Virology 2018, 525, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.T.; Muller, M. Next generation prophylactic human papillomavirus vaccines. Lancet. Oncol. 2015, 16, e217–e225. [Google Scholar] [CrossRef] [PubMed]
- Pogoda, C.S.; Roden, R.B.; Garcea, R.L. Immunizing against Anogenital Cancer: HPV Vaccines. PLoS Pathog. 2016, 12, e1005587. [Google Scholar] [CrossRef] [PubMed]
- Hasche, D.; Rosl, F. Mastomys Species as Model Systems for Infectious Diseases. Viruses 2019, 11, 182. [Google Scholar] [CrossRef] [Green Version]
- Vinzon, S.E.; Braspenning-Wesch, I.; Muller, M.; Geissler, E.K.; Nindl, I.; Grone, H.J.; Schafer, K.; Rosl, F. Protective vaccination against papillomavirus-induced skin tumors under immunocompetent and immunosuppressive conditions: A preclinical study using a natural outbred animal model. PLoS Pathog. 2014, 10, e1003924. [Google Scholar] [CrossRef]
- Fu, Y.; Cao, R.; Schafer, M.; Stephan, S.; Braspenning-Wesch, I.; Schmitt, L.; Bischoff, R.; Muller, M.; Schafer, K.; Vinzon, S.E.; et al. Expression of different L1 isoforms of Mastomys natalensis papillomavirus as mechanism to circumvent adaptive immunity. eLife 2020, 9, e57626. [Google Scholar] [CrossRef]
- Hasche, D.; Ahmels, M.; Braspenning-Wesch, I.; Stephan, S.; Cao, R.; Schmidt, G.; Muller, M.; Rosl, F. Isoforms of the Papillomavirus Major Capsid Protein Differ in Their Ability to Block Viral Spread and Tumor Formation. Front. Immunol. 2022, 13, 811094. [Google Scholar] [CrossRef]
- Ahmels, M.; Mariz, F.C.; Braspenning-Wesch, I.; Stephan, S.; Huber, B.; Schmidt, G.; Cao, R.; Muller, M.; Kirnbauer, R.; Rosl, F.; et al. Next generation L2-based HPV vaccines cross-protect against cutaneous papillomavirus infection and tumor development. Front. Immunol. 2022, 13, 1010790. [Google Scholar] [CrossRef] [PubMed]
- Pouyanfard, S.; Muller, M. Human papillomavirus first and second generation vaccines-current status and future directions. Biol. Chem. 2017, 398, 871–889. [Google Scholar] [CrossRef]
- Huber, B.; Wang, J.W.; Roden, R.B.S.; Kirnbauer, R. RG1-VLP and Other L2-Based, Broad-Spectrum HPV Vaccine Candidates. J. Clin. Med. 2021, 10, 1044. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Ishii, Y.; Ochi, H.; Matsumoto, T.; Yoshikawa, H.; Kanda, T. Neutralization of HPV16, 18, 31, and 58 pseudovirions with antisera induced by immunizing rabbits with synthetic peptides representing segments of the HPV16 minor capsid protein L2 surface region. Virology 2007, 358, 266–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellenbacher, C.; Kwak, K.; Fink, D.; Shafti-Keramat, S.; Huber, B.; Jindra, C.; Faust, H.; Dillner, J.; Roden, R.B.S.; Kirnbauer, R. Efficacy of RG1-VLP vaccination against infections with genital and cutaneous human papillomaviruses. J. Investig. Dermatol. 2013, 133, 2706–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellenbacher, C.; Roden, R.; Kirnbauer, R. Chimeric L1-L2 virus-like particles as potential broad-spectrum human papillomavirus vaccines. J. Virol. 2009, 83, 10085–10095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellenbacher, C.; Huber, B.; Skoll, M.; Shafti-Keramat, S.; Roden, R.B.S.; Kirnbauer, R. Incorporation of RG1 epitope into HPV16L1-VLP does not compromise L1-specific immunity. Vaccine 2019, 37, 3529–3534. [Google Scholar] [CrossRef]
- Olczak, P.; Matsui, K.; Wong, M.; Alvarez, J.; Lambert, P.; Christensen, N.D.; Hu, J.; Huber, B.; Kirnbauer, R.; Wang, J.W.; et al. RG2-VLP: A Vaccine Designed to Broadly Protect against Anogenital and Skin Human Papillomaviruses Causing Human Cancer. J. Virol. 2022, 96, e0056622. [Google Scholar] [CrossRef]
- Schellenbacher, C.; Roden, R.B.S.; Kirnbauer, R. Developments in L2-based human papillomavirus (HPV) vaccines. Virus Res. 2017, 231, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Gambhira, R.; Karanam, B.; Monie, A.; Hung, C.F.; Roden, R.; Wu, T.C. Generation and characterization of a preventive and therapeutic HPV DNA vaccine. Vaccine 2008, 26, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Jiang, R.T.; Wang, J.W.; Peng, S.; Huang, T.C.; Wang, C.; Cannella, F.; Chang, Y.N.; Viscidi, R.P.; Best, S.R.A.; Hung, C.F.; et al. Spontaneous and Vaccine-Induced Clearance of Mus Musculus Papillomavirus 1 Infection. J. Virol. 2017, 91, e00699-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, I.; Bolchi, A.; Moretto, N.; Canali, E.; Gissmann, L.; Tommasino, M.; Muller, M.; Ottonello, S. Potent anti-HPV immune responses induced by tandem repeats of the HPV16 L2 (20 -- 38) peptide displayed on bacterial thioredoxin. Vaccine 2009, 27, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.; Ribeiro-Muller, L.; Canali, E.; Bolchi, A.; Tommasino, M.; Ottonello, S.; Muller, M. Robust In Vitro and In Vivo Neutralization against Multiple High-Risk HPV Types Induced by a Thermostable Thioredoxin-L2 Vaccine. Cancer Prev. Res. 2015, 8, 932–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogun, S.A.; Dumon-Seignovert, L.; Marchand, J.B.; Holder, A.A.; Hill, F. The oligomerization domain of C4-binding protein (C4bp) acts as an adjuvant, and the fusion protein comprised of the 19-kilodalton merozoite surface protein 1 fused with the murine C4bp domain protects mice against malaria. Infect. Immun. 2008, 76, 3817–3823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouyanfard, S.; Spagnoli, G.; Bulli, L.; Balz, K.; Yang, F.; Odenwald, C.; Seitz, H.; Mariz, F.C.; Bolchi, A.; Ottonello, S.; et al. Minor Capsid Protein L2 Polytope Induces Broad Protection against Oncogenic and Mucosal Human Papillomaviruses. J. Virol. 2018, 92, e01930-17. [Google Scholar] [CrossRef] [Green Version]
- Mariz, F.C.; Balz, K.; Dittrich, M.; Zhang, Y.; Yang, F.; Zhao, X.; Bolchi, A.; Ottonello, S.; Muller, M. A broadly protective vaccine against cutaneous human papillomaviruses. NPJ Vaccines 2022, 7, 116. [Google Scholar] [CrossRef]
- Yoo, S.A.; Kim, Y.H.; Han, J.H.; Bang, C.H.; Park, Y.M.; Lee, J.H. Treatment of Actinic Keratosis: The Best Choice through an Observational Study. J. Clin. Med. 2022, 11, 3953. [Google Scholar] [CrossRef]
- Jansen, M.H.E.; Kessels, J.; Nelemans, P.J.; Kouloubis, N.; Arits, A.; van Pelt, H.P.A.; Quaedvlieg, P.J.F.; Essers, B.A.B.; Steijlen, P.M.; Kelleners-Smeets, N.W.J.; et al. Randomized Trial of Four Treatment Approaches for Actinic Keratosis. N. Engl. J. Med. 2019, 380, 935–946. [Google Scholar] [CrossRef]
- Vidal, D. Topical imiquimod: Mechanism of action and clinical applications. Mini. Rev. Med. Chem. 2006, 6, 499–503. [Google Scholar] [CrossRef]
- Gay, J.; Johnson, N.; Kavuru, V.; Phillips, M. Utility of the Human Papillomavirus Vaccination in Management of HPV-associated Cutaneous Lesions. Ski. Ther. Lett. 2021, 26, 6–8. [Google Scholar]
- Mo, Y.; Ma, J.; Zhang, H.; Shen, J.; Chen, J.; Hong, J.; Xu, Y.; Qian, C. Prophylactic and Therapeutic HPV Vaccines: Current Scenario and Perspectives. Front. Cell Infect. Microbiol. 2022, 12, 909223. [Google Scholar] [CrossRef] [PubMed]
- Schaper, I.D.; Marcuzzi, G.P.; Weissenborn, S.J.; Kasper, H.U.; Dries, V.; Smyth, N.; Fuchs, P.; Pfister, H. Development of skin tumors in mice transgenic for early genes of human papillomavirus type 8. Cancer Res. 2005, 65, 1394–1400. [Google Scholar] [CrossRef] [Green Version]
- Marcuzzi, G.P.; Hufbauer, M.; Kasper, H.U.; Weissenborn, S.J.; Smola, S.; Pfister, H. Spontaneous tumour development in human papillomavirus type 8 E6 transgenic mice and rapid induction by UV-light exposure and wounding. J. Gen. Virol. 2009, 90, 2855–2864. [Google Scholar] [CrossRef] [PubMed]
- Hufbauer, M.; Lazic, D.; Reinartz, M.; Akgül, B.; Pfister, H.; Weissenborn, S.J. Skin tumor formation in human papillomavirus 8 transgenic mice is associated with a deregulation of oncogenic miRNAs and their tumor suppressive targets. J. Dermatol. Sci. 2011, 64, 7–15. [Google Scholar] [CrossRef]
- Marcuzzi, G.P.; Awerkiew, S.; Hufbauer, M.; Schadlich, L.; Gissmann, L.; Eming, S.; Pfister, H. Tumor prevention in HPV8 transgenic mice by HPV8-E6 DNA vaccination. Med. Microbiol. Immunol. 2014, 203, 155–163. [Google Scholar] [CrossRef]
- Hufbauer, M.; Rattay, S.; Hagen, C.; Quaas, A.; Pfister, H.; Hartmann, G.; Coch, C.; Akgül, B. Poly(I:C) treatment prevents skin tumor formation in the preclinical HPV8 transgenic mouse model. J. Invest. Dermatol. 2022; in press. [Google Scholar] [CrossRef] [PubMed]
- Rattay, S.; Hufbauer, M.; Hagen, C.; Putschli, B.; Coch, C.; Akgül, B.; Hartmann, G. Human Beta Papillomavirus Type 8 E1 and E2 Proteins Suppress the Activation of the RIG-I-Like Receptor MDA5. Viruses 2022, 14, 1361. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasche, D.; Akgül, B. Prevention and Treatment of HPV-Induced Skin Tumors. Cancers 2023, 15, 1709. https://doi.org/10.3390/cancers15061709
Hasche D, Akgül B. Prevention and Treatment of HPV-Induced Skin Tumors. Cancers. 2023; 15(6):1709. https://doi.org/10.3390/cancers15061709
Chicago/Turabian StyleHasche, Daniel, and Baki Akgül. 2023. "Prevention and Treatment of HPV-Induced Skin Tumors" Cancers 15, no. 6: 1709. https://doi.org/10.3390/cancers15061709
APA StyleHasche, D., & Akgül, B. (2023). Prevention and Treatment of HPV-Induced Skin Tumors. Cancers, 15(6), 1709. https://doi.org/10.3390/cancers15061709