1. Introduction
Exosomes and extracellular vesicles (EVs) are membrane-based nanovesicles, that have been intensely investigated as excellent candidates for the diagnosis and the monitoring of several pathologies (such as cancer) due to their availability and established presence in most biological fluids [
1]. These extracellular nanovesicles serve as delivery vehicles for the transport of a large range of active biomolecules between cells. They are composed of a cytosolic inner aqueous compartment surrounded by a protein-phospholipid bilayer membrane with similar biochemical and physicochemical characteristics to plasma membranes. However, in spite of the efforts invested in studying the physicochemical composition of EV membranes, the relationship between the produced EV membranes and the source parental cell is far from fully understood.
It has been claimed that the membrane of cancer-derived EVs contains a fingerprint that on the one hand is responsible for their selectivity in targeting cells [
2], and on the other hand, gives valuable clues for studying their parental cell composition. It seems reasonable to assume that cancer-cell derived EVs contain essential information regarding the cell of origin and the tumor environment in which it develops. In fact, EVs are currently being considered as potential biomarkers of some specific types of cancers [
3]. For instance, exosomal miRNA has attracted attention for potential use as a diagnostic and prognostic marker in cancer. In this sense, Li et al. showed how miR-3591-3p could be used as a biomarker for glioma [
4]. In another study, exosomal miR-423-3p was demonstrated to be a promising predictive biomarker for prostate cancer castration resistance development [
5]. Other studies have used exosomal proteins as biomarkers in diagnosis. Chu et al. combined bioinformatic predictions with proteomic studies to conclude that higher expression levels of calsyntenin-1 (CLSTN1), clusterin (CLU) and neutrophil gelatinase-associated lipocalin (NGAL) proteins are solid serologic biomarkers for patients with lung adenocarcinoma [
6]. Moreover, several studies have identified differential composition of exosomal lipids according to their malignancy [
7,
8,
9,
10,
11,
12]. For instance, Cheng et al. established that ovarian cancer-derived EVs contain higher levels of gangliosides, zymosterol, lysophosphatidylinositol, lysophosphatidylcholine, acca, lysophosphatidylserine, lysophosphatidylglycerol and cholesterol ester, and lower levels of ceramides, digalactosyldiacylglycerol, phosphatidylserine, phosphatidylinositol, phosphatidylglycerol, sphingomyelin, phosphatidylethanolamine, diacylglycerol and ceramides, compared with EVs derived from epithelial HOSEPiC cells [
13]. Other studies confirmed that the lipid profile of pancreatic ductal adenocarcinoma (PDAC) was significantly dysregulated compared with healthy controls [
14,
15,
16]. Our group has previously analyzed the lipidomic profile of healthy and tumoral metastatic and non-metastatic cell lines, revealing significant differences in the lipidomic profile of these cell lines according to their malignancy [
7]. These data evidenced the potential role of exosomal lipids as diagnostic/prognostic markers in cancer. However, more clinical data are still required to confirm the diagnostic value of these techniques. Another drawback of analytical techniques based on genomics, lipidomics and proteomics is related to their time-consuming and expensive nature [
11,
13,
17].
In view of the above, a strong interest exists in the development of alternative methodologies analyzing EVs as biomarkers of pathological processes. X-Ray Photoelectron Spectroscopy (XPS) is a high sensitivity surface analysis technique able to detect the relative abundance of each single element and also to give information regarding their chemical environment. A highly advantageous feature is the fact that the effective penetration of XPS for analysis is determined by the element being detected, the matrix or the orbital, and is limited to around 10 nanometers [
18], meaning that (since the protein-lipidic membrane of EVs possesses a thickness about 5 nm) this technique will mainly “see” the membrane itself, and therefore the content of the EVs lumen (a lower concentration aqueous solution of biomolecules) will not interfere in the results. All of these reasons make XPS ideally suited to study EV membrane composition.
We have explored the use of XPS as a fast research tool to characterize the composition of EVs, with the objective of assessing its possible application in cancer screening. We propose the use of XPS to analyze the membranes of EVs isolated from bodily fluids of target patients. Notably we have focused our analysis on the N chemical environment of EV membranes, as this can be related to previous complex chemical analyses of lipid contents in exosomes [
7]. Specifically, we have analyzed EV membranes from cell cultures of different cancer cells (B16-F10 cells and B16-F1 cells) and compared them to EVs from healthy cells (NIH-3T3 fibroblasts and human placental mesenchymal stem cells (hpMSCs)) as controls. We have observed how tumoral and healthy cells have different nitrogen chemical environments that can be related to their malignancy. In order to assess the potential of this technique in cancer screening, a collection of human serum samples from patients and healthy donors was analyzed. The differential analysis of XPS patterns of EV membranes confirmed the significance of the nitrogen environment as a marker of ovarian and pancreatic tumors. Additionally, a clear correlation between CEA (a broadly used clinical cancer blood marker used currently in many diagnostic test) and the content of secondary amines in EV membranes of pancreatic cancer patients was detected. In summary, although XPS analysis cannot relate nitrogen environment with tumor progression, it gives a set of data that can open a new line of research regarding the different atomic environments of cancer EVs as a potential diagnosis/prognosis tool. These results open up interesting possibilities for the use of XPS as a new, fast and non-invasive cancer research technique useful for liquid biopsy and assessment of the nitrogen environment, describing a potential new biomarker.
2. Materials and Methods
2.1. Cell Culture
Human placental mesenchymal stem cells (hpMSCs) were obtained from Cellular Engineering Technologies (CET) (Caralville, IA, USA), while B16-F1 (low metastatic variant) and B16-F10 (high metastatic variant) murine skin melanoma cells, were provided by cell services from Cancer Research-UK. NIH-3T3 murine healthy fibroblasts were obtained from Dr. Antonio de la Vieja’s group (Instituto de Salud Carlos lll). hpMSCs were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Biowest, France) supplemented with 5 μg mL−1 of FGF-2 growth factor (PeproTech, Cranbury, NJ, USA), with 10% of fetal bovine serum (FBS, GIBCO, Waltham, MA, USA), 1% penicillin/streptomycin and 1% amphotericin (Biowest, France) and maintained at 37 °C in a 5% CO2-humidified atmosphere under hypoxic conditions (3% O2). For culturing B16-F1, B16-F10, and NIH-3T3 cells, DMEM with 10% of FBS (GIBCO), supplemented with 1% penicillin/streptomycin and 1% amphotericin (Biowest, France) was used. Finally, monocytes were cultured in RMPI Medium 1640 (Biowest, France) supplemented with 10% FBS (GIBCO), 1% penicillin/streptomycin and 1% amphotericin (Biowest, France). B16-F1, B16-F10 and NIH-3T3 cells were maintained under normoxic conditions.
To obtain culture media free of EVs, they were depleted from serum by ultracentrifugation at 100,000× g for 8 h at 4 °C.
2.2. Human Samples
Human serum samples and data from patients included in this study were provided by the Biobank of the Aragon Health System, integrated in the Spanish National Biobanks Network (PT20/00112), and they were processed following standard operating procedures with the appropriate approval of the Ethics and Scientific Committees. The present study (PI18/198) was approved by the human research review committee of the Research Ethics Committee of the Community of Aragon (Comité de Ética de la Investigación de la Comunidad de Aragón, CEICA), and was conducted in compliance with the ethical standards formulated in the Helsinki Declaration of 1996 (revised in 2000), upon obtaining the informed consent from all patients.
Human serum samples were obtained from healthy donors (n = 10) and from stage III-IV pancreatic cancer patients (n = 10) using the first sample at diagnosis and two sub-sequential samples taken during disease progression. Ovarian cancer patients’ blood samples (n = 16) were obtained at diagnosis, including from patients in different stages (I, II, III, IV). Ovarian EV samples were divided for analysis according to the different stages, considering the possible variations of EVs depending on the disease stage. Samples were collected in vacutainer sodium citrate tubes in Hospital Universitario Miguel Servet, Zaragoza (Spain). Samples were stored at −80 °C until use. Then, samples were defrosted and 500 µL of serum was employed for isolation of the EVs and further XPS characterization. Blood markers, CEA and CA 19:9, were quantified by the National Health System, following standard procedures. The pancreatic samples used here for analysis of atomic composition and nitrogen environment are samples extracted at diagnosis, meaning, samples extracted before starting any treatment. Additionally, we have studied the evolution of secondary amines (first sample at diagnosis and two sub-sequential samples during disease progression) in relation to blood markers, CEA and CA 19:9. In this case, our purpose was to analyze if the variation of secondary amines occurs with the same trend as the evolution of blood markers, which are also modified depending on treatment. In this case, every patient has been studied independently, because we cannot compare secondary amine evolution during time among patients due to the differences in treatments and disease progression of each patient.
2.3. Isolation of EVs
EVs from cell culture samples were isolated following a protocol previously published and based on successive ultracentrifugation cycles from cell culture supernatants of the cells [
19]. Briefly, cells were cultured until confluence and their supernatants were centrifuged for 20 min at 2000×
g and at 4 °C (to remove remaining debris). For the elimination of the microvesicles, another centrifugation step was carried out for 1 h at 10,000×
g and at 4 °C. To obtain the exosome fraction, the samples were ultracentrifuged for 2 h at 100,000×
g and at 4 °C. A further washing step of exosomes with PBS was carried out to discard the co-isolated and adsorbed proteins on exosomes (2 h, 100,000, 4 °C). The exosomes pellet was finally resuspended in PBS for further analysis [
1]. Some of these exosomes were previously described and characterized [
2,
20].
Ultracentrifugation is currently the gold standard for exosome isolation; thus far, some in vitro separation methods, such as ultracentrifugation, polymer-based exosome separation kits and immune affinity-based isolation using antibodies against exosome surface proteins, have been used for tumor exosome isolation [
21]. We have compared the isolation efficiency between ultracentrifugation and affinity-based resin for serum samples. Due to the low volume of serum, the isolation efficiency by ultracentrifugation was too low to allow us to perform XPS analysis; therefore, we selected instead an affinity-based resin method to isolate exosomes from serum, but as the fraction collected might not be purely exosomes, we named all of them EVs. In the case of exosomes isolated from cell culture supernatants there is not a volume limitation and ultracentrifugation was the selected isolation procedure. In the case of human samples, serum samples (
n = 56) were divided into control samples (
n = 10 samples/patients), ovarian tumor samples (
n = 16 samples/patients) and pancreatic tumor samples (
n = 10 patients, 30 samples, 3 samples per patient, extracted at different time-points of the disease). EVs from serum samples (
n = 56) were extracted using Plasma/Serum Exosome Purification Mini Kit, 50 Preps (57400, Norgen Biotek, Thorold, Canada). Following manufacturer instructions, the Plasma/Serum Exosome Purification Kit provides a reliable and convenient method to purify and enrich pure intact EVs factions from different plasma/serum samples, free from any cell-free circulating protein-bound RNA. Purification was performed according to the producer protocol. In brief, to 500 µL serum aliquots from each donor were added, in the following order, 3.5 mL nuclease-free water, 100 µL f ExoC buffer and 200 µL of Slurry E (resin). The mixture was mixed by vortexing and let stand at room temperature for 5 min, to allow the EVs to adhere to the resin. After incubation, the mixtures were centrifugated to recover the resin, the supernatant was discarded and the resin resuspended in 200 µL ExoR buffer; incubation was performed at room temperature for 5 min to release EVs from resin. After incubation, the mixture was centrifugated for 2 min at 500 rpm and the supernatant (containing free EVs) was recovered and filtered by centrifugation through a Mini Filter Spin column (1 min at 6000 rpm) to discard possible remaining resin (retained on the filter). After centrifugation, the EVs were ready for downstream applications. As negative control for following analysis, we used the same protocol starting with 4 mL nuclease-free water and the supernatant from the last step was analysed in the same way as the EVs from serum.
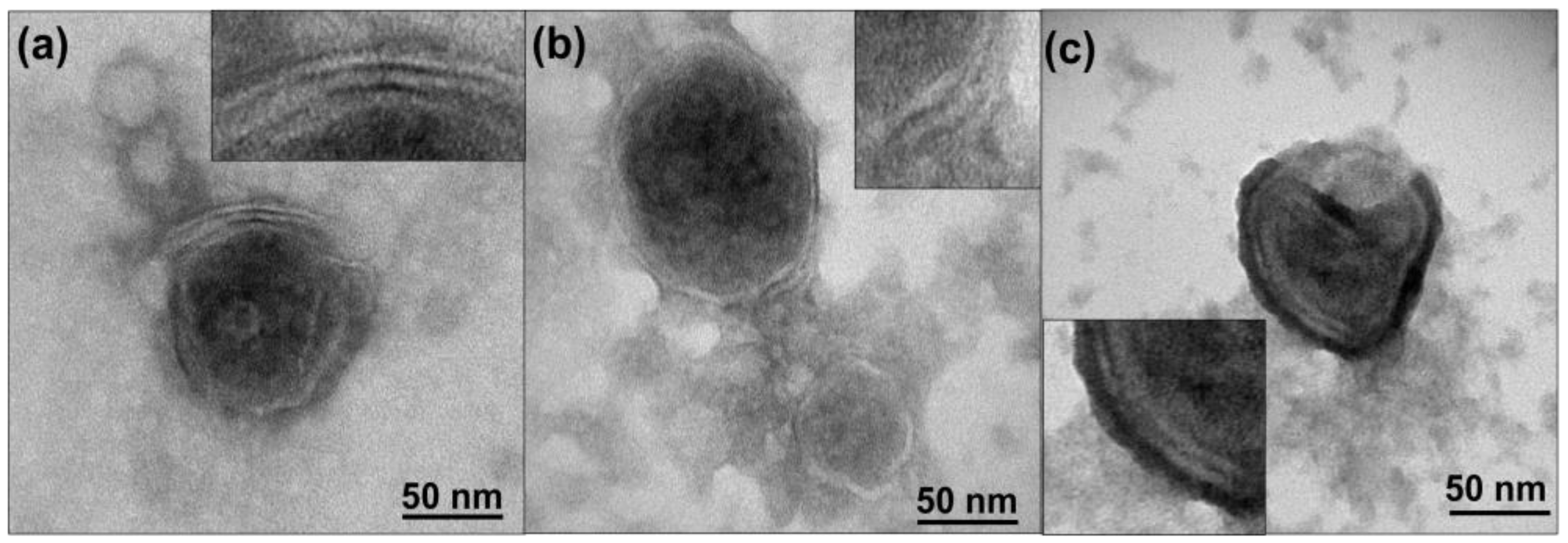
2.4. Transmission Electronic Microscopy (TEM) Sample Preparation and Analysis
The EVs’ morphology and the thickness of the EV membrane were characterized by transmission electron microscopy (T20-FEI Tecnai thermoionic transmission electron microscope) operated at 200 kV with a LaB6 electron source fitted with a “SuperTwin
®” objective lens allowing a point-to-point resolution of 2.4 A [
22]. EVs were stained with 3% phosphotungstic acid (Sigma Aldrich, St. Louis, MO, USA) in order to reveal their membrane structural details, as was previously described by our group.
2.5. XPS Sample Preparation and Analysis
EV pellets isolated from cell cultures were finally resuspended in PBS and deposited drop by drop on a circular cover glass slip on a hotplate set at 30 °C.
EVs from serum human samples and healthy controls were recovered in 200 µL ExoR buffer final volume. The resuspension was dried drop by drop (5 µL) on a circular cover glass slip on a hotplate set at 30 °C.
The XPS analysis of all samples was performed 24 h after sample desiccation with an Axis Supra (Kratos Tech., San Diego, CA, USA) apparatus to determine the atomic percentage and the relative abundance of the different nitrogen species analyzed (R3-N, R2-NH/N-C, -NH2 and pyridine type bounding). The spectra were excited by the monochromatized Al Kα source (1486.6 eV) run at 10 kV and 8 mA. For the individual peak regions, a pass energy of 20 eV was employed. Survey spectra were measured at 120 eV pass energy. The CasaXPS software was used to analyze the peaks. After the subtraction of Shirley background, a weighted sum of Lorentzian and Gaussian components curve was employed.
2.6. Statistical Analysis
The results are expressed as mean ± SD (standard deviation). Statistical analysis of the data and the significant differences among the means were evaluated by two-way analysis of variance (ANOVA) and unpaired Student’s t-test for binary comparisons. Analysis was performed using GraphPad Software. Statistically significant differences were indicated as follows: * p < 0.05; ** p < 0.01; *** p < 0.0001 and **** p < 0.00001.
4. Discussion
The cell membrane is composed of lipids and proteins. The lipid fraction of the cell membrane consists mainly of phospholipids, glycolipids, sphingolipids and cholesterol [
25]. The most abundant membrane lipids are the phospholipids. These have a polar head group and two hydrophobic hydrocarbon tails. The presence of carbon and oxygen is mainly related to the aliphatic chains in the tails. On the other hand, the main nitrogen contribution is associated with the polar groups of the phospholipidic head [
26]. Cancer cells are notorious for the numerous adaptations that affect signaling cascades involved in therapy and immune system response; obviously, these involve changes in the type and/or amount of lipids that can be associated with the different cancer states. These changes are so prominent that lipid profile modifications could even be considered as a new cancer biomarker tool [
25,
27].
EVs are limited by a lipidic membrane, which encapsulates the cargo molecules in an inner aqueous core [
28]. Lipid, and specifically phospholipids, are essential components of EV membranes and it is also known that specific lipids are enriched in EVs compared to their parent cells, playing indispensable roles on the structural and regulatory functions of EV biogenesis, release, targeting and cellular uptake [
29,
30,
31]. Studies have showed that cholesterol, sphingomyelin, glycosphingolipids, phosphatidylserine, phosphatidylcholine are the most abundant lipids in exosomes [
32]. It seems obvious that lipidomic studies are key to unravelling the biological relevance of EVs and will provide important clues on their functions [
13].
We have recently proposed a robust analytical procedure to isolate EVs and obtain a wide-ranging and reliable analysis of the phospholipid (PL) content of their membranes [
8]. Using this methodology, we have been able to analyze the lipidomic profile of healthy and tumoral metastatic and non-metastatic cell lines and patients by a novel methodology, revealing significant differences in the lipidomic profile of these cell lines [
7]. Other recent studies using mass spectrometry platforms to analyze the lipidomic profile of EVs from urine have revealed significant differences between healthy control subjects and renal cell carcinoma patients [
8], and for prostate cancer patients [
9,
10]. Similarly, differences in the lipidomic profile were detected among EVs isolated from plasma of healthy control, non-metastatic and metastatic colorectal cancer patients [
11], hepatocellular carcinoma cancer patients [
33] and lung cancer patients [
12,
34]. These results support the possible application of the EV lipidomic profile analysis as a diagnostic tool [
32]. Additionally, the presence of EVs in urine or plasma provides significant advantages as a non-invasive tool for diagnosis [
35].
These studies rely on the use of mass spectrometry protocols to analyze the lipidomic profile of EVs. While a powerful tool, these methods are rightly considered to constitute a complex technique, which requires the analysis of a complete lipidomic profile, distinguishing among the different lipids present in the EV membranes. The same could be said of our previous work involving EV isolation and analysis of the lipid EV extract by High-Performance Thin-Layer Chromatography-densitometry directly coupled to mass spectrometry, again a relatively complex procedure [
7].
Instead, here we propose the characterization of the nitrogen environment using a single technique (XPS) and a relatively standard measurement. We have focused our analysis on nitrogen as this single element can be present in a variety of environments that can be easily analyzed by XPS, and their relative abundances can be used to characterize the evolution of the EV membrane composition. As already mentioned, XPS is ideal not only as a fast and reliable technique to identify different chemical environments of nitrogen atoms based on their binding energies, but also because being a surface technique that only responds to signals from the uppermost atomic layers, it has obvious advantages when one wants to analyze the membrane composition with minimal interference from the inner content of the EVs.
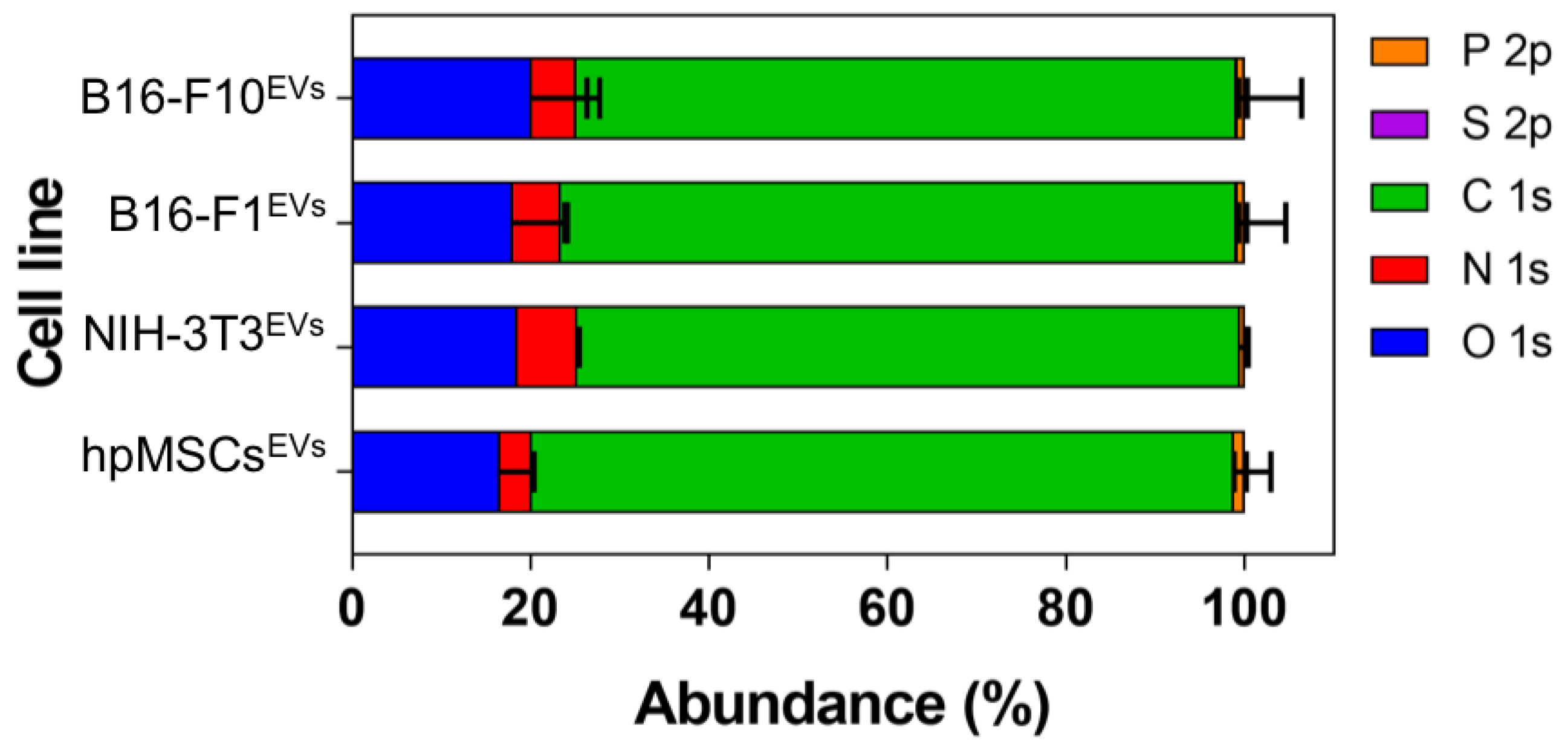
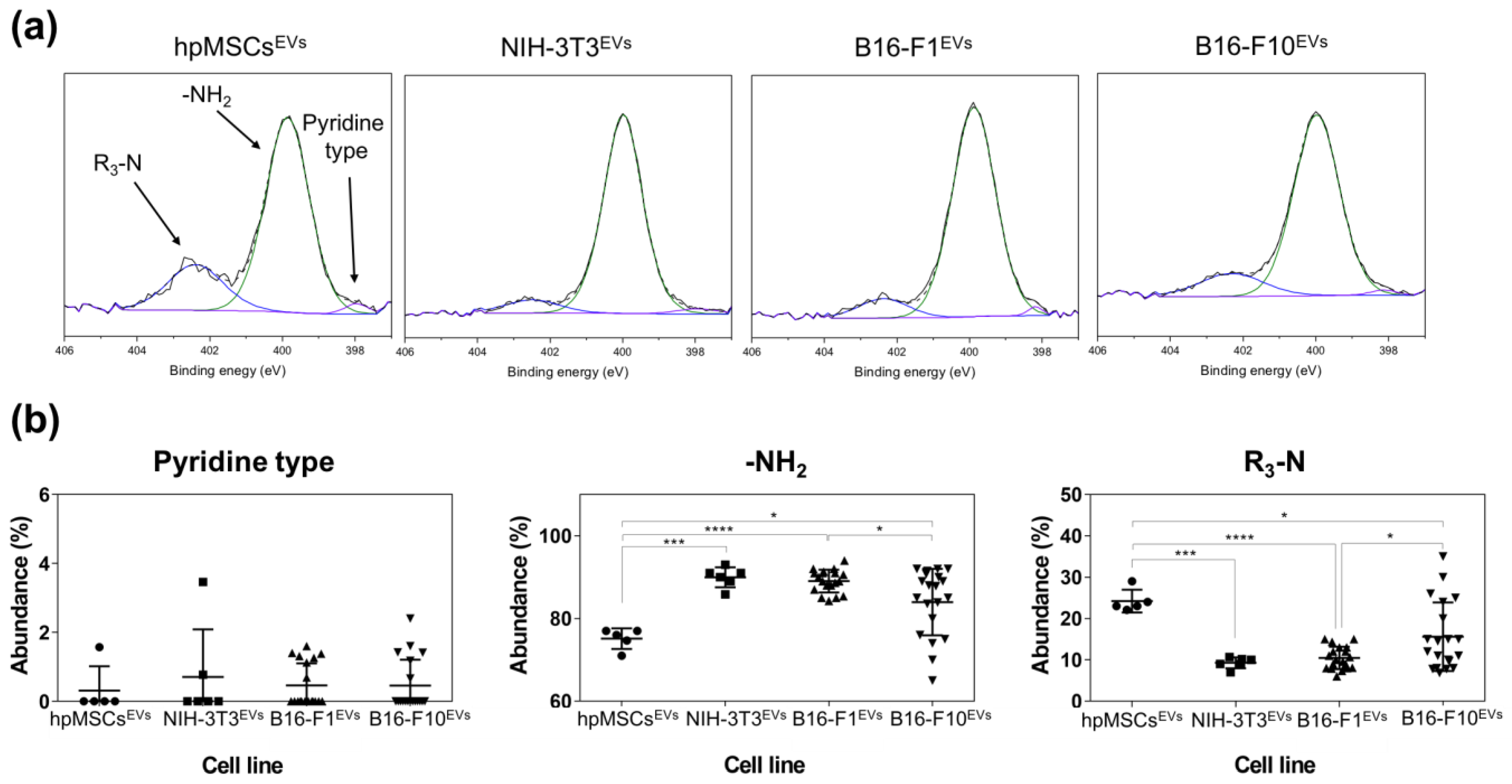
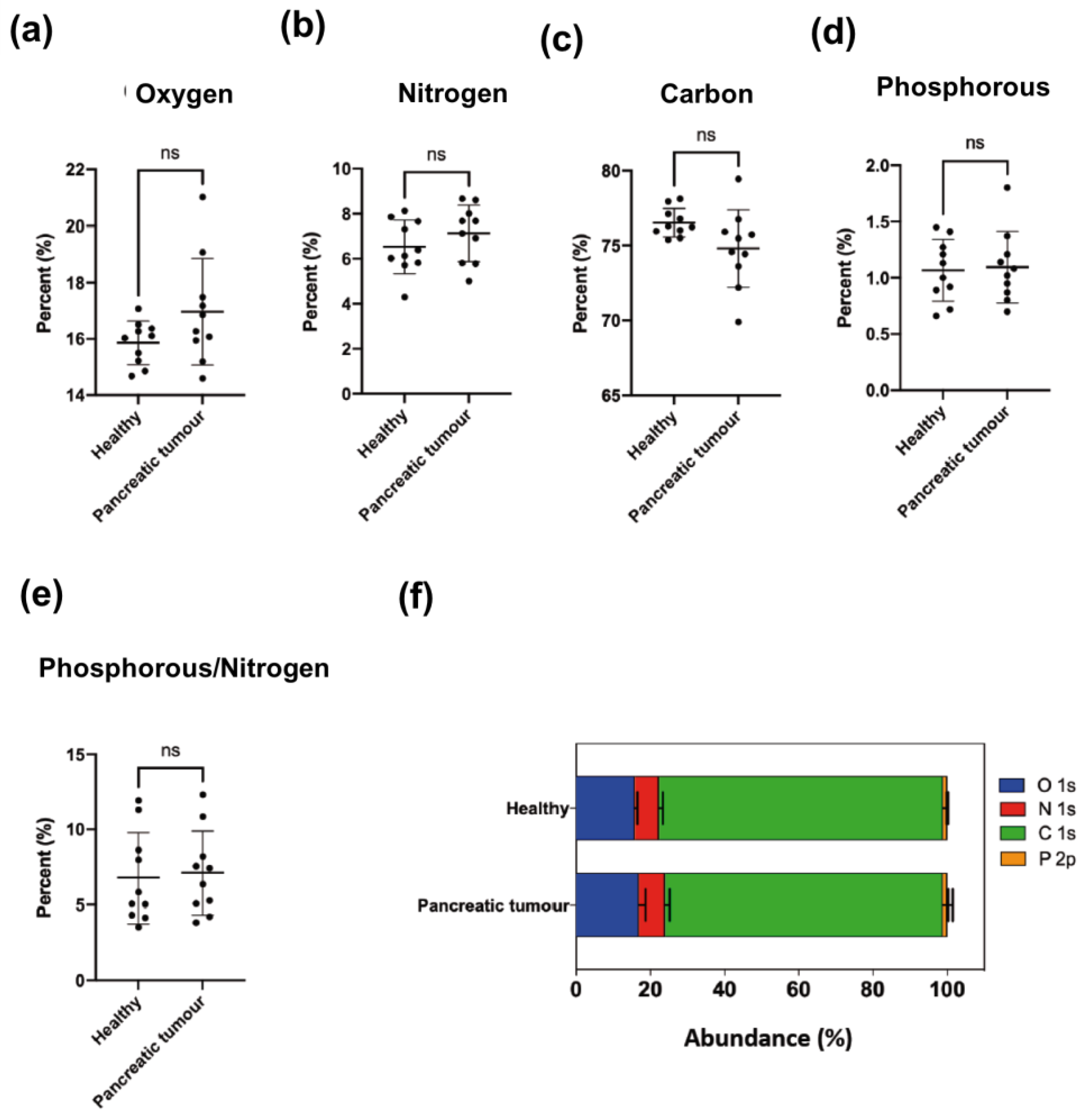
The atomic composition of EV membranes isolated from cell cultures by ultracentrifugation (
Table S1 and
Figure 2) did not show significant differences among them. This is not surprising since the elemental composition analysis represents a high-level description of the sample and as such it is unlikely to detect differences in the chemical structure of the membranes (changing functional groups often gives rise to only small variations in elemental analysis). However, when the nitrogen chemical environments were considered, differences were detected between healthy and cancer cells (
Table S2 and
Figure 2 and
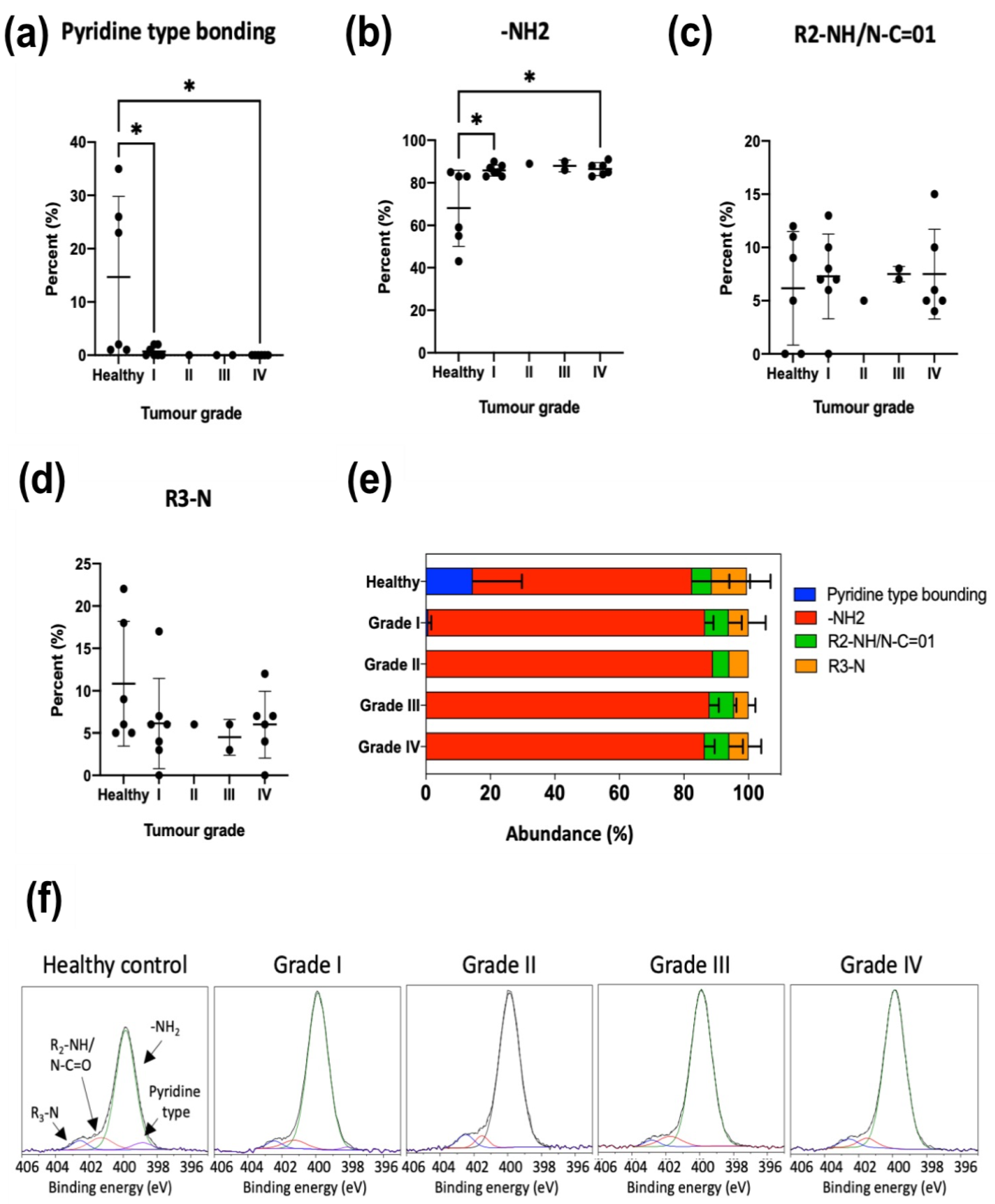
Figure 3). Particularly, tertiary amines present in the EV membrane appeared to increase as the malignity of the parental source cells was higher.
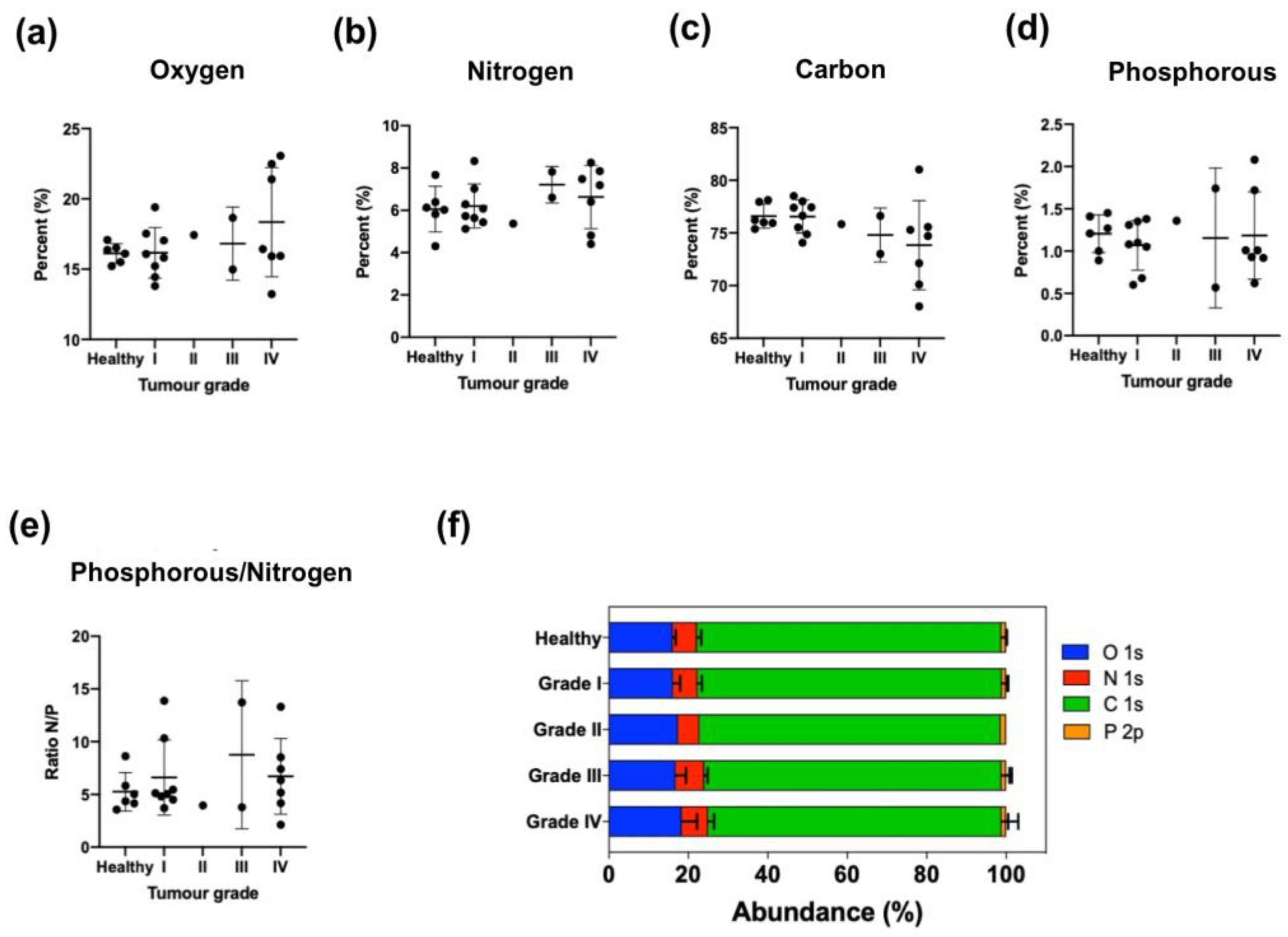
To further study this trend, EVs were isolated from human serum from healthy donors and from ovarian and pancreatic cancer patients (
Tables S3 and S5,
Figure 4 and
Figure 6). Again, the elemental analysis did not show any significant differences, but analyzing the nitrogen environment allowed us to detect differences between healthy donor and ovarian and pancreas cancer patients (
Tables S4 and S6,
Figure 5 and
Figure 7), confirming a modification of the lipidomic profile. However, in this case, the changes observed were different from those of the in vitro studies, the main difference being the disappearance of the pyridine-type nitrogen species.
Different studies have shown significant differences in the lipid profile of EVs from healthy and cancer-suffering persons [
7,
8,
9,
10,
11,
12]. However, a clear trend has not yet been found. Our knowledge about the lipid composition of cancer EVs is still scarce. Cheng et al. identified 30 lipid classes, including 1227 lipid species, in exosomes derived from ovarian cancer cells (SKOV-3) compared to those from ovarian surface epithelial cells (HOSEPiC). In particular, SKOV-3 derived EVs contained higher levels of many of the lipids studied than exosomes derived from HOSEPiC cells [
13]. Our XPS results are effective in showing in a fast and simple measurement that EVs isolated from ovarian cancer patients show a strong decrease of pyridine-type bonding and also a noticeable increase of primary amines (-NH2) (
Table S4,
Figure 5), which implies that a significant alteration of the lipidomic profile has taken place, although identification of the specific lipid species altered would require a complex in-depth analysis.
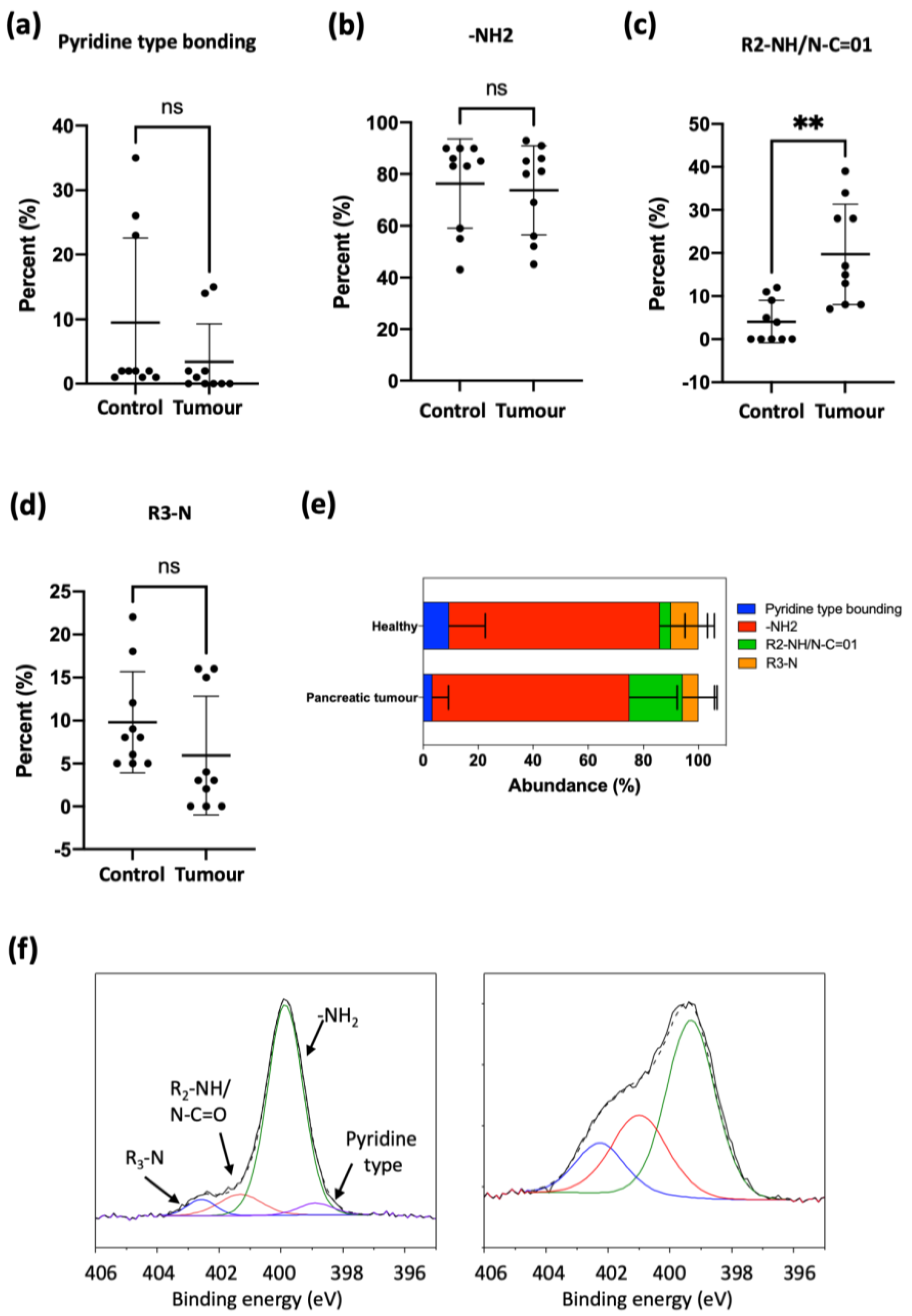
Regarding PDAC, few studies relating diagnosis/prognosis and exosomal or EV lipid composition have been published. It was found that 1021 lipid species from different PDAC cell-derived exosomes (Panc-1, Capan-1, SW-1990, Mia PaCa-2, PPCL-68 and PPCL-46) and normal cell lines (hTERT-HPNE, HPDE-H6c7) were dysregulated between cancer-derived sEVs and normal sEVs, especially lipid species containing palmitic acid (16:0) and sphingomyelin [
14]. In a genetic pancreatic mouse model, phosphatidylserine positive exosomes in blood were analyzed by ELISA, revealing a significant increase of these exosomes in PDAC-bearing mouse, suggesting the potential of phosphatidylserine positive exosomes for PDAC detection [
15]. In another study by Tao et al., mass spectrometry was performed to analyze the lipid expression profile in exosomes derived from peripheral blood of PDAC patients and healthy patients. The authors found that about 270 lipids were significantly dysregulated between the exosomes of PDAC patients and healthy controls [
16]. Again, these findings indicate the potential of lipids in exosomes from PDAC patients as biomarkers. Our own results for PDAC patient EVs show a significant increase of secondary amines (R
2-NH/N-C=01) and also a trend towards the decrease of pyridine-type bonding, although without statistical significance (
Table S6,
Figure 7). As with ovarian cancer EVs, we do not identify specific lipids responsible for these changes, but XPS results provide a clear discriminating pattern, with a considerable simplification of the analysis for a potential liquid biopsy application.
The fact that these general alterations of the nitrogen environment can be ascertained from EVs sampled from the blood of healthy and cancer patients is in itself remarkable, because cancer cell-originated EVs are necessarily diluted with EVs from every other kind of cell shedding EVs to the bloodstream and also because the EV isolation method based on columns is far from perfect and the EVs collected are likely to be contaminated with foreign nitrogen-containing species, such as plasma proteins.
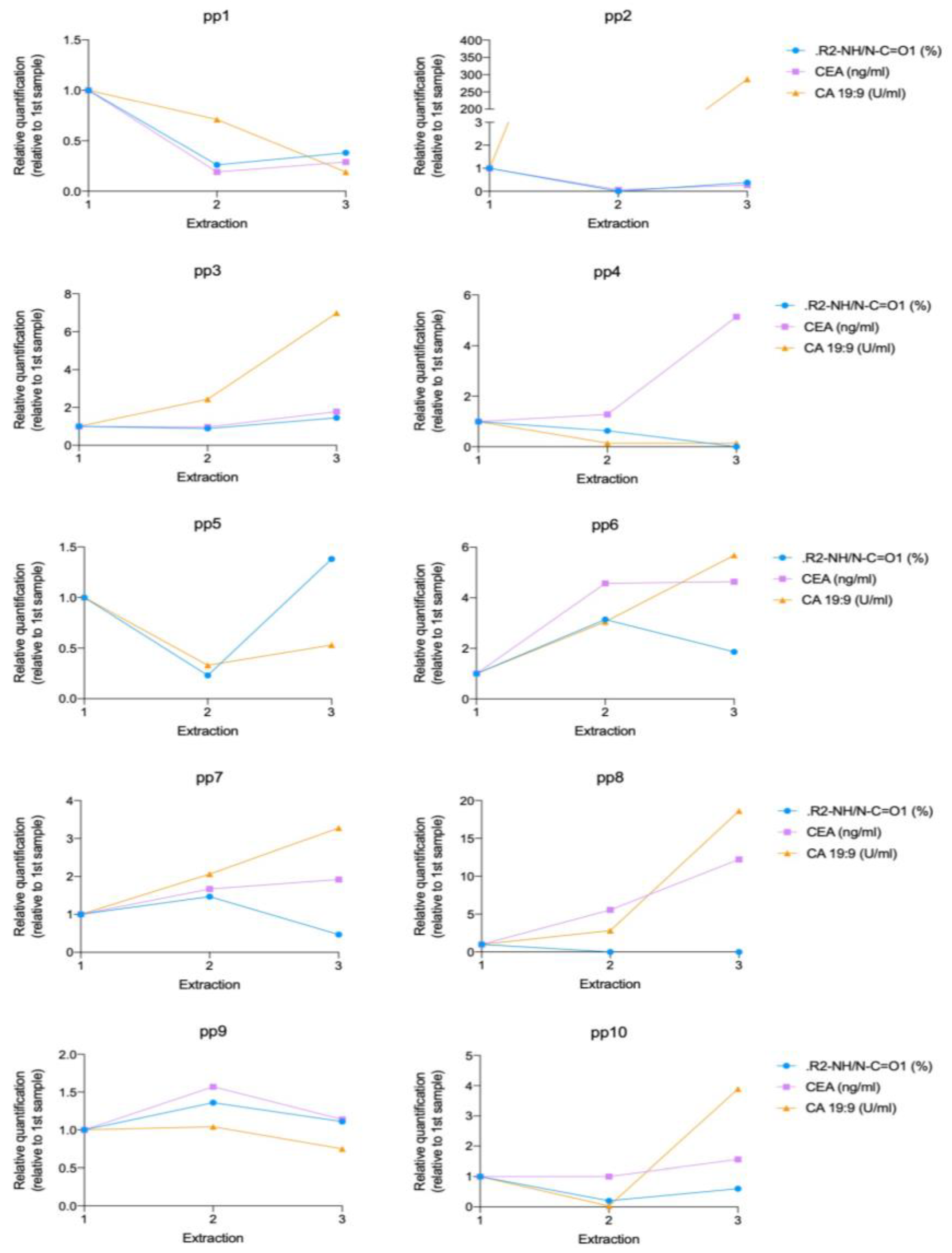
Finally, the analysis of the nitrogen environment was also useful to analyze pancreatic cancer evolution. This is an area of great interest and in fact there is not a universally accepted marker to characterize prognosis and response to treatment. We have compared the XPS results regarding the content of secondary amines with two blood markers, CA19:9 and CEA. Our results show that 67% of analyzed patients showed the same evolution of values for secondary amines and CEA and 40% showed the same evolution of values for secondary amines and CA 19:9 during the disease progression. The agreement found with CEA is better than the agreement between those established markers, since only 55% of patients showed a tendency for accordance between CEA and CA 19:9. We can therefore conclude from these preliminary results that the analysis of secondary amines’ evolution during pancreatic cancer disease has potential as a new PDAC biomarker and should be further investigated with a larger number of individuals.
5. Conclusions
In summary, analyzing by XPS the nitrogen environment of EVs isolated from the blood of two types of cancer patients reveals variations of pyridine-type bonding and primary amines in ovarian cancer, and secondary amines in pancreatic cancer, which can be related with significant changes in the lipidomic profile of their membranes, and provides analytical parameters that allow discrimination between cancer patients and healthy individuals. Furthermore, the analysis of secondary amines has shown potential as a marker of cancer evolution in pancreatic cancer. This type of measurement is attractive because it is relatively simple and could be used as a new, fast and non-invasive diagnostic tool. However, the differences found, while statistically significant in at least one nitrogen environmental parameter, do vary among the different types of cancer. It is known that exosome composition is dependent of cell origin [
36,
37]; for this reason, we could also expect variations in the exosome membrane. In consequence, the analysis of the nitrogen environment should be specific for every tumor type and, whenever possible, referenced to healthy donors of the same gender and age group. The data provided in this work support the consideration of XPS analysis as a new research tool for research-based EV investigators. Herein, we have identified the atomic composition of nitrogen species in EV membranes, opening a new line of research regarding the different atomic environments of cancer EVs as potential diagnosis/prognosis tools.
Another important point to discuss is the possibility of contaminants coming from other cell types, including EVs from other strains. The tumor EVs analyzed here are collected from the blood together with EVs from immune and other cells. Indeed, we isolate the total pool of EVs from the sample, including those from cancer cells and they will be mixed with the rest. This is a comparative study where all the samples were processed equally and the presence of EVs from non-cancer cells will happen in all the samples, adding to the “background noise” and reducing the statistical significance of the results. Fortunately, significant differences between healthy and cancer patient samples were shown, meaning that in tumoral samples the pool would have been highly enriched with tumoral EVs, leading to sufficient differences to achieve statistical significance between samples.
Further work is needed to validate the results with a wider set of clinical samples and also to identify, for each type of cancer, the relevant analytical parameters that could be used in an EV-based liquid biopsy scenario.



 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







