
Synergistic Effects of the RARalpha Agonist Tamibarotene and the Menin Inhibitor Revumenib in Acute Myeloid Leukemia Cells with KMT2A Rearrangement or NPM1 Mutation

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Cell Lines and Primary AML Cells
2.2. Reagents and Antibodies
2.3. RNA Preparation and RT-qPCR Analysis
2.4. Protein Isolation and Western Blot Analysis
2.5. Flow Cytometry
2.6. Statistics
3. Results
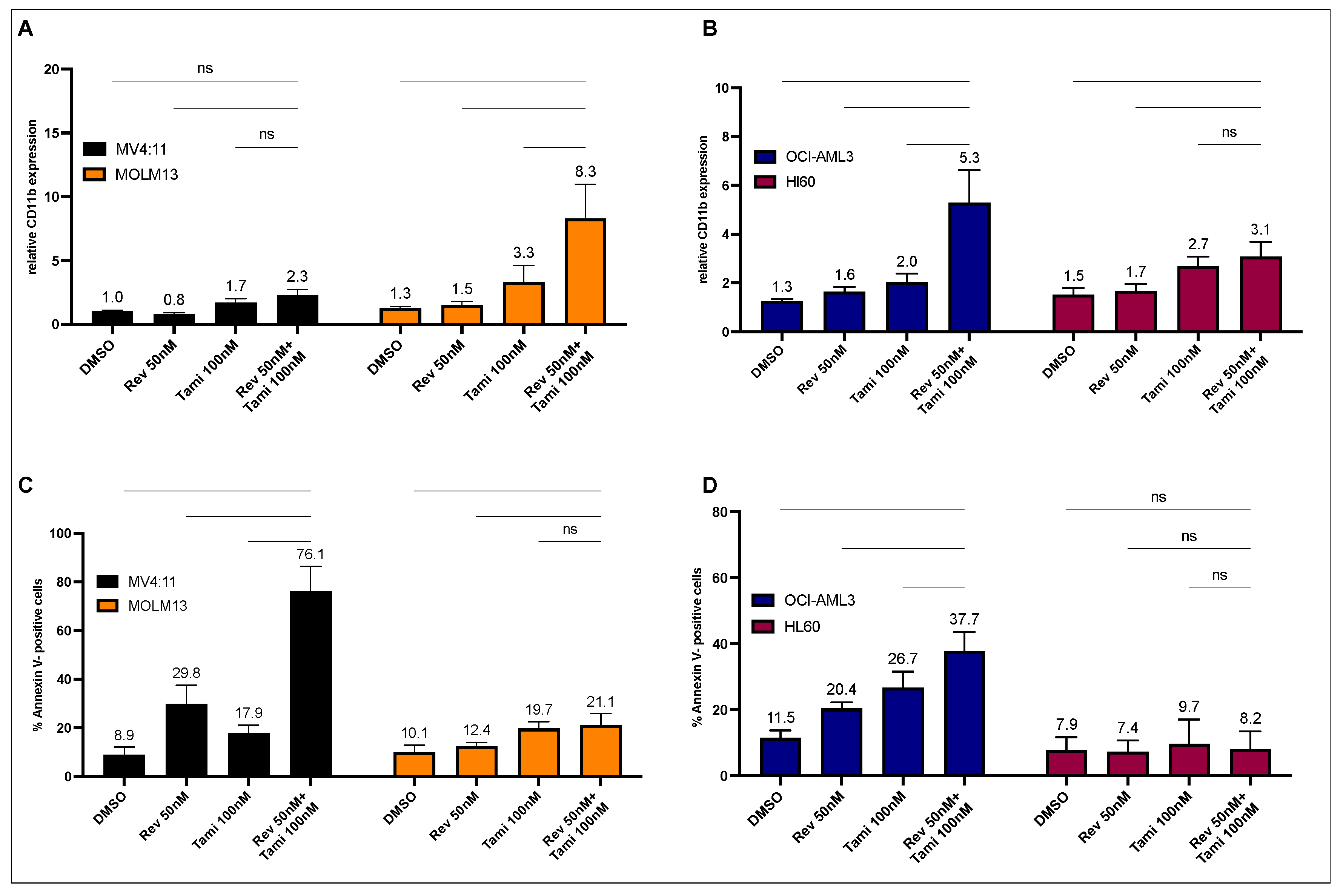
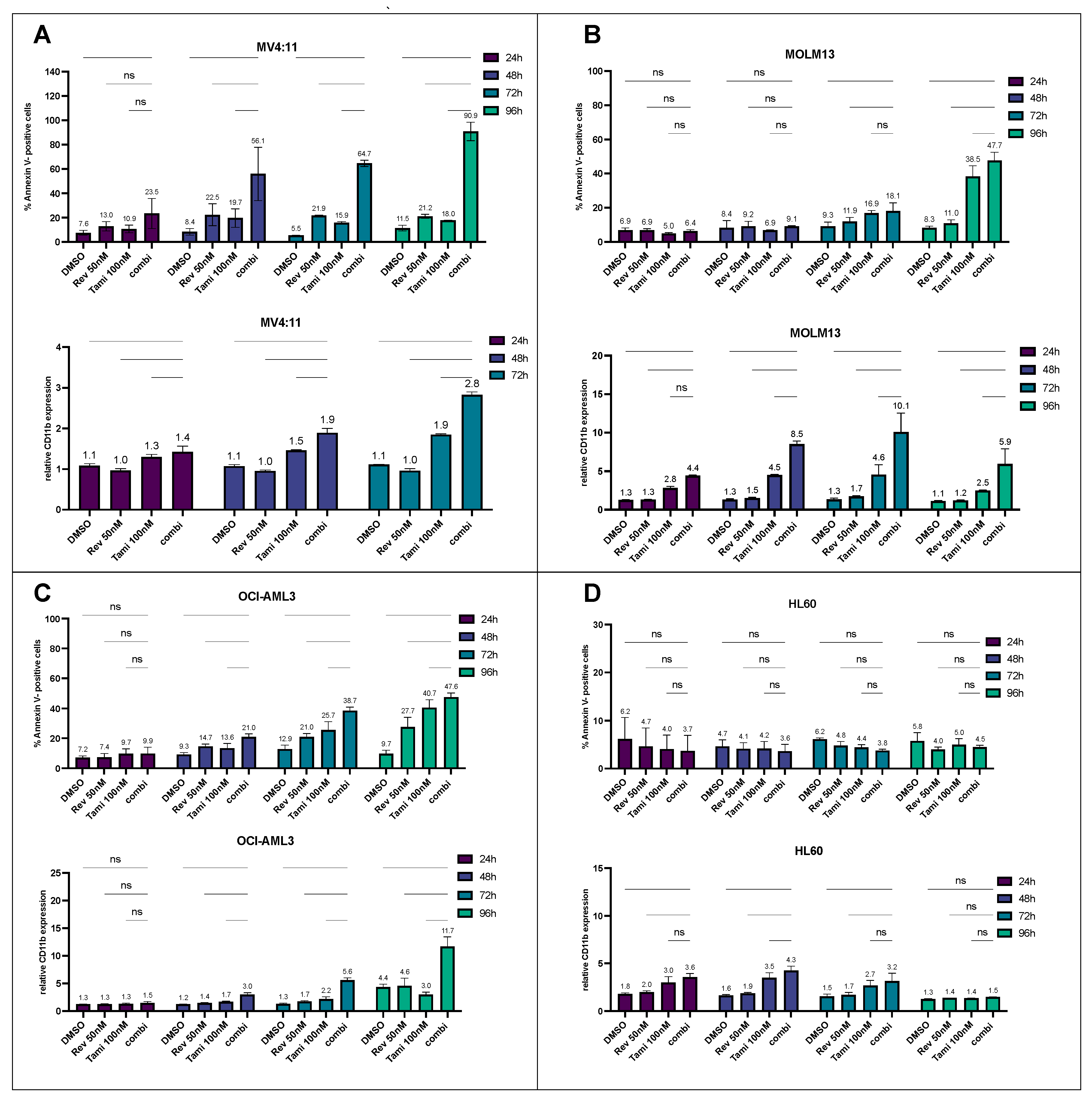
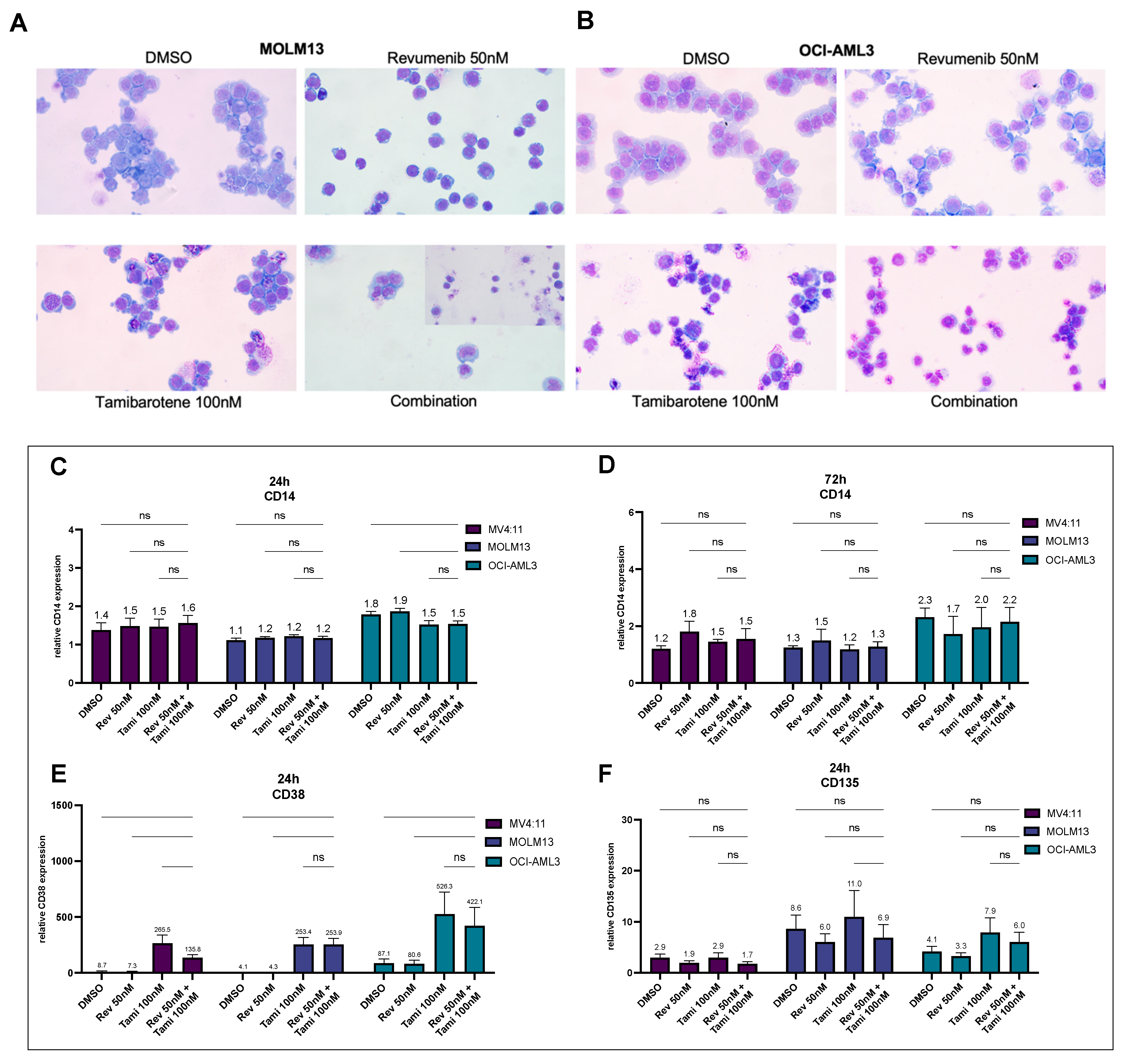
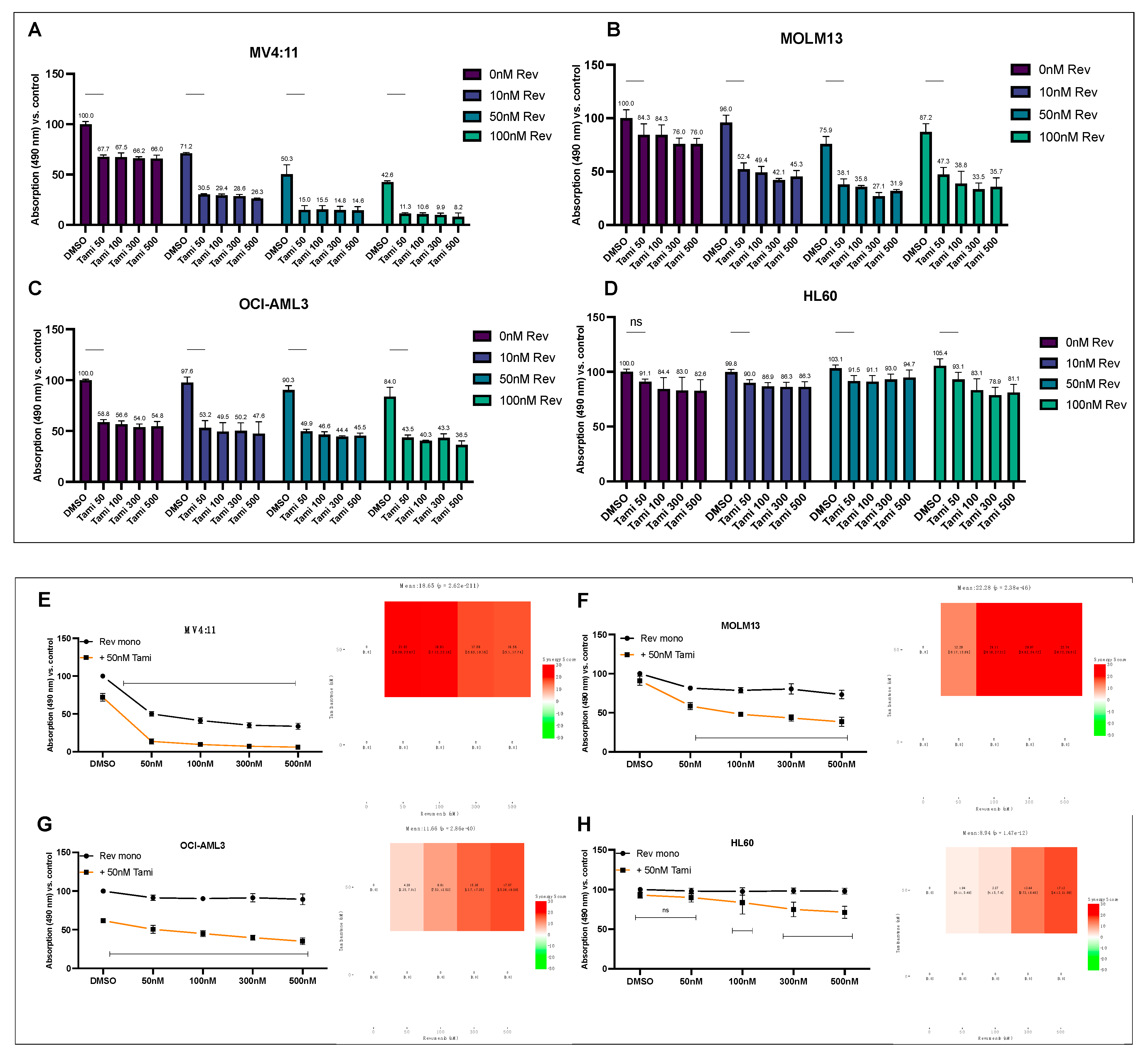
3.1. Determination of Differentiation and Apoptosis Induction after Combination Treatment with Revumenib and Tamibarotene in AML Cell Lines
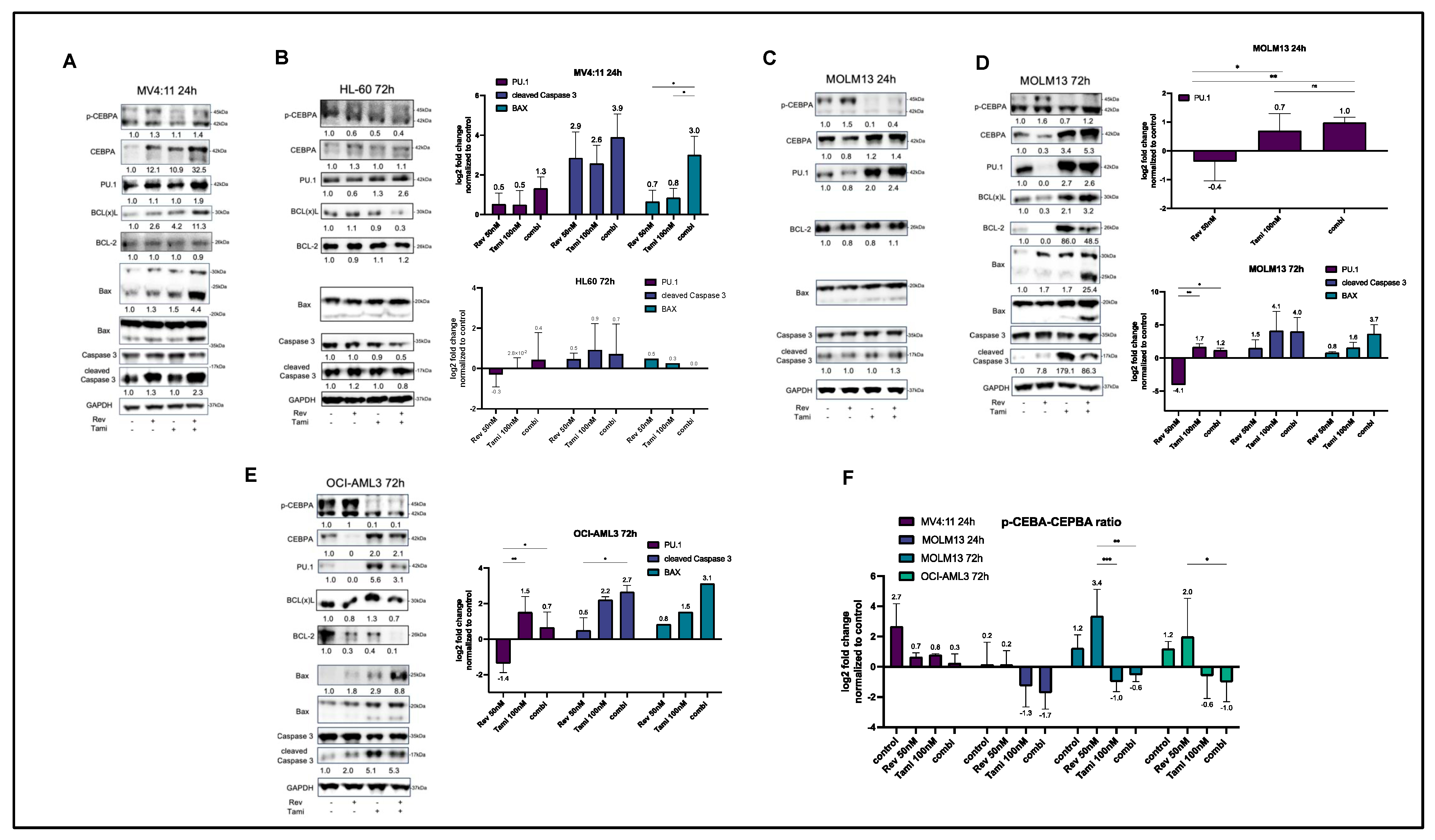
3.2. Induction of Differentiation and Apoptosis upon Combination Treatment Is Mediated by Restoration of CEBPA and PU.1 Expression and Activation of Caspase-3 and BAX
3.3. Combination of Revumenib and Tamibarotene Demonstrates a Synergistic Reduction in Metabolic Activity
3.4. Transcriptional Effects of Revumenib and Tamibarotene Detected by Quantitative Real-Time PCR
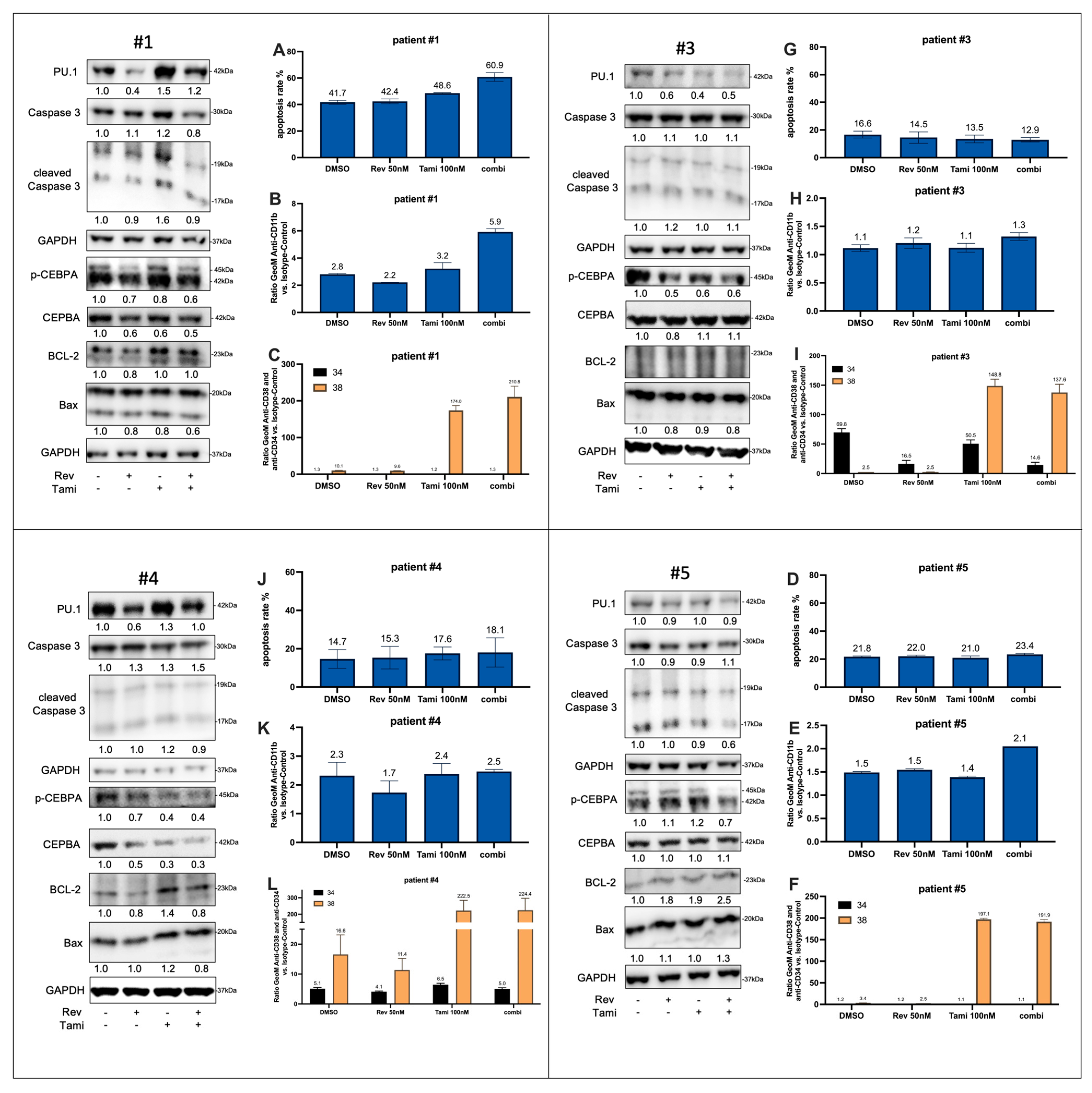
3.5. Effects of Tamibarotene and Revumenib on Differentiation and Apoptosis in Primary AML Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuhn, M.W.; Song, E.; Feng, Z.; Sinha, A.; Chen, C.W.; Deshpande, A.J.; Cusan, M.; Farnoud, N.; Mupo, A.; Grove, C.; et al. Targeting Chromatin Regulators Inhibits Leukemogenic Gene Expression in NPM1 Mutant Leukemia. Cancer Discov. 2016, 6, 1166–1181. [Google Scholar] [CrossRef] [PubMed]
- Mill, C.P.; Fiskus, W.; Das, K.; Davis, J.A.; Birdwell, C.E.; Kadia, T.M.; DiNardo, C.D.; Daver, N.; Takahashi, K.; Sasaki, K.; et al. Causal linkage of presence of mutant NPM1 to efficacy of novel therapeutic agents against AML cells with mutant NPM1. Leukemia 2023, 37, 1336–1348. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Somervaille, T.C.; Smith, K.S.; Rozenblatt-Rosen, O.; Meyerson, M.; Cleary, M.L. The menin tumor suppressor protein is an essential oncogenic cofactor for MLL-associated leukemogenesis. Cell 2005, 123, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Uckelmann, H.J.; Haarer, E.L.; Takeda, R.; Wong, E.M.; Hatton, C.; Marinaccio, C.; Perner, F.; Rajput, M.; Antonissen, N.J.C.; Wen, Y.; et al. Mutant NPM1 Directly Regulates Oncogenic Transcription in Acute Myeloid Leukemia. Cancer Discov. 2023, 13, 746–765. [Google Scholar] [CrossRef] [PubMed]
- Dohner, H.; Wei, A.H.; Appelbaum, F.R.; Craddock, C.; DiNardo, C.D.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Godley, L.A.; Hasserjian, R.P.; et al. Diagnosis and management of AML in adults: 2022 recommendations from an international expert panel on behalf of the ELN. Blood 2022, 140, 1345–1377. [Google Scholar] [CrossRef] [PubMed]
- Uckelmann, H.J.; Kim, S.M.; Wong, E.M.; Hatton, C.; Giovinazzo, H.; Gadrey, J.Y.; Krivtsov, A.V.; Rucker, F.G.; Dohner, K.; McGeehan, G.M.; et al. Therapeutic targeting of preleukemia cells in a mouse model of NPM1 mutant acute myeloid leukemia. Science 2020, 367, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Issa, G.C.; Aldoss, I.; DiPersio, J.; Cuglievan, B.; Stone, R.; Arellano, M.; Thirman, M.J.; Patel, M.R.; Dickens, D.S.; Shenoy, S.; et al. The menin inhibitor revumenib in KMT2A-rearranged or NPM1-mutant leukaemia. Nature 2023, 615, 920–924. [Google Scholar] [CrossRef] [PubMed]
- Perner, F.; Stein, E.M.; Wenge, D.V.; Singh, S.; Kim, J.; Apazidis, A.; Rahnamoun, H.; Anand, D.; Marinaccio, C.; Hatton, C.; et al. MEN1 mutations mediate clinical resistance to menin inhibition. Nature 2023, 615, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Soto-Feliciano, Y.M.; Sanchez-Rivera, F.J.; Perner, F.; Barrows, D.W.; Kastenhuber, E.R.; Ho, Y.J.; Carroll, T.; Xiong, Y.; Anand, D.; Soshnev, A.A.; et al. A Molecular Switch between Mammalian MLL Complexes Dictates Response to Menin-MLL Inhibition. Cancer Discov. 2023, 13, 146–169. [Google Scholar] [CrossRef] [PubMed]
- Fiskus, W.; Boettcher, S.; Daver, N.; Mill, C.P.; Sasaki, K.; Birdwell, C.E.; Davis, J.A.; Takahashi, K.; Kadia, T.M.; DiNardo, C.D.; et al. Effective Menin inhibitor-based combinations against AML with MLL rearrangement or NPM1 mutation (NPM1c). Blood Cancer J. 2022, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Fiskus, W.; Mill, C.P.; Birdwell, C.; Davis, J.A.; Das, K.; Boettcher, S.; Kadia, T.M.; DiNardo, C.D.; Takahashi, K.; Loghavi, S.; et al. Targeting of epigenetic co-dependencies enhances anti-AML efficacy of Menin inhibitor in AML with MLL1-r or mutant NPM1. Blood Cancer J. 2023, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- McKeown, M.R.; Corces, M.R.; Eaton, M.L.; Fiore, C.; Lee, E.; Lopez, J.T.; Chen, M.W.; Smith, D.; Chan, S.M.; Koenig, J.L.; et al. Superenhancer Analysis Defines Novel Epigenomic Subtypes of Non-APL AML, Including an RARalpha Dependency Targetable by SY-1425, a Potent and Selective RARalpha Agonist. Cancer Discov. 2017, 7, 1136–1153. [Google Scholar] [CrossRef]
- Muindi, J.R.; Young, C.W.; Warrell, R.P., Jr. Clinical pharmacology of all-trans retinoic acid. Leukemia 1994, 8, 1807–1812. [Google Scholar] [PubMed]
- Perez, M.W.; Sias-Garcia, O.; Daramola, A.; Wei, H.; Terrell, M.; Rashid, R.; Park, W.D.; Duong, K.; Horton, T.M.; Li, F.; et al. Defining the transcriptional control of pediatric AML highlights RARA as a superenhancer-regulated druggable dependency. Blood Adv. 2021, 5, 4864–4876. [Google Scholar] [CrossRef] [PubMed]
- Radomska, H.S.; Basseres, D.S.; Zheng, R.; Zhang, P.; Dayaram, T.; Yamamoto, Y.; Sternberg, D.W.; Lokker, N.; Giese, N.A.; Bohlander, S.K.; et al. Block of C/EBP alpha function by phosphorylation in acute myeloid leukemia with FLT3 activating mutations. J. Exp. Med. 2006, 203, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Imamura, T.; Yano, M.; Yoshida, H.; Fujiki, A.; Hirashima, Y.; Hosoi, H. Sensitivity of MLL-rearranged AML cells to all-trans retinoic acid is associated with the level of H3K4me2 in the RARalpha promoter region. Blood Cancer J. 2014, 4, e205. [Google Scholar] [CrossRef] [PubMed]
- McKeown, M.R.; Johannessen, L.; Lee, E.; Fiore, C.; di Tomaso, E. Antitumor synergy with SY-1425, a selective RARalpha agonist, and hypomethylating agents in retinoic acid receptor pathway activated models of acute myeloid leukemia. Haematologica 2019, 104, e138–e142. [Google Scholar] [CrossRef] [PubMed]
- de Botton, S.; Cluzeau, T.; Vigil, C.; Cook, R.J.; Rousselot, P.; Rizzieri, D.A.; Liesveld, J.L.; Fenaux, P.; Braun, T.; Banos, A.; et al. Targeting RARA overexpression with tamibarotene, a potent and selective RARalpha agonist, is a novel approach in AML. Blood Adv. 2023, 7, 1858–1870. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Wang, W.; Aldahdooh, J.; Malyutina, A.; Shadbahr, T.; Tanoli, Z.; Pessia, A.; Tang, J. SynergyFinder Plus: Toward Better Interpretation and Annotation of Drug Combination Screening Datasets. Genom. Proteom. Bioinform. 2022, 20, 587–596. [Google Scholar] [CrossRef]
- Yadav, B.; Wennerberg, K.; Aittokallio, T.; Tang, J. Searching for Drug Synergy in Complex Dose-Response Landscapes Using an Interaction Potency Model. Comput. Struct. Biotechnol. J. 2015, 13, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.M.; Mandrekar, S.J.; Sanford, B.L.; Laumann, K.; Geyer, S.; Bloomfield, C.D.; Thiede, C.; Prior, T.W.; Dohner, K.; Marcucci, G.; et al. Midostaurin plus Chemotherapy for Acute Myeloid Leukemia with a FLT3 Mutation. N. Engl. J. Med. 2017, 377, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Perl, A.E.; Martinelli, G.; Cortes, J.E.; Neubauer, A.; Berman, E.; Paolini, S.; Montesinos, P.; Baer, M.R.; Larson, R.A.; Ustun, C.; et al. Gilteritinib or Chemotherapy for Relapsed or Refractory FLT3-Mutated AML. N. Engl. J. Med. 2019, 381, 1728–1740. [Google Scholar] [CrossRef] [PubMed]
- DiNardo, C.D.; Jonas, B.A.; Pullarkat, V.; Thirman, M.J.; Garcia, J.S.; Wei, A.H.; Konopleva, M.; Dohner, H.; Letai, A.; Fenaux, P.; et al. Azacitidine and Venetoclax in Previously Untreated Acute Myeloid Leukemia. N. Engl. J. Med. 2020, 383, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, P.; Recher, C.; Vives, S.; Zarzycka, E.; Wang, J.; Bertani, G.; Heuser, M.; Calado, R.T.; Schuh, A.C.; Yeh, S.P.; et al. Ivosidenib and Azacitidine in IDH1-Mutated Acute Myeloid Leukemia. N. Engl. J. Med. 2022, 386, 1519–1531. [Google Scholar] [CrossRef] [PubMed]
- Matthews, A.H.; Pratz, K.W.; Carroll, M.P. Targeting Menin and CD47 to Address Unmet Needs in Acute Myeloid Leukemia. Cancers 2022, 14, 5906. [Google Scholar] [CrossRef] [PubMed]
- Issa, G.C.; Ravandi, F.; DiNardo, C.D.; Jabbour, E.; Kantarjian, H.M.; Andreeff, M. Therapeutic implications of menin inhibition in acute leukemias. Leukemia 2021, 35, 2482–2495. [Google Scholar] [CrossRef] [PubMed]
- Quentmeier, H.; Reinhardt, J.; Zaborski, M.; Drexler, H.G. FLT3 mutations in acute myeloid leukemia cell lines. Leukemia 2003, 17, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Dohner, K.; Thiede, C.; Jahn, N.; Panina, E.; Gambietz, A.; Larson, R.A.; Prior, T.W.; Marcucci, G.; Jones, D.; Krauter, J.; et al. Impact of NPM1/FLT3-ITD genotypes defined by the 2017 European LeukemiaNet in patients with acute myeloid leukemia. Blood 2020, 135, 371–380. [Google Scholar] [CrossRef]
- Roussel, M.; Moreau, P.; Hebraud, B.; Laribi, K.; Jaccard, A.; Dib, M.; Slama, B.; Dorvaux, V.; Royer, B.; Frenzel, L.; et al. Bortezomib, thalidomide, and dexamethasone with or without daratumumab for transplantation-eligible patients with newly diagnosed multiple myeloma (CASSIOPEIA): Health-related quality of life outcomes of a randomised, open-label, phase 3 trial. Lancet Haematol. 2020, 7, e874–e883. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Yan, Y.; Royal, M. A Phase 1, Open-Label, Dose-Escalation Study of the Safety and Efficacy of Anti-CD38 Antibody Drug Conjugate (STI-6129) in Patients with Relapsed or Refractory Multiple Myeloma. Blood 2021, 138, 4763. [Google Scholar] [CrossRef]
- Yu, S.; Li, Y.; Zhao, H.; Wang, Q.; Chen, P. The Histone Demethylase JMJD1C Regulates CAMKK2-AMPK Signaling to Participate in Cardiac Hypertrophy. Front. Physiol. 2020, 11, 539. [Google Scholar] [CrossRef] [PubMed]
- Salesse, S.; Verfaillie, C.M. BCR/ABL-mediated increased expression of multiple known and novel genes that may contribute to the pathogenesis of chronic myelogenous leukemia. Mol. Cancer Ther. 2003, 2, 173–182. [Google Scholar] [PubMed]
- Rogers, H.A.; Sousa, S.; Salto, C.; Arenas, E.; Coyle, B.; Grundy, R.G. WNT/beta-catenin pathway activation in Myc immortalised cerebellar progenitor cells inhibits neuronal differentiation and generates tumours resembling medulloblastoma. Br. J. Cancer 2012, 107, 1144–1152. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Dai, Y.; Liu, Z.; Hu, W.; Wan, Y. miR-342-3p Suppresses glioblastoma development via targeting CDK6. Acta Biochim. Pol. 2022, 69, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Todorov, V.T.; Desch, M.; Schubert, T.; Kurtz, A. The Pal3 promoter sequence is critical for the regulation of human renin gene transcription by peroxisome proliferator-activated receptor-gamma. Endocrinology 2008, 149, 4647–4657. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Molecular Genetics | Cytogenetics | ELN 2017 Risk Group |
|---|---|---|---|
| #1 | KMT2A/MLL, DNMT3A, SF3B1 | 46,XY, t(11;19)(q23;p13.3) | high |
| #2 | KMT2A/MLLT4 and KMT2A-PTD | 46,XX, t(6;11)(q27;q23) | high |
| #3 | n.a. | 46,XX, t(4;11)(X;X) | high |
| #4 | n.a. | 46,XY, t(11;19)(q23;p13.1) | high |
| #5 | NPM1, BCOR, GATA2, NRAS | 46,XY | intermediate |
| #6 | negative | 46,XY, t(9;11)(p21.3;q23) | high |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fleischmann, M.; Bechwar, J.; Voigtländer, D.; Fischer, M.; Schnetzke, U.; Hochhaus, A.; Scholl, S. Synergistic Effects of the RARalpha Agonist Tamibarotene and the Menin Inhibitor Revumenib in Acute Myeloid Leukemia Cells with KMT2A Rearrangement or NPM1 Mutation. Cancers 2024, 16, 1311. https://doi.org/10.3390/cancers16071311
Fleischmann M, Bechwar J, Voigtländer D, Fischer M, Schnetzke U, Hochhaus A, Scholl S. Synergistic Effects of the RARalpha Agonist Tamibarotene and the Menin Inhibitor Revumenib in Acute Myeloid Leukemia Cells with KMT2A Rearrangement or NPM1 Mutation. Cancers. 2024; 16(7):1311. https://doi.org/10.3390/cancers16071311
Chicago/Turabian StyleFleischmann, Maximilian, Julia Bechwar, Diana Voigtländer, Mike Fischer, Ulf Schnetzke, Andreas Hochhaus, and Sebastian Scholl. 2024. "Synergistic Effects of the RARalpha Agonist Tamibarotene and the Menin Inhibitor Revumenib in Acute Myeloid Leukemia Cells with KMT2A Rearrangement or NPM1 Mutation" Cancers 16, no. 7: 1311. https://doi.org/10.3390/cancers16071311
APA StyleFleischmann, M., Bechwar, J., Voigtländer, D., Fischer, M., Schnetzke, U., Hochhaus, A., & Scholl, S. (2024). Synergistic Effects of the RARalpha Agonist Tamibarotene and the Menin Inhibitor Revumenib in Acute Myeloid Leukemia Cells with KMT2A Rearrangement or NPM1 Mutation. Cancers, 16(7), 1311. https://doi.org/10.3390/cancers16071311