




Ianus Bifrons: The Two Faces of Metformin

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Historic Perspective
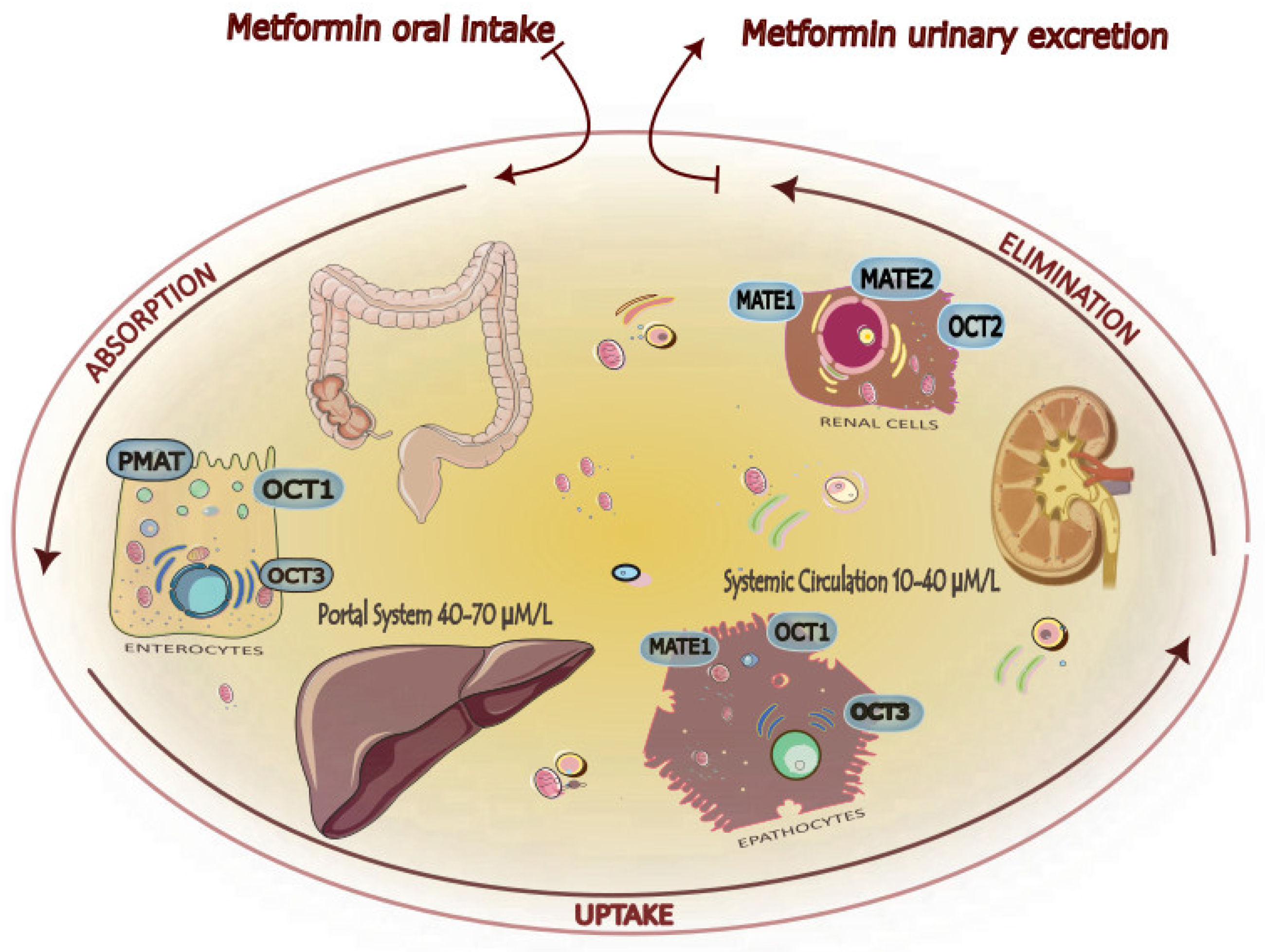
1.2. Chemical and Pharmacological Features
2. Metformin Target Organs
2.1. Liver
2.2. Gastrointestinal System
2.3. Brown Adipose Tissue
3. Molecular Mechanisms Involved in Metformin’s Action
3.1. Metformin-Induced Reduction in Blood Levels of Glucose (“Metabolic Face”)
3.1.1. Complex I Inhibition-Dependent Mechanism
3.1.2. Mitochondrial Glycerol-3-Phosphate Dehydrogenase (mGPDH)-Dependent and Complex IV Inhibition-Dependent Mechanism
3.1.3. AMPK Activation-Dependent Mechanisms in Lysosomes
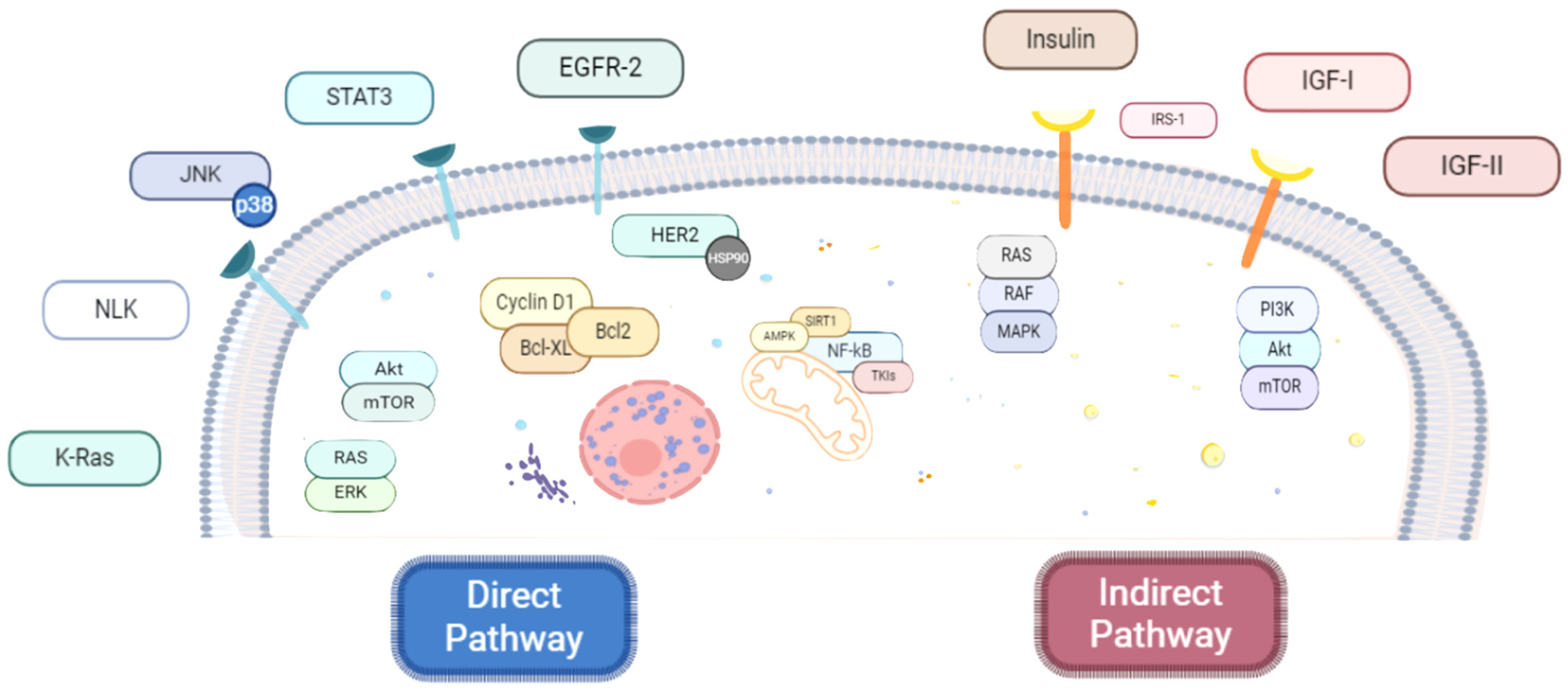
3.2. Anticancer Molecular Mechanisms of Metformin (“Anti-Proliferative Face”)
3.2.1. Direct Anticancer Mechanisms of Metformin
The mTOR Pathway
The PI3K/AKT/mTOR Pathway
The K-Ras Pathway
The NKL Pathway
The JNK Pathway
The STAT3 Pathway
The HER2 Pathway
The NF-κB Pathway
3.2.2. Indirect Anticancer Mechanisms of Metformin
4. Clinical Trials on Metformin Use
4.1. Clinical Studies on Hyperglicemic Conditions (“Metabolic Face”)
4.1.1. Type 2 Diabetes Mellitus
4.1.2. Pre-Diabetes
4.1.3. Type 1 Diabetes
4.2. Clinical Studies on Tumoral Conditions (“Anti-Proliferative Face”)
Colorectal Cancer (CRC)
5. Breast Cancer (BC)
6. Prostate Cancer
7. Endometrial Cancer (EC)
8. Other Malignancies
9. Conclusions and Future Research Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naso, P.O. Fasti I. In Ovidio—Opere; UTET: Torino, Italy, 1999; Volume IV, p. 104. [Google Scholar]
- Hill, J. The Vegetable System. Or the Internal Structure and the Life of Plants; Their Parts, and Nourishment, Explained; Their Classes, Orders, Genera, and Species, Ascertained, and Described; in a Methods Altogether New: Comprehending an Artificial Index, and a Natural System. With Figures of All the Plants; Designed and Engraved by the Author. The Whole from Nature Only; Containing Plants and Four-Petal’d Irregular Flowers; Pater-Noster-Row: London, UK, 1772; Volume 21, p. 54. [Google Scholar]
- Hadden, D.R. Goat’s rue—French lilac—Italian fitch—Spanish sainfoin: Gallega officinalis and metformin: The Edinburgh connection. J. R. Coll. Physicians Edinb. 2005, 35, 258–260. [Google Scholar]
- Pasik, C. Diabetes and the biguanides: The mystery of each. In Glucophage: Serving Diabetology for 40 Years; Pasik, C., Ed.; Groupe Lipha: Lyon, France, 1997; p. 79. [Google Scholar]
- Rathke, B. Uber Biguanid. Ber. Dtsch. Chem. Ges. 1879, 12, 776–784. (In German) [Google Scholar] [CrossRef]
- Werner, E.A.; Bell, J. The preparation of methylguanidine, and of ββ-dimethylguanidine by the interaction of dicyandiamide, and methylammonium and dimethylammonium chlorides respectively. J. Chem. Soc. Trans. 1922, 121, 1790–1794. [Google Scholar] [CrossRef]
- Slotta, K.H.; Tschesche, R. Uber Biguanide, II.: Die blutzuckersenkende Wirkung der Biguanides. Ber. Dtsch. Chem. Ges. 1929, 62, 1398–1405. (In German) [Google Scholar] [CrossRef]
- Curd, F.H.S.; Davey, D.G.; Rose, F.L. Studies on synthetic antimalarial drugs. Some biguanide derivatives as new types of antimalarial substances with both therapeutic and causal prophylactic activity. Ann. Trop. Med. Parasitol. 1945, 39, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.K.; Anderson, R.C. The toxicity and general pharmacology of N1-p-chlorophenyl-N5-isopropyl biguanide. J. Pharmacol. Exp. Ther. 1947, 91, 157–160. [Google Scholar]
- Pasik, C. Jean Sterne: A passion for research. In Glucophage: Serving Diabetology for 40 Years; Pasik, C., Ed.; Groupe Lipha: Lyon, France, 1997; pp. 29–31. [Google Scholar]
- Sterne, J. Du nouveau dans les antidiabétiques. La NN dimethylamine guanyl guanidine (N.N.D.G.). Maroc. Med. 1957, 36, 1295–1296. (In French) [Google Scholar]
- Sterne, J. Blood sugar-lowering effect of 1,1-dimethylbiguanide. Therapie 1958, 13, 650–659. (In French) [Google Scholar] [PubMed]
- Sterne, J. Treatment of diabetes mellitus with N,N-dimethylguanylguanidine (LA. 6023, glucophage). Therapie 1959, 14, 625–630. (In French) [Google Scholar]
- Sterne, J. Report on 5-years’ experience with dimethylbiguanide (metformin, glucophage) in diabetic therapy. Wien. Med. Wochenschr. 1963, 113, 599–602. (In German) [Google Scholar]
- Sterne, J.; Hirsch, C. Experimental basis for combined treatment of diabetes with the biguanide-sulfonamide association. Diabete 1964, 12, 171–175. (In French) [Google Scholar] [PubMed]
- Sterne, J. Mechanism of action of antidiabetic biguanides. Presse Med. 1964, 72, 17–19. (In French) [Google Scholar]
- Sterne, J. Pharmacology and mode of action of the hypoglycemic guanidine derivatives. In Oral Hypoglycemic Agents; Campbell, G.D., Ed.; Academic Press: London, UK, 1969; pp. 193–245. [Google Scholar]
- Sterne, J.; Duval, D.; Junien, J.L. Aspects of pharmacology and mechanisms of action. In Metformin: Current Aspects and Future Developments; Cudwoth, A.G., Ed.; Research and Clinical Forums: Tunbridge Wells, UK, 1979; Volume 1, pp. 13–20. [Google Scholar]
- Schäfer, G. Biguanides: A review of history, pharmacodynamics and therapy. Diabete Metab. 1983, 9, 148–163. [Google Scholar] [PubMed]
- Hermann, L.S. Metformin: A review of its pharmacological properties and therapeutic use. Diabete Metab. 1979, 5, 233–245. [Google Scholar] [PubMed]
- Apampa, B. Pharmacology and safe prescribing of metformin. Nurse Prescr. 2012, 10, 597–602. [Google Scholar] [CrossRef]
- Glossmann, H.H.; Lutz, O.M. Pharmacology of metformin—An update. Eur. J. Pharmacol. 2019, 865, 172782. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.-S.; Jusko, W.J. Meta-assessment of metformin absorption and disposition pharmacokinetics in nine species. Pharmaceuticals 2021, 14, 545. [Google Scholar] [CrossRef]
- Scheen, A.J. Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 1996, 30, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Graham, G.G.; Punt, J.; Arora, M.; Day, R.O.; Doogue, M.P.; Duong, J.K.; Furlong, T.J.; Greenfield, J.R.; Greenup, L.C.; Kirkpatrick, C.M.; et al. Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 2011, 50, 81–98. [Google Scholar] [CrossRef]
- Song, R. Mechanism of metformin: A tale of two sites. Diabetes Care 2016, 39, 187–189. [Google Scholar] [CrossRef]
- Chan, P.; Shao, L.; Tomlinson, B.; Zhang, Y.; Liu, Z.M. Metformin transporter pharmacogenomics: Insights into drug disposition—Where are we now? Expert. Opin. Drug Metab. Toxicol. 2018, 14, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Sundelin, E.; Jensen, J.B.; Jakobsen, S.; Gormsen, L.C.; Jessen, N. Metformin biodistribution: A key to mechanisms of action? J. Clin. Endocrinol. Metab. 2020, 105, dgaa332. [Google Scholar] [CrossRef]
- Owen, M.R.; Doran, E.; Halestrap, A.P. Evidence that metformin exerts its anti-diabetic effects through inhibition of complex 1 of the mitochondrial respiratory chain. Biochem. J. 2000, 348, 607–614. [Google Scholar] [CrossRef]
- El-Mir, M.Y.; Nogueira, V.; Fontaine, E.; Avéret, N.; Rigoulet, M.; Leverve, X. Dimethylbiguanide inhibits cell respiration via an indirect effect targeted on the respiratory chain complex I. J. Biol. Chem. 2000, 275, 223–228. [Google Scholar] [CrossRef]
- He, L.; Wondisford, F.E. Metformin action: Concentrations matter. Cell Metab. 2015, 21, 159–162. [Google Scholar] [CrossRef]
- Gong, L.; Goswami, S.; Giacomini, K.M.; Altman, R.B.; Klein, T.E. Metformin pathways: Pharmacokinetics and pharmacodynamics. Pharmacogenet. Genom. 2012, 22, 820. [Google Scholar] [CrossRef] [PubMed]
- Vecchio, S.; Giampreti, A.; Petrolini, V.; Lonati, D.; Protti, A.; Papa, P. Metformin accumulation: Lactic acidosis and high plasmatic metformin levels in a retrospective case series of 66 patients on chronic therapy. Clin. Toxicol. 2014, 52, 129–135. [Google Scholar] [CrossRef]
- Duong, J.K.; Kumar, S.S.; Kirkpatrick, C.M.; Greenup, L.C.; Arora, M.; Lee, T.C. Population pharmacokinetics of metformin in healthy subjects and patients with type 2 diabetes mellitus: Simulation of doses according to renal function. Clin. Pharmacokinet. 2013, 52, 373–384. [Google Scholar] [CrossRef]
- Foretz, M.; Guigas, B.; Viollet, B. Metformin: Update on mechanisms of action and repurposing potential. Nat. Rev. Endocrinol. 2023, 19, 460–476. [Google Scholar] [CrossRef] [PubMed]
- Gunton, J.E.; Delhanty, P.J.; Takahashi, S.; Baxter, R.C. Metformin rapidly increases insulin receptor activation in human liver and signals preferentially through insulin-receptor substrate-2. J. Clin. Endocrinol. Metab. 2003, 88, 1323–1332. [Google Scholar] [CrossRef]
- Cusi, K.; Consoli, A.; DeFronzo, R.A. Metabolic effects of metformin on glucose and lactate metabolism in noninsulin-dependent diabetes mellitus. J. Clin. Endocrinol. Metab. 1996, 81, 4059–4067. [Google Scholar]
- Hundal, R.S.; Krssak, M.; Dufour, S.; Laurent, D.; Lebon, V.; Chandramouli, V.; Inzucchi, S.E.; Schumann, W.C.; Petersen, K.F.; Landau, B.R.; et al. Mechanism by which metformin reduces glucose production in type 2 diabetes. Diabetes 2000, 49, 2063–2069. [Google Scholar] [CrossRef]
- Argaud, D.; Roth, H.; Wiernsperger, N.; Leverve, X.M. Metformin decreases gluconeogenesis by enhancing the pyruvate kinase flux in isolated rat hepatocytes. Eur. J. Biochem. 1993, 213, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Large, V.; Beylot, M. Modifications of citric acid cycle activity and gluconeogenesis in streptozotocin-induced diabetes and effects of metformin. Diabetes 1999, 48, 1251–1257. [Google Scholar] [CrossRef]
- Mithieux, G.; Guignot, L.; Bordet, J.C.; Wiernsperger, N. Intrahepatic mechanisms underlying the effect of metformin in decreasing basal glucose production in rats fed a high-fat diet. Diabetes 2002, 51, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Radziuk, J.; Zhang, Z.; Wiernsperger, N.; Pye, S. Effects of metformin on lactate uptake and gluconeogenesis in the perfused rat liver. Diabetes 1997, 46, 1406–1413. [Google Scholar] [CrossRef]
- Wilcock, C.; Bailey, C.J. Accumulation of metformin by tissues of the normal and diabetic mouse. Xenobiotica 1994, 24, 49–57. [Google Scholar] [CrossRef]
- Tobar, N.; Rocha, G.Z.; Santos, A.; Guadagnini, D.; Assalin, H.B.; Camargo, J.A.; Goncalves, A.E.S.S.; Pallis, F.R.; Oliveira, A.G.; Rocco, S.A.; et al. Metformin acts in the gut and induces gut–liver crosstalk. Proc. Natl. Acad. Sci. USA 2023, 120, e2211933120. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J.; Wilcock, C.; Scarpello, J.H. Metformin and the intestine. Diabetologia 2008, 51, 1552–1553. [Google Scholar] [CrossRef]
- Proctor, W.R.; Ming, X.; Bourdet, D.; Han, T.K.; Everett, R.S.; Thakker, D.R. Why does the intestine lack basolateral eflux transporters for cationic compounds? A provocative hypothesis. J. Pharm. Sci. 2016, 105, 484–496. [Google Scholar] [CrossRef]
- Wu, T.; Xie, C.; Wu, H.; Jones, K.L.; Horowitz, M.; Rayner, C.K. Metformin reduces the rate of small intestinal glucose absorption in type 2 diabetes. Diabetes Obes. Metab. 2017, 19, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J. Metformin and intestinal glucose handling. Diabetes Metab. Rev. 1995, 11, S23–S32. [Google Scholar] [CrossRef] [PubMed]
- Zubiaga, L.; Brinad, O.; Auger, F.; Touche, V.; Hubert, T.; Thvenet, J.; Marciniak, C.; Quenon, Q.; Bonner, C.; Peschard, S.; et al. Oral metformin transiently lowers post-prandial glucose response by reducing the apical expression of sodium-glucose co-transporter 1 in enterocytes. iScience 2023, 26, 106057. [Google Scholar] [CrossRef] [PubMed]
- Maida, A.; Lamont, B.J.; Cao, X.; Drucker, D.J. Metformin regulates the incretin receptor axis via a pathway dependent on peroxisome proliferator-activated receptor-α in mice. Diabetologia 2011, 54, 339–349. [Google Scholar] [CrossRef]
- Borg, M.J.; Boung, M.; Grivell, J.; Sun, Z.; Jones, K.L.; Horowitz, M.; Rayner, C.K.; Wu, T. Comparative effects of proximal and distal small intestinal administration of metformin on plasma glucose and glucagon-like peptide-1, and gastric emptying after oral glucose, in type 2 diabetes. Diabetes Obes. Metab. 2019, 21, 640–647. [Google Scholar] [CrossRef]
- Chondronikola, M.; Volpi, E.; Børsheim, E.; Porter, C.; Annamalai, P.; Enerbäck, S.; Lidell, M.E.; Saraf, M.K.; Labbe, S.M.; Hurren, N.M.; et al. Brown adipose tissue improves whole-body glucose homeostasis and insulin sensitivity in humans. Diabetes 2014, 63, 4089–4099. [Google Scholar] [CrossRef]
- Sponton, C.H.; Hosono, T.; Taura, J.; Jedrychowski, M.P.; Yoneshiro, T.; Wang, Q.; Takahashi, M.; Matsui, Y.; Ikeda, K.; Oguri, Y.; et al. The regulation of glucose and lipid homeostasis via PLTP as a mediator of BAT–liver communication. EMBO Rep. 2020, 21, e49828. [Google Scholar] [CrossRef] [PubMed]
- Breining, P.; Jensen, J.B.; Sundelin, E.I.; Gormsen, L.C.; Jakobsen, S.; Busk, M.; Rolighed, L.; Bross, P.; Fernandez-Guerra, P.; Markussen, L.K.; et al. Metformin targets brown adipose tissue in vivo and reduces oxygen consumption in vitro. Diabetes Obes. Metab. 2018, 20, 2264–2273. [Google Scholar] [CrossRef] [PubMed]
- Geerling, J.J.; Boon, M.R.; Van der Zon, G.C.; Van den Berg, S.A.; Van den Hoek, A.M.; Lombès, M.; Princen, H.M.G.; Havekes, L.M.; Rensen, P.C.N.; Guigas, B. Metformin lowers plasma triglycerides by promoting VLDLtriglyceride clearance by brown adipose tissue in mice. Diabetes 2014, 63, 880–891. [Google Scholar] [CrossRef]
- Tokubuchi, I.; Tajiri, Y.; Iwata, S.; Hara, K.; Wada, N.; Hashinaga, T.; Nakayama, H.; Mifune, H.; Yamada, K. Beneficial effects of metformin on energy metabolism and visceral fat volume through a possible mechanism of fatty acid oxidation in human subjects and rats. PLoS ONE 2017, 12, e0171293. [Google Scholar] [CrossRef]
- Riedmaier, A.E.; Nies, A.T.; Schaeffeler, E.; Schwab, M. Organic anion transporters and their implications in pharmacotherapy. Pharmacol. Rev. 2012, 64, 421–449. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.C.; Sobti, M.; Quinn, A.; Smith, N.J.; Brown Simon, H.J.; Vanderberg, J.I.; Ryan, R.M.; O’Mara, M.L.; Stewart, A.G. Structural basis of promiscuous substrate transport by Organic Cation Transporter 1. Nat. Commun. 2023, 14, 6374. [Google Scholar] [CrossRef] [PubMed]
- Dujic, T.; Zhou, K.; Donnelly, L.A.; Tavendale, R.; Palmer, C.N.A.; Pearson, E.R. Association of organic cation transporter 1 with intolerance to metformin in type 2 diabetes: A GoDARTS study. Diabetes 2015, 64, 1786–1793. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Sheardown, S.A.; Brown, C.; Owen, R.P.; Zhang, S.; Castro, R.A.; Ianculescu, A.G.; Yue, L.; Lo, J.C.; Burchard, E.G.; et al. Effect of genetic variation in the organic cation transporter 1 (OCT1) on metformin action. J. Clin. Investig. 2007, 117, 1422–1431. [Google Scholar] [CrossRef]
- Kawoosa, F.; Zafar, A.S.; Masoodi, S.R.; Amin, A.; Rasool, R.; Fazili, K.M.; Dar, A.H.; Lone, A.; Bashir, S.U.I. Role of human organic cation transporter-1 (OCT1/SLC22A1) in modulating the response to metformin in patients with type 2 diabetes. BMC Endocr. Disord. 2022, 22, 140. [Google Scholar] [CrossRef]
- Bridges, H.R.; Blaza, J.N.; Yin, Z.; Chung, I.; Pollak, M.N.; Hirst, J. Structural basis of mammalian respiratory complex I inhibition by medicinal biguanides. Science 2023, 379, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Bridges, H.R.; Jones, A.J.; Pollak, M.N.; Hirst, J. Effects of metformin and other biguanides on oxidative phosphorylation in mitochondria. Biochem. J. 2014, 462, 475–487. [Google Scholar] [CrossRef]
- LaMoia, T.E.; Butrico, G.M.; Kalpage, H.A.; Goedeke, L.; Hubbard, B.T.H.; Vatner, D.F.; Gaspar, R.C.; Zhang, X.M.; Cline, G.W.; Nakahara, K.; et al. Metformin, phenformin, and galegine inhibit complex IV activity and reduce glycerol-derived gluconeogenesis. Proc. Natl. Acad. Sci. USA 2022, 119, e2122287119. [Google Scholar] [CrossRef] [PubMed]
- Madiraju, A.K.; Erion, D.M.; Rahimi, Y.; Zhang, X.M.; Braddock, D.T.; Albright, R.A.; Prigaro, B.J.; Wood, J.L.; Bhanot, S.; MacDonald, M.J.; et al. Metformin suppresses gluconeogenesis by inhibiting mitochondrial glycerophosphate dehydrogenase. Nature 2014, 510, 542–546. [Google Scholar] [CrossRef]
- Ma, T.; Tian, X.; Zhang, B.; Li, M.; Wang, Y.; Yang, C.; Wu, J.; Wei, X.; Qu, Q.; Yu, Y.; et al. Low-dose metformin targets the lysosomal AMPK pathway through PEN2. Nature 2022, 603, 159–165. [Google Scholar] [CrossRef]
- Aljofan, M.; Riethmacher, D. Anticancer activity of metformin: A systematic review of the literature. Future Sci. OA 2019, 5, FSO410. [Google Scholar] [CrossRef]
- Hua, Y.; Zheng, Y.; Yao, Y.; Jia, R.; Ge, S.; Zhuang, A. Metformin and cancer hallmarks: Shedding new lights on therapeutic repurposing. J. Transl. Med. 2023, 21, 403. [Google Scholar] [CrossRef] [PubMed]
- Howell, J.J.; Hellberg, K.; Turner, M.; Talbott, G.; Kolar, M.J.; Ross, D.S.; Hoxhaj, G.; Saghatelian, A.; Shaw, R.J.; Manning, B.D. Metformin Inhibits Hepatic mTORC1 Signaling via Dose-Dependent Mechanisms Involving AMPK and the TSC Complex. Cell Metab. 2017, 25, 463–471. [Google Scholar] [CrossRef]
- Lama-Sherpa, T.D.; Jeong, M.H.; Jewell, J.L. Regulation of mTORC1 by the Rag GTPases. Biochem. Soc. Trans. 2023, 51, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.; Liu, E.; Knutzen, C.A.; Lang, E.S.; Lombardo, C.R.; Sankar, S.; Toth, J.I.; Petroski, M.D.; Ronai, Z.; Chiang, G.G. REDD1, an inhibitor of mTOR signalling, is regulated by the CUL4A-DDB1 ubiquitin ligase. EMBO Rep. 2009, 10, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Sahra, I.; Regazzetti, C.; Robert, G.; Laurent, K.; Le Marchand-Brustel, Y.; Auberger, P.; Tanti, J.F.; Giorgetti-Peraldi, S.; Bost, F. Metformin, Independent of AMPK, Induces mTOR Inhibition and Cell-Cycle Arrest through REDD1. Cancer Res. 2011, 71, 4366–4372. [Google Scholar] [CrossRef]
- Han, P.; Zhou, J.; Xiang, J.; Liu, Q.; Sun, K. Research progress on the therapeutic effect and mechanism of metformin for lung cancer (Review). Oncol. Rep. 2023, 49, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.-W.; Zhou, N.; Jin, F.; Wang, R.; Zhao, J.-Q. Metformin reduces pancreatic cancer cell proliferation and increases apoptosis through MTOR signaling pathway and its dose-effect relationship. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5336–5344. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Cui, J.; Wang, H.; Medina, R.; Zhang, S.; Zhang, X.; Zhuang, Z.; Lin, Y. Metformin enhances anti-cancer effects of cisplatin in meningioma through AMPK-mTOR signaling pathways. Mol. Ther.-Oncolytics 2021, 20, 119–131. [Google Scholar] [CrossRef]
- Grimaldi, C.; Chiarini, F.; Tabellini, G. Amp-dependent kinase/mammalian target of rapamycin complex 1 signaling in t-cell acute lymphoblastic leukemia: Therapeutic implications. Leukemia 2012, 26, 91–100. [Google Scholar] [CrossRef]
- Green, A.S.; Chapuis, N.; Maciel, T.T. The lkb1/ampk signaling pathway has tumor suppressor activity in acute myeloid leukemia through the repression of mtor-dependent oncogenic mrna translation. Blood 2010, 116, 4262–4273. [Google Scholar] [CrossRef]
- Rocha, G.Z.; Dias, M.M.; Ropelle, E.R. Metformin amplifies chemotherapy-induced ampk activation and antitumoral growth. Clin. Cancer Res. 2011, 17, 3993–4005. [Google Scholar] [CrossRef]
- Shi, W.Y.; Xiao, D.; Wang, L. Therapeutic metformin/ampk activation blocked lymphoma cell growth via inhibition of mTOR pathway and induction of autophagy. Cell Death Dis. 2012, 3, e275. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Yi, K.H.; Han, S.K. Therapeutic potential of metformin in papillary thyroid cancer in vitro and in vivo. Mol. Cell. Endocrinol. 2014, 393, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Glaviano, A.; Foo, A.S.C.; Lam, H.Y. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol. Cancer 2023, 22, 138. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Xue, D.; Wang, K.; Zhang, F.; Shi, J.; Jia, B.; Yang, D.; Zhang, Q.; Zhang, S.; Jiang, H.; et al. Metformin exerts an antitumor effect by inhibiting bladder cancer cell migration and growth, and promoting apoptosis through the PI3K/AKT/mTOR pathway. BMC Urol. 2022, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Nozhat, Z.; Mohammadi-Yeganeh, S.; Azizi, F.; Zarkesh, M.; Hedayati, M. Effects of metformin on the PI3K/AKT/FOXO1 pathway in anaplastic thyroid cancer cell lines. DARU 2018, 26, 93–103. [Google Scholar] [CrossRef]
- Tang, J.C.; An, R.; Jiang, Y.Q.; Yang, J. Effects and Mechanisms of Metformin on the Proliferation of Esophageal Cancer Cells In Vitro and In Vivo. Cancer Res. Treat. 2017, 49, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Liu, C.; He, Z.; Cai, Y.; Chen, J. Metformin inhibits cervical cancer cell proliferation by modulating PI3K/Akt-induced major histocompatibility complex class I-related chain A gene expression. J. Exp. Clin. Cancer Res. 2020, 39, 127. [Google Scholar] [CrossRef]
- Xiao, Q.; Xiao, J.; Liu, J. Metformin suppresses the growth of colorectal cancer by targeting INHBA to inhibit TGF-β/PI3K/AKT signaling transduction. Cell Death Dis. 2022, 13, 202. [Google Scholar] [CrossRef]
- Cui, Y.; Zhou, J.; Rong, F. Combination of metformin and RG7388 enhances inhibition of growth and induction of apoptosis of ovarian cancer cells through the PI3K/AKT/mTOR pathway. Biochem. Biophys. Res. Commun. 2020, 533, 665–671. [Google Scholar] [CrossRef]
- Qiang, P.; Shao, Y.; Sun, Y.-P.; Zhang, J.; Chen, L.-J. Metformin inhibits proliferation and migration of endometrial cancer cells through regulating PI3K/AKT/MDM2 pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1778–1785. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhu, W.; Yang, B.; Chai, R.; Liu, T.; Li, F.; Ren, G.; Ji, S.; Liu, S.; Li, G. The cotreatment of metformin with flavone synergistically induces apoptosis through inhibition of PI3K/AKT pathway in breast cancer cells. Oncol. Lett. 2018, 15, 5952–5958. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhai, R.; Ma, C.; Miao, W. Combination of aloin and metformin enhances the antitumor effect by inhibiting the growth and invasion and inducing apoptosis and autophagy in hepatocellular carcinoma through PI3K/AKT/mTOR pathway. Cancer Med. 2020, 9, 1141–1151. [Google Scholar] [CrossRef]
- Iglesias, D.A.; Yates, M.S.; van der Hoeven, D.; Rodkey, T.L.; Zhang, Q.; Co, N.N.; Burzawa, J.; Chigurupati, S.; Celestino, J.; Bowser, J.; et al. Another surprise from Metformin: Novel mechanism of action via K-Ras influences endometrial cancer response to therapy. Mol. Cancer Ther. 2013, 12, 2847–2856. [Google Scholar] [CrossRef]
- Xie, J.; Xia, L.; Xiang, W.; He, W.; Yin, H.; Wang, F.; Gao, T.; Qi, W.; Yang, Z.; Yang, X.; et al. Metformin selectively inhibits metastatic colorectal cancer with the KRAS mutation by intracellular accumulation through silencing MATE1. Proc. Natl. Acad. Sci. USA 2020, 117, 13012–13022. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Guo, F.; Wang, W.; Shi, H.; Li, D.; Wang, Y. K-ras gene mutation as a predictor of cancer cell responsiveness to metformin. Mol. Med. Rep. 2013, 8, 763–768. [Google Scholar] [CrossRef]
- Dong, S.; Zeng, L.; Liu, Z.; Li, R.; Zou, Y.; Li, Z.; Ge, C.; Lai, Z.; Xue, Y.; Yang, J.; et al. NLK functions to maintain proliferation and stemness of NSCLC and is a target of metformin. J. Hematol. Oncol. 2015, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, U.K.; Kini, S.G.; Garg, V.; Agrawal, S.; Tomar, P.K.; Pathak, P.; Chaudhary, A.; Gupta, P.; Malik, A. JNK pathway signaling: A novel and smarter therapeutic targets for various biological diseases. Future Med. Chem. 2015, 7, 2065–2086. [Google Scholar] [CrossRef]
- Li, B.; Zhou, P.; Xu, K.; Chen, T.; Jiao, J.; Wei, H.; Yang, X.; Xu, W.; Wan, W.; Xiao, J. Metformin induces cell cycle arrest, apoptosis and autophagy through ROS/JNK signaling pathway in human osteosarcoma. Int. J. Biol. Sci. 2020, 16, 74–84. [Google Scholar] [CrossRef]
- Wu, N.; Gu, C.; Gu, H.; Hu, H.; Han, Y.; Li, Q. Metformin induces apoptosis of lung cancer cells through activating JNK/p38 MAPK pathway and GADD153. Neoplasma 2011, 58, 482–490. [Google Scholar] [CrossRef]
- Lu, C.; Chiang, J.; Tsai, F.; Hsu, Y.; Juan, Y.; Yang, J.; Chiu, H. Metformin triggers the intrinsic apoptotic response in human AGS gastric adenocarcinoma cells by activating AMPK and suppressing mTOR/AKT signaling. Int. J. Oncol. 2019, 54, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cao, L.; Wang, L.; Liu, L.; Huang, Y.; Gong, X. Metformin Inhibits Proliferation of Human Thyroid Cancer TPC-1 Cells by Decreasing LRP2 to Suppress the JNK Pathway. OncoTargets Ther. 2020, 13, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Montalto, F.I.; De Amicis, F. Cyclin D1 in Cancer: A Molecular Connection for Cell Cycle Control, Adhesion and Invasion in Tumor and Stroma. Cells 2020, 9, 2648. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ke, C.; Tang, Q. Metformin promotes autophagy and apoptosis in esophageal squamous cell carcinoma by downregulating Stat3 signaling. Cell Death Dis. 2014, 5, e1088. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.-S.; Wang, S.; Deng, A.; Liu, B.; Edgerton, S.; Lind, S.; Wahdan-Alaswad, R.; Thor, A. Metformin targets Stat3 to inhibit cell growth and induce apoptosis in triple-negative breast cancers. Cell Cycle 2012, 11, 367–376. [Google Scholar] [CrossRef]
- Wallbillich, J.J.; Josyula, S.; Saini, U.; Zingarelli, R.A.; Dorayappan, K.D.P. High Glucose-Mediated STAT3 Activation in Endometrial Cancer Is Inhibited by Metformin: Therapeutic Implications for Endometrial Cancer. PLoS ONE 2017, 12, e0170318. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, M.; Zhang, Q. Metformin Antagonizes Ovarian Cancer Cells Malignancy Through MSLN Mediated IL-6/STAT3 Signaling. Cell Transplant. 2021, 30, 9636897211027819. [Google Scholar] [CrossRef]
- Saengboonmee, C.; Seubwai, W.; Cha’on, U.; Sawanyawisuth, K.; Wongkham, S.; Wongkham, C. Metformin Exerts Antiproliferative and Anti-metastatic Effects Against Cholangiocarcinoma Cells by Targeting STAT3 and NF-ĸB. Anticancer Res. 2017, 37, 115–123. [Google Scholar] [CrossRef]
- Fonseca, N.P.; Fenerich, B.A.; Pereira-Martins, D.A.; Garibaldi, P.M.M.; Coelho-Silva, J.L.; Alves Silva, A.B.; Fernandes, J.C.; Silva, C.L.; Rego, M.E.; Machado-Neto, J.A.; et al. Metformin Inhibits STAT3 and MAPK Signaling through AMPK Activation in CSF3RT618I Cells. Blood 2022, 140 (Suppl. 1), 12153–12154. [Google Scholar] [CrossRef]
- Leidgens, V.; Proske, J.; Rauer, L.; Moeckel, S.; Renner, K.; Bogdahn, U.; Riemenschneider, M.J.; Proescholdt, M.; Vollmann-Zwerenz, A.; Hau, P.; et al. Stattic and metformin inhibit brain tumor initiating cells by reducing STAT3-phosphorylation. Oncotarget 2017, 8, 8250–8263. [Google Scholar] [CrossRef]
- Pan, Q.; Yang, G.-L.; Yang, J.-H.; Lin, S.-L.; Liu, N.; Liu, S.-S.; Liu, M.-Y.; Zhang, L.-H.; Huang, Y.-R.; Shen, R.L.; et al. Metformin can block precancerous progression to invasive tumors of bladder through inhibiting STAT3-mediated signaling pathways. J. Exp. Clin. Cancer Res. 2015, 34, 77. [Google Scholar] [CrossRef] [PubMed]
- Esparza-López, J.; Alvarado-Muñoz, J.F.; Escobar-Arriaga, E. Metformin reverses mesenchymal phenotype of primary breast cancer cells through STAT3/NF-κB pathways. BMC Cancer 2019, 19, 728. [Google Scholar] [CrossRef]
- Bashraheel, S.S.; Kheraldine, H.; Khalaf, S.; Al Moustafa, A.-E. Metformin and HER2-positive breast cancer: Mechanisms and therapeutic implications. Biomed. Pharmacother. 2023, 162, 114676. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kim, M.Y.; Hong, S. Synergistic Effects of Metformin and Trastuzumab on HER2 Positive Gastroesophageal Adenocarcinoma Cells In Vitro and In Vivo. Cancers 2023, 15, 4768. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kwon, H.; Lee, J.W. Metformin increases survival in hormone receptor-positive, HER2-positive breast cancer patients with diabetes. Breast Cancer Res. 2015, 17, 64. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Martin, A.; Oliveras-Ferraros, C.; Menendez, J.A. The antidiabetic drug metformin suppresses HER2 (erbB-2) oncoprotein overexpression via inhibition of the mTOR effector p70S6K1 in human breast carcinoma cells. Cell Cycle 2009, 8, 88–96. [Google Scholar] [CrossRef]
- Chen, T.W.; Liang, Y.N.; Feng, D.; Tao, L.Y.; Qi, K.; Zhang, H.Y.; Wang, H.X.; Lin, Q.S.; Kong, H. Metformin inhibits proliferation and promotes apoptosis of HER2 positive breast cancer cells by downregulating HSP90. J. Balk. Union Oncol. 2013, 18, 51–56. [Google Scholar]
- Wang, J.; Li, G.; Wang, Y.; Tang, S.; Sun, X.; Feng, X.; Li, Y.; Bao, G.; Li, P.; Mao, X.; et al. Suppression of tumor angiogenesis by metformin treatment via a mechanism linked to targeting of HER2/HIF-1α/VEGF secretion axis. Oncotarget 2015, 6, 44579–44592. [Google Scholar] [CrossRef]
- Tanaka, K.; Babic, I.; Nathanson, D.; Akhavan, D.; Guo, D.; Gini, B. Oncogenic EGFR signaling activates an mTORC2-NF-kappaB pathway that promotes chemotherapy resistance. Cancer Discov. 2011, 1, 524–538. [Google Scholar] [CrossRef]
- Li, L.; Wang, T.; Hu, M.; Zhang, Y.; Chen, H.; Xu, L. Metformin Overcomes Acquired Resistance to EGFR TKIs in EGFR-Mutant Lung Cancer via AMPK/ERK/NF-κB Signaling Pathway. Front. Oncol. 2020, 10, 1605. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Bian, Y.; Zhang, Y.; Ren, G.; Li, G. Metformin activates AMPK/SIRT1/NF-κB pathway and induces mitochondrial dysfunction to drive caspase3/GSDME-mediated cancer cell pyroptosis. Cell Cycle 2020, 19, 1089–1104. [Google Scholar] [CrossRef] [PubMed]
- Yenmis, G.; Yaprak Sarac, E.; Besli, N.; Soydas, T.; Tastan, C.; Dilek Kancagi, D.; Yilanci, M.; Senol, K.; Karagulle, O.O.; Ekmekci, C.G.; et al. Anti-cancer effect of metformin on the metastasis and invasion of primary breast cancer cells through mediating NF-kB activity. Acta Histochem. 2021, 123, 151709. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, X.; Chen, Z.; Chen, H.; Tan, Y. Metformin Suppressed Tumor Necrosis Factor-α-Induced Epithelial- Mesenchymal Transition in Prostate Cancer by Inactivating the NF-κB Signaling Pathway. Transl. Cancer Res. 2020, 9, 6086–6095. Available online: https://tcr.amegroups.org/article/view/44883 (accessed on 17 October 2023). [CrossRef] [PubMed]
- Xu, S.; Yang, Z.; Jin, P.; Yang, X.; Li, X.; Wei, X.; Wang, Y.; Long, S.; Zhang, T.; Chen, G.; et al. Metformin Suppresses Tumor Progression by Inactivating Stromal Fibroblasts in Ovarian Cancer. Mol. Cancer Ther. 2018, 17, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Yi, Y.; Liu, Y.; Liu, X.; Keller, E.T.; Qian, C.N.; Zhang, J.; Lu, Y. Metformin targets multiple signaling pathways in cancer. Chin. J. Cancer 2017, 36, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.-X.; Wang, H.; Zeng, Z.; Li, X.-M. Metformin Down-regulates Endometrial Carcinoma Cell Secretion of IGF-1 and Expression of IGF-1R. Asian Pac. J. Cancer Prev. 2015, 16, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Chen, F.; Yue, F.; Camacho, L.; Kothapalli, S.; Wei, G.; Huang, S.; Mo, Q.; Ma, F.; Li, Y.; et al. Metformin and an insulin/IGF-1 receptor inhibitor are synergistic in blocking growth of triple-negative breast cancer. Breast Cancer Res. Treat. 2021, 185, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Esposito, A.I.; Amaro, A.; Angelini, G.; Emionite, L.; Gennari, A.; Indraccolo, S.; Maggi, D.; Marini, C.; Salani, B.; Sambuceti, G.; et al. Abstract 1182: Metformin affects breast cancer cell growth and disturbs an IGF1/insulin related gene network that correlates with breast cancer progression. Cancer Res. 2015, 75 (Suppl. 15), 1182. [Google Scholar] [CrossRef]
- UK Prospective Diabetes Study (UKPDS) Group. Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33). Lancet 1998, 352, 837–853. [Google Scholar] [CrossRef]
- Home, P.D. Impact of the UKPDS—An overview. Diabet. Med. 2008, 25 (Suppl. 2), 2–8. [Google Scholar] [CrossRef]
- UK Prospective Diabetes Study (UKPDS). VIII. Study design, progress and performance. Diabetologia 1991, 34, 877–890.
- Metropolitan Life Insurance Company. New weight standards for men and women. Stat. Bull. Metrop. Insur. Co. 1959, 40, 1–4. [Google Scholar]
- Holman, R.R.; Paul, S.K.; Bethel, M.A.; Matthews, D.R.; Neil, H.A.W. 10-year follow-up of intensive glucose control in type 2 diabetes. N. Engl. J. Med. 2008, 359, 1577–1589. [Google Scholar] [CrossRef] [PubMed]
- Maruthur, N.M.; Tseng, E.; Hutfless, S.; Wilson, L.M.; Suarez-Cuervo, C.; Berger, Z.; Chu, Y.; Iyoha, E.; Segal, J.B.; Bolen, S. Diabetes Medications as Monotherapy or Metformin-Based Combination Therapy for Type 2 Diabetes: A Systematic Review and Meta-analysis. Ann. Intern. Med. 2016, 164, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Out, M.; Kooy, A.; Lehert, P.; Schalkwijk, C.A.; Stehouwer, C.D.A. Long-term treatment with metformin in type 2 diabetes and methylmalonic acid: Post hoc analysis of a randomized controlled 4.3-year trial. J. Diabetes Complicat. 2018, 32, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Aroda, V.R.; Edelstein, S.L.; Goldberg, R.B.; Knowler, W.C.; Marcovina, S.M.; Orchard, T.J.; Bray, G.A.; Schade, D.S.; Temprosa, M.G.; White, N.H.; et al. Diabetes Prevention Program Research Group. Long-term metformin use and vitamin B12 deficiency in the Diabetes Prevention Program Outcomes Study. J. Clin. Endocrinol. Metab. 2016, 101, 1754–1761. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, X.; Zheng, Y.; Li, J.; Wang, Y.; Lin, Y.; He, Z.; Zhao, W.; Chen, C.; Qiu, K.; et al. Association of glucose-lowering medications with cardiovascular outcomes: An umbrella review and evidence map. Lancet Diabetes Endocrinol. 2020, 8, 192–205. [Google Scholar] [CrossRef]
- Diabetes Prevention Program Research Group. Long-term effects of lifestyle intervention or metformin on diabetes development and microvascular complications over 15-year follow-up: The Diabetes Prevention Program Outcomes Study. Lancet Diabetes Endocrinol. 2015, 3, 866–875. [Google Scholar] [CrossRef]
- Patel, D.; Ayesha, I.E.; Monson, N.R.; Klair, N.; Patel, U.; Saxena, A.; Hamid, P. The Effectiveness of Metformin in Diabetes Prevention: A Systematic Review and Meta-Analysis. Cureus 2023, 15, e46108. [Google Scholar] [CrossRef]
- American Diabetes Association. Pharmacologic Approaches to Glycemic Treatment: Standards of Care in Diabetes—2023. Diabetes Care 2023, 46 (Suppl. 1), S140–S157. [Google Scholar] [CrossRef]
- Meng, H.; Zhang, A.; Liang, Y.; Hao, J.; Zhang, X.; Lu, J. Effect of metformin on glycaemic control in patients with type 1 diabetes: A meta-analysis of randomized controlled trials. Diabetes Metab. Res. Rev. 2018, 34, e2983. [Google Scholar] [CrossRef] [PubMed]
- Petrie, J.R.; Chaturvedi, N.; Ford, I.; Brouwers, M.C.G.J.; Greenlaw, N.; Tillin, T.; Hramiak, I.; Hughes, A.D.; Jenkins, A.J.; Klein, B.E.K.; et al. Cardiovascular and metabolic effects of metformin in patients with type 1 diabetes (REMOVAL): A double-blind, randomised, placebo controlled trial. Lancet Diabetes Endocrinol. 2017, 5, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Miranda, V.C.; Braghiroli, M.I.; Faria, L.D.; Bariani, G.; Alex, A.; Bezerra Neto, J.E.; Capareli, F.C.; Sabbaga, J.; Lobo Dos Santos, J.F.; Hoff, P.M.; et al. Phase 2 Trial of Metformin Combined with 5-Fluorouracil in Patients with Refractory Metastatic Colorectal Cancer. Clin. Color. Cancer 2016, 15, 321–328.e1. [Google Scholar] [CrossRef]
- Akce, M.; Farran, B.; Switchenko, J.M.; Rupji, M.; Kang, S.; Khalil, L.; Ruggieri-Joyce, A.; Olson, B.; Shaib, W.L.; Wu, C.; et al. Phase II trial of nivolumab and metformin in patients with treatment-refractory microsatellite stable metastatic colorectal cancer. J. Immunother. Cancer 2023, 11, e007235. [Google Scholar] [CrossRef] [PubMed]
- Vernieri, C.; Galli, F.; Ferrari, L.; Marchetti, P.; Lonardi, S.; Maiello, E.; Iaffaioli, R.V.; Zampino, M.G.; Zaniboni, A.; De Placido, S.; et al. Impact of Metformin Use and Diabetic Status During Adjuvant Fluoropyrimidine-Oxaliplatin Chemotherapy on the Outcome of Patients with Resected Colon Cancer: A TOSCA Study Subanalysis. Oncologist 2019, 24, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, P.J.; Chen, B.E.; Gelmon, K.A.; Whelan, T.J.; Ennis, M.; Lemieux, J.; Ligibel, J.A.; Hershman, D.L.; Mayer, I.A.; Hobday, T.J.; et al. Effect of Metformin vs Placebo on Invasive Disease-Free Survival in Patients with Breast Cancer: The MA.32 Randomized Clinical Trial. JAMA 2022, 327, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Essa, N.M.; Salem, H.F.; Elgendy, M.O.; Gabr, A.; Omran, M.M.; Hassan, N.A.; Tashkandi, H.M.; Harakeh, S.; Boshra, M.S. Efficacy of Metformin as Adjuvant Therapy in Metastatic Breast Cancer Treatment. J. Clin. Med. 2022, 11, 5505. [Google Scholar] [CrossRef] [PubMed]
- Salah, H.; Rabea, H.; Hassan, A.; Elberry, A.A. Metformin as an Adjuvant Treatment in Non-Diabetic Metastatic Breast Cancer. Bahrain Med. Bull. 2021, 43, 2050312119865114. [Google Scholar]
- Cuyàs, E.; Buxó, M.; Ferri Iglesias, M.J.; Verdura, S.; Pernas, S.; Dorca, J.; Álvarez, I.; Martínez, S.; Pérez-Garcia, J.M.; Batista-López, N.; et al. The C Allele of ATM rs11212617 Associates with Higher Pathological Complete Remission Rate in Breast Cancer Patients Treated with Neoadjuvant Metformin. Front. Oncol. 2019, 28, 193. [Google Scholar] [CrossRef]
- Othman, A.S.; Fatehy, H.; Khadiga, M.A.; Nasef, K.; Hussein, O.; Elkodary, T.R. Neoadjuvant chemotherapy with or without metformin in invasive nonmetastatic breast cancer. Randomized controlled trial. Eur. Chem. Bull. 2023, 12, 7543–7550. [Google Scholar] [CrossRef]
- Azazy, H.A.; Gado, N.M.; Salem, D.A.; El-Ghamry, W.R. Metformin with neoadjuvant chemotherapy in stage II-III breast cancer: A phase II clinical trial. Ann. Oncol. 2020, 31 (Suppl. 4), S323–S324. [Google Scholar] [CrossRef]
- El-Khayat, S.M.; Abouegylah, M.; Abdallah, D.; Geweil, A.G.; Elenbaby, A.M.; Zahra, O.S. The effect of metformin when combined with neoadjuvant chemotherapy in breast cancer patients. Med. Oncol. 2021, 39, 1. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Tong, Y.; Hong, J.; Huang, O.; Wu, J.; He, J.; Chen, W.; Li, Y.; Chen, X.; Shen, K. Neoadjuvant docetaxel, epirubicin, and cyclophosphamide with or without metformin in breast cancer patients with metabolic abnormality: Results from the randomized Phase II NeoMET trial. Breast Cancer Res. Treat. 2023, 197, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, I.; Lohmann, A.E.; Ennis, M.; Dowling, R.J.O.; Cescon, D.; Elser, C.; Potvin, K.R.; Haq, R.; Hamm, C.; Chang, M.C.; et al. A phase II randomized clinical trial of the effect of metformin versus placebo on progression-free survival in women with metastatic breast cancer receiving standard chemotherapy. Breast 2019, 48, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Nanni, O.; Amadori, D.; De Censi, A.; Rocca, A.; Freschi, A.; Bologna, A.; Gianni, L.; Rosetti, F.; Amaducci, L.; Cavanna, L.; et al. Metformin plus chemotherapy versus chemotherapy alone in the first-line treatment of HER2-negative metastatic breast cancer. The MYME randomized, phase 2 clinical trial. Breast Cancer Res. Treat. 2019, 174, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gong, C.; Wang, Z.; Zhang, J.; Wang, L.; Zhang, S.; Cao, J.; Tao, Z.; Li, T.; Wang, B.; et al. A randomized phase II study of aromatase inhibitors plus metformin in pre-treated postmenopausal patients with hormone receptor positive metastatic breast cancer. Oncotarget 2017, 8, 84224–84236. [Google Scholar] [CrossRef] [PubMed]
- Rothermundt, C.A.; Cathomas, R.; Gysel, K.; Fischer, N.; Pereira Mestre, N.; Hermanns, T.; Rothschild, S.I.; Mach, N.; Mingrone, W.; Ciriolo, M.; et al. SAKK 08/14—IMPROVE Investigation of metformin in patients with metastatic castration-resistant prostate cancer (mCRPC) in combination with enzalutamide vs. enzalutamide alone. A randomized, open label, phase II trial. Ann. Oncol. 2022, 33 (Suppl. 7), S808–S869. [Google Scholar] [CrossRef]
- Martin, P.J.; Borchiellini, D.; Guillot, B.T.A.; Paoli, J.-B.; Besson, D.; Hilgers, W.; Priou, F.; El Kouri, C.; Hoch, B.; Deville, J.L.; et al. TAXOMET: A French prospective multicentric randomized controlled phase II study comparing docetaxel plus metformin versus docetaxel plus placebo in mCRPC. Clin. Genitourin. Cancer 2021, 19, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Alghandour, R.; Ebrahim, M.; Elshal, A.; Ghobrial, F.; Elzaafarany, M.; ELbaiomy, A.M. Repurposing metformin as anticancer drug: Randomized controlled trial in advanced prostate cancer (MANSMED). Urol. Oncol. Semin. Orig. Investig. 2021, 39, 831.e1–831.e10. [Google Scholar] [CrossRef]
- Kitson, S.J.; Maskell, Z.; Sivalingam, V.N.; Allen, J.L.; Ali, S.; Burns, S.; Gilmour, K.; Latheef, R.; Slade, R.J.; Pemberton, P.W.; et al. PRE-surgical Metformin in Uterine Malignancy (PREMIUM): A Multi-Center, Randomized Double-Blind, Placebo-Controlled Phase III Trial. Clin. Cancer Res. 2019, 25, 2424–2432. [Google Scholar] [CrossRef]
- Mitsuhashi, A.; Sato, Y.; Kiyokawa, T.; Koshizaka, M.; Hanaoka, H.; Shozu, M. Phase II study of medroxyprogesterone acetate plus metformin as a fertility-sparing treatment for atypical endometrial hyperplasia and endometrial cancer. Ann. Oncol. 2016, 27, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Glasberg, J.; Talans, A.; Giollo, T.R.; Recchimuzzi, D.Z.; Neto, J.E.B.; Lopez, R.V.M.; Hoff, P.M.G.; Riechelmann, R.P. METNET: A phase II trial of metformin in patients with well-differentiated neuroendocrine tumours. Ecancermedicalscience 2022, 16, 1369. [Google Scholar] [CrossRef] [PubMed]
- Hosono, K.; Endo, H.; Takahashi, H.; Sugiyama, M.; Sakai, E.; Uchiyama, T.; Suzuki, K.; Iida, H.; Sakamoto, Y.; Yoneda, K.; et al. Metformin suppresses colorectal aberrant crypt foci in a short-term clinical trial. Cancer Prev. Res. 2010, 3, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Higurashi, T.; Hosono, K.; Takahashi, H.; Komiya, Y.; Umezawa, S.; Sakai, E.; Uchiyama, T.; Taniguchi, L.; Hata, Y.; Uchiyama, S.; et al. Metformin for chemoprevention of metachronous colorectal adenoma or polyps in post-polypectomy patients without diabetes: A multicentre double-blind, placebo-controlled, randomised phase 3 trial. Lancet Oncol. 2016, 17, 475–483. [Google Scholar] [CrossRef]
- Albertelli, M.; Nazzari, E.; Dotto, A.; Grasso, L.F.; Sciallero, S.; Pirchio, R.; Rebora, A.; Boschetti, M.; Pivonello, R.; Ricci Bitti, S.; et al. Possible protective role of metformin therapy on colonic polyps in acromegaly: An exploratory cross-sectional study. Eur. J. Endocrinol. 2021, 184, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Zell, J.A.; McLaren, C.E.; Morgan, T.R.; Lawson, M.J.; Rezk, S.; Albers, C.G.; Chen, W.-P.; Carmichael, J.C.; Chung, J.; Richmond, E.; et al. A Phase IIa Trial of Metformin for Colorectal Cancer Risk Reduction among Individuals with History of Colorectal Adenomas and Elevated Body Mass Index. Cancer Prev. Res. 2020, 13, 203–212. [Google Scholar] [CrossRef]
- Han, J.; Kim, J.H.; Lee, J.W.; Han, S.H.; Kim, H. Neoadjuvant therapy of metformin is associated with good tumor response after preoperative concurrent chemoradiotherapy for rectal cancer. Sci. Rep. 2022, 12, 3716. [Google Scholar] [CrossRef]
- Singh, P.P.; Shi, Q.; Foster, N.R.; Grothey, A.; Nair, S.G.; Chan, E.; Shields, A.F.; Goldberg, R.M.; Gill, S.; Kahlenberg, M.S.; et al. Relationship Between Metformin Use and Recurrence and Survival in Patients with Resected Stage III Colon Cancer Receiving Adjuvant Chemotherapy: Results from North Central Cancer Treatment Group N0147 (Alliance). Oncologist 2016, 21, 1509–1521. [Google Scholar] [CrossRef]
- Available online: https://www.clinicaltrials.gov/study/NCT05921942 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT02614339 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT01905046 (accessed on 18 October 2023).
- Joshua, A.M.; Armstrong, A.; Crumbaker, M.; Scher, H.I.; de Bono, J.; Tombal, B.; Hussain, M.; Sternberg, C.N.; Gillessen, S.; Carles, J.; et al. Statin and metformin use and outcomes in patients with castration-resistant prostate cancer treated with enzalutamide: A meta-analysis of AFFIRM, PREVAIL and PROSPER. Eur. J. Cancer 2022, 170, 285–295. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/study/NCT01864096 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT00268476 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT02511665 (accessed on 18 October 2023).
- Mitsuhashi, A.; Kawasaki, Y.; Hori, M.; Fujiwara, T.; Hanaoka, H.; Shozu, M. Medroxyprogesterone acetate plus metformin for fertility-sparing treatment of atypical endometrial hyperplasia and endometrial carcinoma: Trial protocol for a prospective, randomised, open, blinded-endpoint design, dose-response trial (FELICIA trial). BMJ Open 2020, 10, e035416. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/study/NCT01697566 (accessed on 18 October 2023).
- Available online: https://www.clinicaltrials.gov/study/NCT01797523 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT02065687 (accessed on 18 October 2023).
- Pusceddu, S.; Vernieri, C.; Di Maio, M.; Marconcini, R.; Spada, F.; Massironi, S.; Ibrahim, T.; Brizzi, M.P.; Campana, D.; Faggiano, A.; et al. Metformin use is associated with longer progression-free survival of patients with diabetes and pancreatic neuroendocrine tumors receiving everolimus and/or somatostatin analogues. Gastroenterology 2018, 155, 479–489.e7. [Google Scholar] [CrossRef]
- Available online: https://www.clinicaltrials.gov/study/NCT02294006 (accessed on 27 February 2024).
- Available online: https://www.clinicaltrials.gov/study/NCT02823691 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT05445791 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT02190838 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT03311308 (accessed on 27 February 2024).
- Available online: https://www.clinicaltrials.gov/study/NCT02672488 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT02005419 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT01666730 (accessed on 27 February 2024).
- Yu, O.H.Y.; Suissa, S. Metformin and Cancer: Solutions to a Real-World Evidence Failure. Diabetes Care 2023, 46, 904–912. [Google Scholar] [CrossRef]
- Lord, S.R.; Harris, A.L. Is it still worth pursuing the repurposing of metformin as a cancer therapeutic? Br. J. Cancer 2023, 128, 958–966. [Google Scholar] [CrossRef]
- Wang, P.Y.; Li, J.; Walcott, F.L.; Kang, J.G.; Starost, M.F.; Talagala, S.L.; Zhuang, J.; Park, J.H.; Huffstutler, R.D.; Bryla, C.M.; et al. Inhibiting mitochondrial respiration prevents cancer in a mouse model of Li-Fraumeni syndrome. J. Clin. Investig. 2017, 127, 132–136. [Google Scholar] [CrossRef]
- Panaampon, J.; Zhou, Y.; Saengboonmee, C. Metformin as a booster of cancer immunotherapy. Int. Immunopharmacol. 2023, 121, 110528. [Google Scholar] [CrossRef]
- Amin, D.; Richa, T.; Mollaee, M.; Zhan, T.; Tassone, P.; Johnson, J.; Luginbuhl, A.; Cognetti, D.; Martinez-Outschoorn, U.; Stapp, R.; et al. Metformin Effects on FOXP3+ and CD8+ T Cell Infiltrates of Head and Neck Squamous Cell Carcinoma. Laryngoscope 2020, 130, E490–E498. [Google Scholar] [CrossRef]
- Curry, J.M.; Johnson, J.; Mollaee, M.; Tassone, P.; Amin, D.; Knops, A.; Whitaker-Menezes, D.; Mahoney, M.G.; South, A.; Rodeck, U.; et al. Metformin clinical trial in HPV+ and HPV–head and neck squamous cell carcinoma: Impact on cancer cell apoptosis and immune infiltrate. Front. Oncol. 2018, 8, 436. [Google Scholar] [CrossRef]
- Wang, S.; Lin, Y.; Xiong, X.; Wang, L.; Guo, Y.; Chen, Y.; Chen, S.; Wang, G.; Lin, P.; Chen, H.; et al. Low-Dose Metformin Reprograms the Tumor Immune Microenvironment in Human Esophageal Cancer: Results of a Phase II Clinical Trial. Clin. Cancer Res. 2020, 26, 4921–4932. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Chemical Formula | Chemical Structure | Features and Origin | Solubility LogP | Binding to Mithocondrial Membranes | Tissue of Anaerobic Glycolysis | Metabolism | Risk of Lactic Acidosis (Events for 1000 Subjects/Year) |
|---|---|---|---|---|---|---|---|---|
| GUANINE | C5H5N5O |  | In 1844, the German chemist Julius Bodo Unger obtained it as a mineral formed from the excreta of sea birds (guano). | Insoluble in water | ||||
| GUANIDINE | HNC(NH2)2 |  | It is a strong base, obtained from natural source, via the oxidative degradation of guanine | Soluble in water and ethanol −1.7 | ||||
| METFORMIN | C4H11N5 |  | N,N-dimethylamine guanylguanidine (chemical syntesis) | More hydrophilic −1.43 | Weaker | Mostly intestinal tissue exposed to high drug concentration | Not metabolized, eliminated unchanged | 0.03–0.09 |
| PHENFORMIN | C10H15N5 |  | It was developed in 1957 by Ungar, Freedman, and Shapiro | More lipophilic −0.83 | Stronger | More generalized, including muscle | Not metabolized, eliminated unchanged | 0.40–0.90 |
| BUFORMIN | C6H15N5 |  | It is a strong base, and is freely soluble in water, methanol, and ethanol | Intermediate −1.20 | Stronger | More generalised, including muscle | Not metabolized, eliminated unchanged | >0.1 |
| Trial Title | Tumor | Type of Study | Intervention/Treatment | Patients, n | Primary Outcome (s) | Results | References |
|---|---|---|---|---|---|---|---|
| Phase 2 Trial of Metformin Combined With 5-Fluorouracil in Patients With Refractory Metastatic Colorectal Cancer | Colorectal cancer | Single-arm, phase II study | Metformin 850 mg bid plus 5-FU 425 mg/m2 and leucovorin 50 mg i.v. weekly | 50 | DCR at 8 weeks | Modest activity of metformin plus 5-FU with major benefits observed in patients with BMI ≥ 30 kg/m2 11 (22%) patients presented DCR at 8 weeks with mPFS = 5.6 months and mOS = 16.2 months | Miranda et al., 2016 [140] |
| Phase II trial of nivolumab and metformin in patients with treatment-refractory microsatellite stable metastatic colorectal cancer. | Colorectal cancer | Single-arm, phase II study | Metformin 1000 mg bid plus Nivolumab 480 mg i.v. every 4 weeks | 24 | ORR | No ORR was observed; the study did not proceed with further enrollment | Akce et al., 2023 [141] |
| Impact of Metformin Use and Diabetic Status During Adjuvant Fluoropyrimidine-Oxaliplatin Chemotherapy on the Outcome of Patients with Resected Colon Cancer: A TOSCA Study Subanalysis | Colon cancer | Substudy of phase III TOSCA trial | Metformin plus adjuvant fluoropyrimidine-oxaliplatin | 3759 | OS and RFS | No impact on OS or RFS | Vernieri et al., 2019 [142] |
| Effect of Metformin vs. Placebo on Invasive Disease-Free Survival in Patients With Breast Cancer: The MA.32 Randomized Clinical Trial | Breast cancer | Phase III, placebo-controlled, double-blind RCT | Drug: metformin 850 mg/day for 4 weeks, then 850 mg bid for 5 years plus adjuvant standard breast cancer treatment Control arm: placebo plus adjuvant standard breast cancer treatment | 3649 | Invasive disease-free survival | No significant improvement of the invasive disease-free survival | Goodwin et al., 2022 [143] |
| Efficacy of Metformin as Adjuvant Therapy in Metastatic Breast Cancer Treatment | Breast cancer | Prospective, placebo-controlled RCT | Drug: metformin 500 mg bid plus adjuvant CT Control arm: adjuvant CT | 107 | PFS and RR | No significant benefits on PFS and RR | Essa et al., 2022 [144] |
| Metformin as an Adjuvant Treatment in Non-Diabetic Metastatic Breast Cancer | Breast cancer | Phase II RCT | Drug: metformin 1000 mg bid plus adjuvant CT Control arm: CT | 50 | OS and PFS | Metformin group vs. control group presented no significant improvement of OS and PFS, but higher radiological response (p = 0.002) | Salah et al., 2021 [145] |
| The C Allele of ATM rs11212617 Associates With Higher Pathological Complete Remission Rate in Breast Cancer Patients Treated With Neoadjuvant Metformin (METTEN) | Breast cancer | Phase II, open-label, multicenter RCT | Drug: metformin 850 mg bid for 24 weeks plus anthracycline/taxane-based CT and trastuzumab Control arm: neoadjuvant anthracycline/taxane-based CT and trastuzumab | 79 | PCR | Metformin group was associated with a higher PCR than control group | Cuyàs et al., 2019 [146] |
| Neoadjuvant chemotherapy with or without metformin in invasive nonmetastatic breast cancer. Randomized controlled trial | Breast cancer | Phase II/III RCT | Drug: metformin 850 mg/day, then 850 mg bid plus neoadjuvant CT Control arm: neoadjuvant CT | 140 | Tumor RR | Metformin plus neoadjuvant CT was associated with higher PCR, especially in ER-negative BC (63.2% vs. 22.2%, p = 0.02) and ER-negative BC (50% vs. 34.6%, p = 0.3) | Othman et al., 2023 [147] |
| Metformin with neoadjuvant chemotherapy in stage II-III breast cancer: A phase II clinical trial. | Breast cancer | Phase II RCT | Drug: metformin 850 mg bid plus neoadjuvant CT Control arm: neoadjuvant CT | 60 | OPR | A higher pCR was observed in the metformin group, but without statistical significance (p = 0.09) | Azazy et al., 2020 [148] |
| The effect of metformin when combined with neoadjuvant chemotherapy in breast cancer patients. | Breast cancer | Prospective study | Drug: metformin 850 mg bid plus neoadjuvant CT Control arm: neoadjuvant CT | 59 | PCR | The addition of metformin may improve PCR particularly in individuals with triple-positive BC and BMI ≥ 25 kg/m2 | El-Khayat et al., 2021 [149] |
| Neoadjuvant docetaxel, epirubicin, and cyclophosphamide with or without metformin in breast cancer patients with metabolic abnormality: results from the randomized Phase II NeoMET trial | Breast cancer | Phase II RCT | Drug: metformin 850 mg/day for the first cycle, then 850 mg bid plus neoadjuvant CT (TEC) Control arm: neoadjuvant CT (TEC) | 92 | PCR | No improvement of PCR | Huang et al., 2023 [150] |
| A phase II randomized clinical trial of the effect of metformin versus placebo on progression-free survival in women with metastatic breast cancer receiving standard chemotherapy | Breast cancer | Phase II, double-blind, RCT | Drug: metformin 850 mg bid plus CT Control arm: placebo plus CT | 40 | PFS | No significant impact on PFS | Pimentel et al., 2019 [151] |
| Metformin plus chemotherapy versus chemotherapy alone in the first-line treatment of HER2-negative metastatic breast cancer. The MYME randomized, phase 2 clinical trial. | Breast cancer | Phase II RCT | Drug: metformin 2000 mg/day plus CT Control arm: CT | 122 | PFS | No significant impact on PFS | Nanni et al., 2019 [152] |
| A randomized phase II study of aromatase inhibitors plus metformin in pre-treated postmenopausal patients with hormone receptor positive metastatic breast cancer. | Breast cancer | Phase II RCT | Drug: metformin 500 mg bid plus aromatase inhibitor (exemestane 25 mg/d or letrozole 2.5 mg/d) Control arm: aromatase inhibitor (exemestane 25 mg/d or letrozole 2.5 mg/d) | 60 | PFS | No significant impact on PFS | Zhao et al., 2017 [153] |
| SAKK 08/14—IMPROVE Investigation of metformin in patients with metastatic castration-resistant prostate cancer (mCRPC) in combination with enzalutamide vs. enzalutamide alone. A randomized, open label, phase II trial. | Prostate cancer | Phase II, open-label, multicenter RCT | Drug: metformin 850 mg bid plus enzalutamide 160 mg Control arm: enzalutamide 160 mg | 169 | DCR at 15 months | No benefits on DCR | Rothermundt et al., 2022 [154] |
| TAXOMET: A French prospective multicentric randomized controlled phase II study comparing docetaxel plus metformin versus docetaxel plus placebo in mCRPC. | Prostate cancer | Phase II, placebo-controlled, multicenter RCT | Drug: metformin 850 mg bid plus docetaxel 75 mg/m2 every 21 days plus prednisone 5 mg bid Control arm: placebo plus docetaxel 75 mg/m2 every 21 days plus prednisone 5 mg bid | 99 | PSA response ≥ 50% from baseline | No significant benefits on PSA response, PFS, OS or ORR | Martin et al., 2021 [155] |
| Repurposing metformin as anticancer drug: Randomized controlled trial in advanced prostate cancer (MANSMED) | Prostate cancer | Phase II RCT | Drug: metformin plus standard of care Control arm: standard of care | 124 | CRPC-FS | A significantly higher CRPC-FS (29 months vs. 20 months, p = 0.01) was observed in the metformin group, especially in individuals with high-risk localized disease or metastatic low tumor volume disease | Alghandour et al., 2021 [156] |
| PRE-surgical Metformin In Uterine Malignancy (PREMIUM): a Multi-Center, Randomized Double-Blind, Placebo-Controlled Phase III Trial | Endometrial cancer or AEH | Phase III, multicenter, double-blind RCT | Drug: neoadjuvant metformin 850 mg/d for 3 days, then 850 mg bid for 1 to 5 weeks Control arm: placebo for 1 to 5 weeks until surgery | 88 | Post-treatment IHC expression of Ki-67 | No differences in Ki67 expression were detected | Kitson et al., 2019 [157] |
| Phase II study of medroxyprogesterone acetate plus metformin as a fertility-sparing treatment for atypical endometrial hyperplasia and endometrial cancer. | Endometrial cancer or AEH | Single-arm, phase II trial | Drug: metformin (750–2250 mg/day) plus medroxyprogesterone acetate 400 mg/day | 17 with AEH and 19 with EC | RFS after remission | Beneficial effect of metformin in inhibiting disease relapse (3-year RFS rate = 89%) | Mitsuhashi et al., 2016 [158] |
| METNET: a phase II trial of metformin in patients with well-differentiated neuroendocrine tumours. | GEP-NET or pulmonary NET | Single-arm, phase II trial | Drug: metformin 850 mg bid | 28 | DCR at 6 months | Modest antineoplastic activity of metformin in well-differentiated GEP or lung NET 26 patients had progression, 13 (46%) of whom presented DCR at 6 months and mPFS 6.3 months | Glasberg et al., 2022 [159] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goglia, U.; Hasballa, I.; Teti, C.; Boschetti, M.; Ferone, D.; Albertelli, M. Ianus Bifrons: The Two Faces of Metformin. Cancers 2024, 16, 1287. https://doi.org/10.3390/cancers16071287
Goglia U, Hasballa I, Teti C, Boschetti M, Ferone D, Albertelli M. Ianus Bifrons: The Two Faces of Metformin. Cancers. 2024; 16(7):1287. https://doi.org/10.3390/cancers16071287
Chicago/Turabian StyleGoglia, Umberto, Iderina Hasballa, Claudia Teti, Mara Boschetti, Diego Ferone, and Manuela Albertelli. 2024. "Ianus Bifrons: The Two Faces of Metformin" Cancers 16, no. 7: 1287. https://doi.org/10.3390/cancers16071287
APA StyleGoglia, U., Hasballa, I., Teti, C., Boschetti, M., Ferone, D., & Albertelli, M. (2024). Ianus Bifrons: The Two Faces of Metformin. Cancers, 16(7), 1287. https://doi.org/10.3390/cancers16071287