

TRPV6 Calcium Channel Targeting by Antibodies Raised against Extracellular Epitopes Induces Prostate Cancer Cell Apoptosis

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Electrophysiology and Solutions
2.3. Calcium Imaging
2.4. SDS-PAGE and Western Blotting
2.5. siRNA Transfection
2.6. Nucleofection
2.7. Cell Survival Assay
2.8. Cell Cycle Assay
2.9. TUNEL Assay
2.10. Immunocytochemistry
2.11. Plasmids
2.12. Antibody Production
2.13. Reagents
2.14. Data Analysis
3. Results
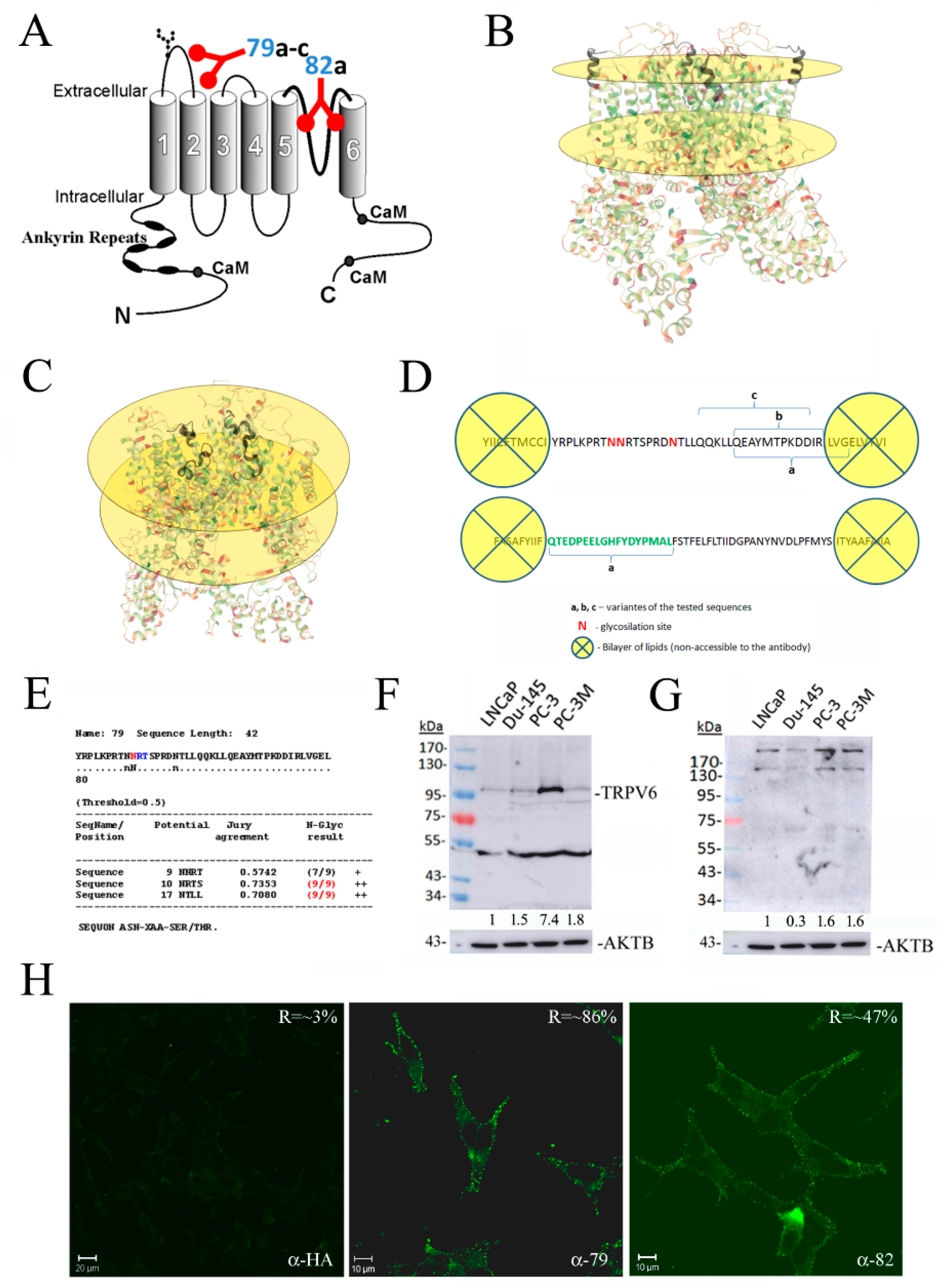
3.1. Design and Validation of Antibodies Raised against Extracellular Epitopes of the TRPV6 Protein
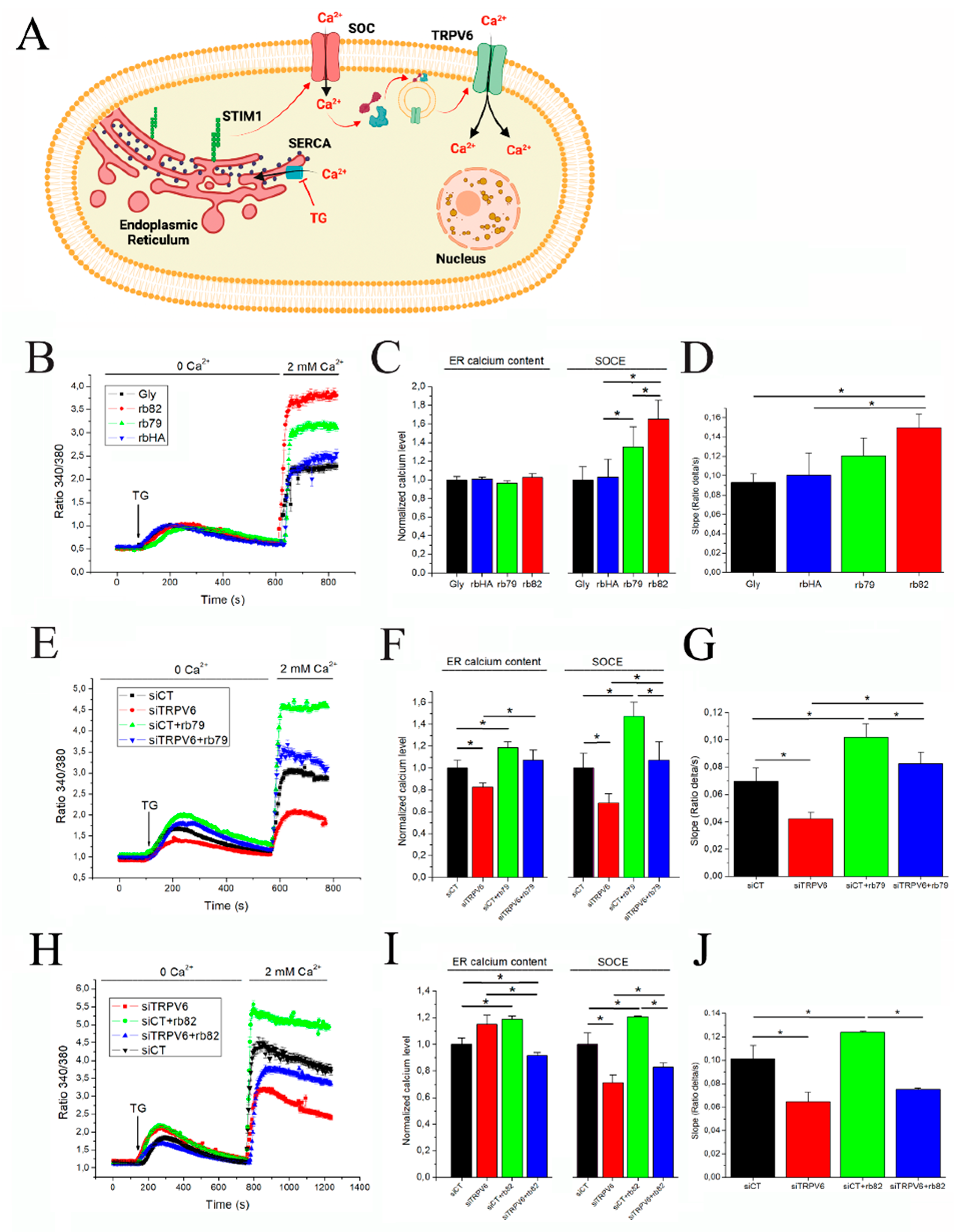
3.2. Antibody Treatments Increase Store-Operated Capacitive Calcium Entry in PCa Cells
3.3. Antibodies 79 and 82 Directly Affect TRPV6-Induced Currents
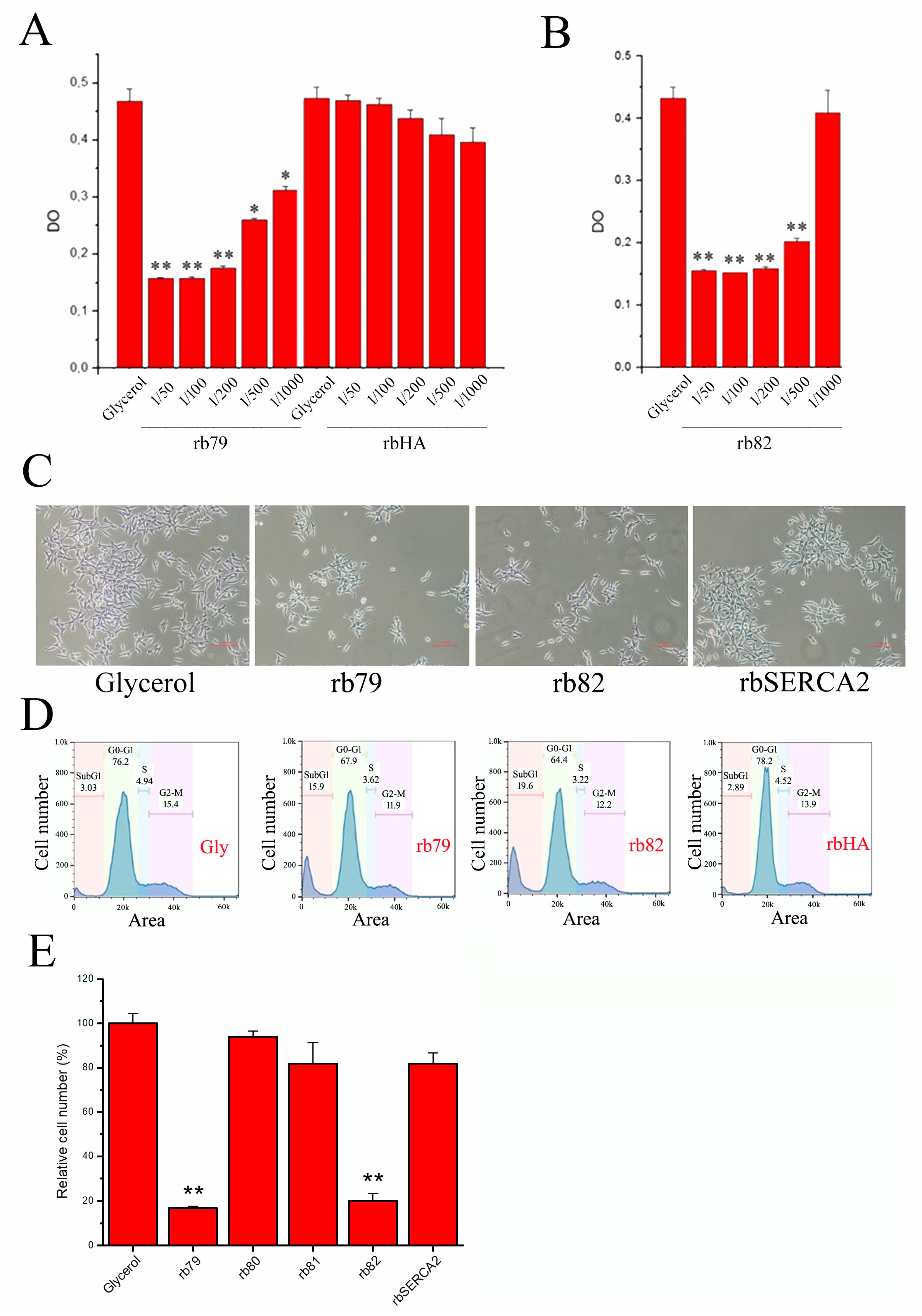
3.4. Decrease in Cell Survival via TRPV6 Channel Activation
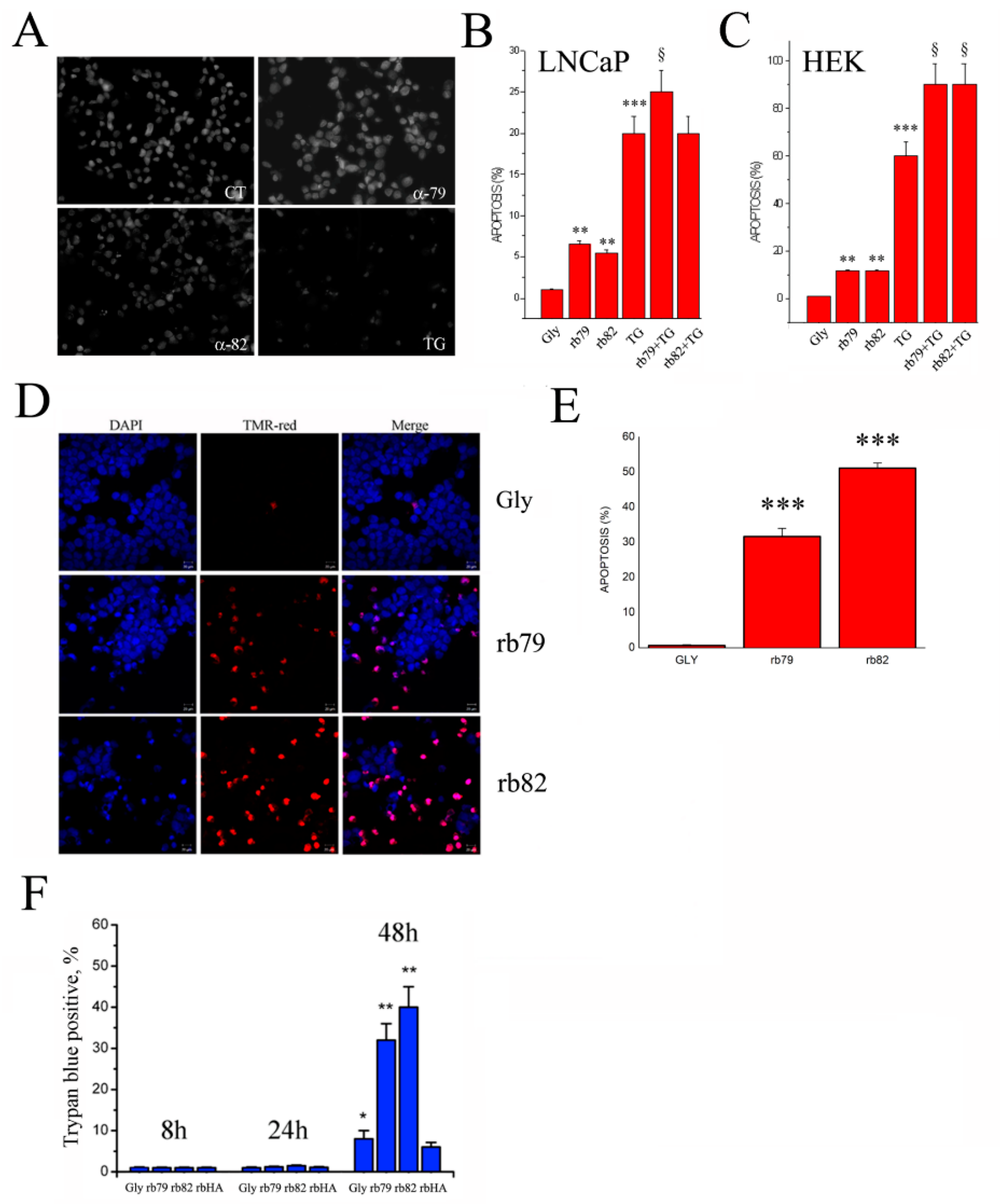
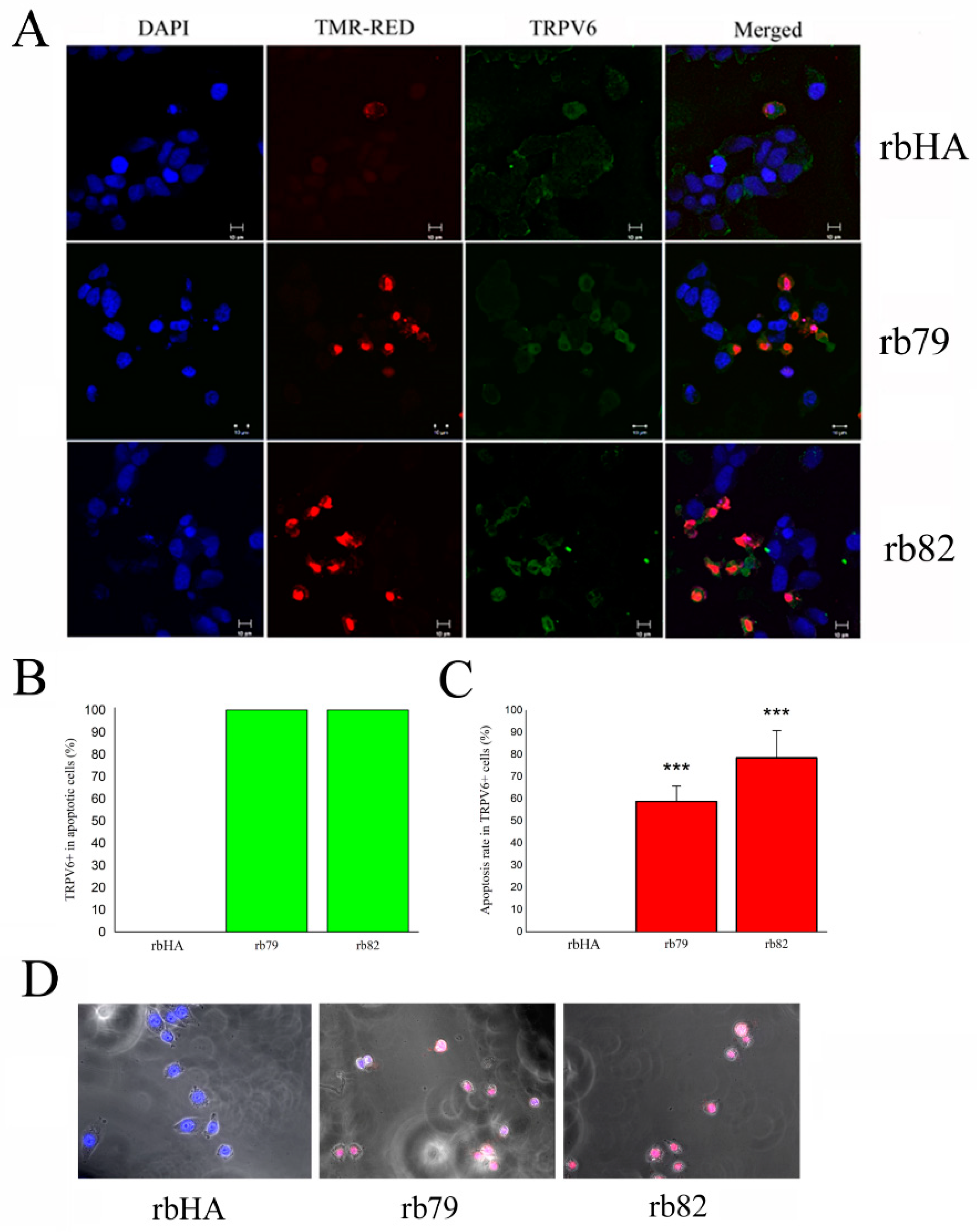
3.5. Plasma Membrane TRPV6 Expression Determinates Cancer Cell Fate
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rove, K.O.; Flaig, T.W. A renaissance in the medical treatment of advanced prostate cancer. Oncology 2010, 24, 1308–1313, 1318. [Google Scholar] [PubMed]
- Clapham, D.E.; Runnels, L.W.; Strübing, C. The trp ion channel family. Nat. Rev. Neurosci. 2001, 2, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Bödding, M.; Flockerzi, V. Ca2+ Dependence of the Ca2+-selective TRPV6 Channel. J. Biol. Chem. 2004, 279, 36546–36552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoenderop, J.G.J.; Nilius, B.; Bindels, R.J.M. Epithelial calcium channels: From identification to function and regulation. Pflügers Arch. 2003, 446, 304–308. [Google Scholar] [CrossRef]
- Dhennin-Duthille, I.; Gautier, M.; Faouzi, M.; Guilbert, A.; Brevet, M.; Vaudry, D.; Ahidouch, A.; Sevestre, H.; Ouadid-Ahidouch, H. High Expression of Transient Receptor Potential Channels in Human Breast Cancer Epithelial Cells and Tissues: Correlation with Pathological Parameters. Cell. Physiol. Biochem. 2011, 28, 813–822. [Google Scholar] [CrossRef]
- Lehen’Kyi, V.; Raphaël, M.; Prevarskaya, N. The role of the TRPV6 channel in cancer. J. Physiol. 2012, 590, 1369–1376. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.-B.; Zhuang, L.; Berger, U.V.; Adam, R.M.; Williams, B.; Brown, E.M.; Hediger, M.A.; Freeman, M.R. CaT1 Expression Correlates with Tumor Grade in Prostate Cancer. Biochem. Biophys. Res. Commun. 2001, 282, 729–734. [Google Scholar] [CrossRef]
- Zhuang, L.; Peng, J.-B.; Tou, L.; Takanaga, H.; Adam, R.M.; Hediger, M.A.; Freeman, M.R. Calcium-Selective Ion Channel, CaT1, Is Apically Localized in Gastrointestinal Tract Epithelia and Is Aberrantly Expressed in Human Malignancies. Lab. Investig. 2002, 82, 1755–1764. [Google Scholar] [CrossRef] [Green Version]
- Ouadid-Ahidouch, H.; Dhennin-Duthille, I.; Gautier, M.; Sevestre, H.; Ahidouch, A. TRP channels: Diagnostic markers and therapeutic targets for breast cancer? Trends Mol. Med. 2013, 19, 117–124. [Google Scholar] [CrossRef]
- Giona, S. The Epidemiology of Prostate Cancer. In Prostate Cancer; Bott, S.R., Ng, K.L., Eds.; Exon Publications: Brisbane, Australia, 2021; ISBN 978-0-645-00175-4. [Google Scholar]
- Westaby, D.; Jimenez-Vacas, J.M.; Padilha, A.; Varkaris, A.; Balk, S.P.; de Bono, J.S.; Sharp, A. Targeting the Intrinsic Apoptosis Pathway: A Window of Opportunity for Prostate Cancer. Cancers 2021, 14, 51. [Google Scholar] [CrossRef]
- Campbell, K.J.; Leung, H.Y. Evasion of cell death: A contributory factor in prostate cancer development and treatment resistance. Cancer Lett. 2021, 520, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Wissenbach, U.; Niemeyer, B.A.; Fixemer, T.; Schneidewind, A.; Trost, C.; Cavalié, A.; Reus, K.; Meese, E.; Bonkhoff, H.; Flockerzi, V. Expression of CaT-like, a Novel Calcium-selective Channel, Correlates with the Malignancy of Prostate Cancer. J. Biol. Chem. 2001, 276, 19461–19468. [Google Scholar] [CrossRef] [Green Version]
- Bowen, C.V.; DeBay, D.; Ewart, H.S.; Gallant, P.; Gormley, S.; Ilenchuk, T.T.; Iqbal, U.; Lutes, T.; Martina, M.; Mealing, G.; et al. In Vivo Detection of Human TRPV6-Rich Tumors with Anti-Cancer Peptides Derived from Soricidin. PLoS ONE 2013, 8, e58866. [Google Scholar] [CrossRef] [PubMed]
- Landowski, C.P.; Bolanz, K.A.; Suzuki, Y.; Hediger, M. Chemical Inhibitors of the Calcium Entry Channel TRPV6. Pharm. Res. 2010, 28, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Lehen’Kyi, V.; Flourakis, M.; Skryma, R.; Prevarskaya, N. TRPV6 channel controls prostate cancer cell proliferation via Ca2+/NFAT-dependent pathways. Oncogene 2007, 26, 7380–7385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haustrate, A.; Mihalache, A.; Cordier, C.; Gosset, P.; Prevarskaya, N.; Lehen’Kyi, V. A Novel Anti-TRPV6 Antibody and Its Application in Cancer Diagnosis In Vitro. Int. J. Mol. Sci. 2022, 24, 419. [Google Scholar] [CrossRef]
- Saotome, K.; Singh, A.K.; Yelshanskaya, M.V.; Sobolevsky, A.I. Crystal structure of the epithelial calcium channel TRPV6. Nature 2016, 534, 506–511. [Google Scholar] [CrossRef] [Green Version]
- Raphaël, M.; Lehen’Kyi, V.; Vandenberghe, M.; Beck, B.; Khalimonchyk, S.; Abeele, F.V.; Farsetti, L.; Germain, E.; Bokhobza, A.; Mihalache, A.; et al. TRPV6 calcium channel translocates to the plasma membrane via Orai1-mediated mechanism and controls cancer cell survival. Proc. Natl. Acad. Sci. USA 2014, 111, E3870–E3879. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; McGoldrick, L.L.; Twomey, E.C.; Sobolevsky, A.I. Mechanism of calmodulin inactivation of the calcium-selective TRP channel TRPV6. Sci. Adv. 2018, 4, eaau6088. [Google Scholar] [CrossRef] [Green Version]
- Derler, I.; Hofbauer, M.; Kahr, H.; Fritsch, R.; Muik, M.; Kepplinger, K.; Hack, M.E.; Moritz, S.; Schindl, R.; Groschner, K.; et al. Dynamic but not constitutive association of calmodulin with rat TRPV6 channels enables fine tuning of Ca2+-dependent inactivation. J. Physiol. 2006, 577, 31–44. [Google Scholar] [CrossRef]
- Niemeyer, B.A.; Bergs, C.; Wissenbach, U.; Flockerzi, V.; Trost, C. Competitive regulation of CaT-like-mediated Ca 2+ entry by protein kinase C and calmodulin. Proc. Natl. Acad. Sci. USA 2001, 98, 3600–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voets, T.; Janssens, A.; Prenen, J.; Droogmans, G.; Nilius, B. Mg2+-dependent Gating and Strong Inward Rectification of the Cation Channel TRPV6. J. Gen. Physiol. 2003, 121, 245–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flourakis, M.; Lehen’Kyi, V.; Beck, B.; Raphaël, M.; Vandenberghe, M.; Abeele, F.V.; Roudbaraki, M.; Lepage, G.; Mauroy, B.; Romanin, C.; et al. Orai1 contributes to the establishment of an apoptosis-resistant phenotype in prostate cancer cells. Cell Death Dis. 2010, 1, e75. [Google Scholar] [CrossRef] [Green Version]
- Colley, C.S.; England, E.; Linley, J.E.; Wilkinson, T.C.I. Screening Strategies for the Discovery of Ion Channel Monoclonal Antibodies. Curr. Protoc. Pharmacol. 2018, 82, e44. [Google Scholar] [CrossRef] [PubMed]
- Koch-Nolte, F.; Eichhoff, A.; Pinto-Espinoza, C.; Schwarz, N.; Schäfer, T.; Menzel, S.; Haag, F.; Demeules, M.; Gondé, H.; Adriouch, S. Novel biologics targeting the P2X7 ion channel. Curr. Opin. Pharmacol. 2019, 47, 110–118. [Google Scholar] [CrossRef]
- Klionsky, L.; Tamir, R.; Holzinger, B.; Bi, X.; Talvenheimo, J.; Kim, H.; Martin, F.; Louis, J.-C.; Treanor, J.J.S.; Gavva, N.R. A Polyclonal Antibody to the Prepore Loop of Transient Receptor Potential Vanilloid Type 1 Blocks Channel Activation. Experiment 2006, 319, 192–198. [Google Scholar] [CrossRef] [Green Version]
- De Lera Ruiz, M.; Kraus, R.L. Voltage-Gated Sodium Channels: Structure, Function, Pharmacology, and Clinical Indications. J. Med. Chem. 2015, 58, 7093–7118. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, L.; Han, H.; Jin, K.; Lin, N.; Guo, T.; Chen, Y.; Cheng, H.; Lu, F.; Fang, W.; et al. 1B50-1, a mAb Raised against Recurrent Tumor Cells, Targets Liver Tumor-Initiating Cells by Binding to the Calcium Channel α2δ1 Subunit. Cancer Cell 2013, 23, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Bettaieb, L.; Brulé, M.; Chomy, A.; Diedro, M.; Fruit, M.; Happernegg, E.; Heni, L.; Horochowska, A.; Housseini, M.; Klouyovo, K.; et al. Ca2+ Signaling and Its Potential Targeting in Pancreatic Ductal Carcinoma. Cancers 2021, 13, 3085. [Google Scholar] [CrossRef]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion Channels in Cancer: Are Cancer Hallmarks Oncochannelopathies? Physiol. Rev. 2018, 98, 559–621. [Google Scholar] [CrossRef] [Green Version]
- Fliniaux, I.; Germain, E.; Farfariello, V.; Prevarskaya, N. TRPs and Ca2+ in cell death and survival. Cell Calcium 2018, 69, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Kondratskyi, A.; Kondratska, K.; Skryma, R.; Prevarskaya, N. Ion channels in the regulation of apoptosis. Biochim. Biophys. Acta Biomembr. 2015, 1848, 2532–2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.; Seo, H.; Kwak, M.; Jeon, J.; Jang, J.; Jeong, E.M.; Myeong, J.; Hwang, Y.J.; Ha, K.; Kang, M.J.; et al. Increased TRPC5 glutathionylation contributes to striatal neuron loss in Huntington’s disease. Brain 2015, 138, 3030–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Sukumaran, P.; Selvaraj, S.; Cilz, N.I.; Schaar, A.; Lei, S.; Singh, B.B. TRPM2 Promotes Neurotoxin MPP+/MPTP-Induced Cell Death. Mol. Neurobiol. 2016, 55, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Asuthkar, S.; Velpula, K.K.; Elustondo, P.A.; Demirkhanyan, L.; Zakharian, E. TRPM8 channel as a novel molecular target in androgen-regulated prostate cancer cells. Oncotarget 2015, 6, 17221–17236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, H.; Ugawa, S.; Ueda, T.; Morita, A.; Shimada, S. TRPM8 activation suppresses cellular viability in human melanoma. Am. J. Physiol. Physiol. 2008, 295, C296–C301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, S.M.; Gidley Baird, A.; Glazer, S.; Barden, J.A.; Glazer, A.; Teh, L.C.; King, J. A phase I clinical trial demonstrates that nfP2X7 -targeted antibodies provide a novel, safe and tolerable topical therapy for basal cell carcinoma. Br. J. Dermatol. 2017, 177, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Sainz, B., Jr.; Heeschen, C. Standing Out from the Crowd: Cancer Stem Cells in Hepatocellular Carcinoma. Cancer Cell 2013, 23, 431–433. [Google Scholar] [CrossRef] [Green Version]
- Bao, B.; Jiang, J.; Yanase, T.; Nishi, Y.; Morgan, J.R. Connexon-mediated cell adhesion drives microtissue self-assembly. FASEB J. 2010, 25, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Djillani, A.; Doignon, I.; Luyten, T.; Lamkhioued, B.; Gangloff, S.C.; Parys, J.B.; Nüße, O.; Chomienne, C.; Dellis, O. Potentiation of the store-operated calcium entry (SOCE) induces phytohemagglutinin-activated Jurkat T cell apoptosis. Cell Calcium 2015, 58, 171–185. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haustrate, A.; Shapovalov, G.; Spriet, C.; Cordier, C.; Kondratskyi, A.; Noyer, L.; Foulquier, F.; Prevarskaya, N.; Lehen’kyi, V. TRPV6 Calcium Channel Targeting by Antibodies Raised against Extracellular Epitopes Induces Prostate Cancer Cell Apoptosis. Cancers 2023, 15, 1825. https://doi.org/10.3390/cancers15061825
Haustrate A, Shapovalov G, Spriet C, Cordier C, Kondratskyi A, Noyer L, Foulquier F, Prevarskaya N, Lehen’kyi V. TRPV6 Calcium Channel Targeting by Antibodies Raised against Extracellular Epitopes Induces Prostate Cancer Cell Apoptosis. Cancers. 2023; 15(6):1825. https://doi.org/10.3390/cancers15061825
Chicago/Turabian StyleHaustrate, Aurélien, George Shapovalov, Corentin Spriet, Clément Cordier, Artem Kondratskyi, Lucile Noyer, François Foulquier, Natalia Prevarskaya, and V’yacheslav Lehen’kyi. 2023. "TRPV6 Calcium Channel Targeting by Antibodies Raised against Extracellular Epitopes Induces Prostate Cancer Cell Apoptosis" Cancers 15, no. 6: 1825. https://doi.org/10.3390/cancers15061825
APA StyleHaustrate, A., Shapovalov, G., Spriet, C., Cordier, C., Kondratskyi, A., Noyer, L., Foulquier, F., Prevarskaya, N., & Lehen’kyi, V. (2023). TRPV6 Calcium Channel Targeting by Antibodies Raised against Extracellular Epitopes Induces Prostate Cancer Cell Apoptosis. Cancers, 15(6), 1825. https://doi.org/10.3390/cancers15061825