
Roles of TRPM8 Ion Channels in Cancer: Proliferation, Survival, and Invasion

Abstract
:1. Introduction
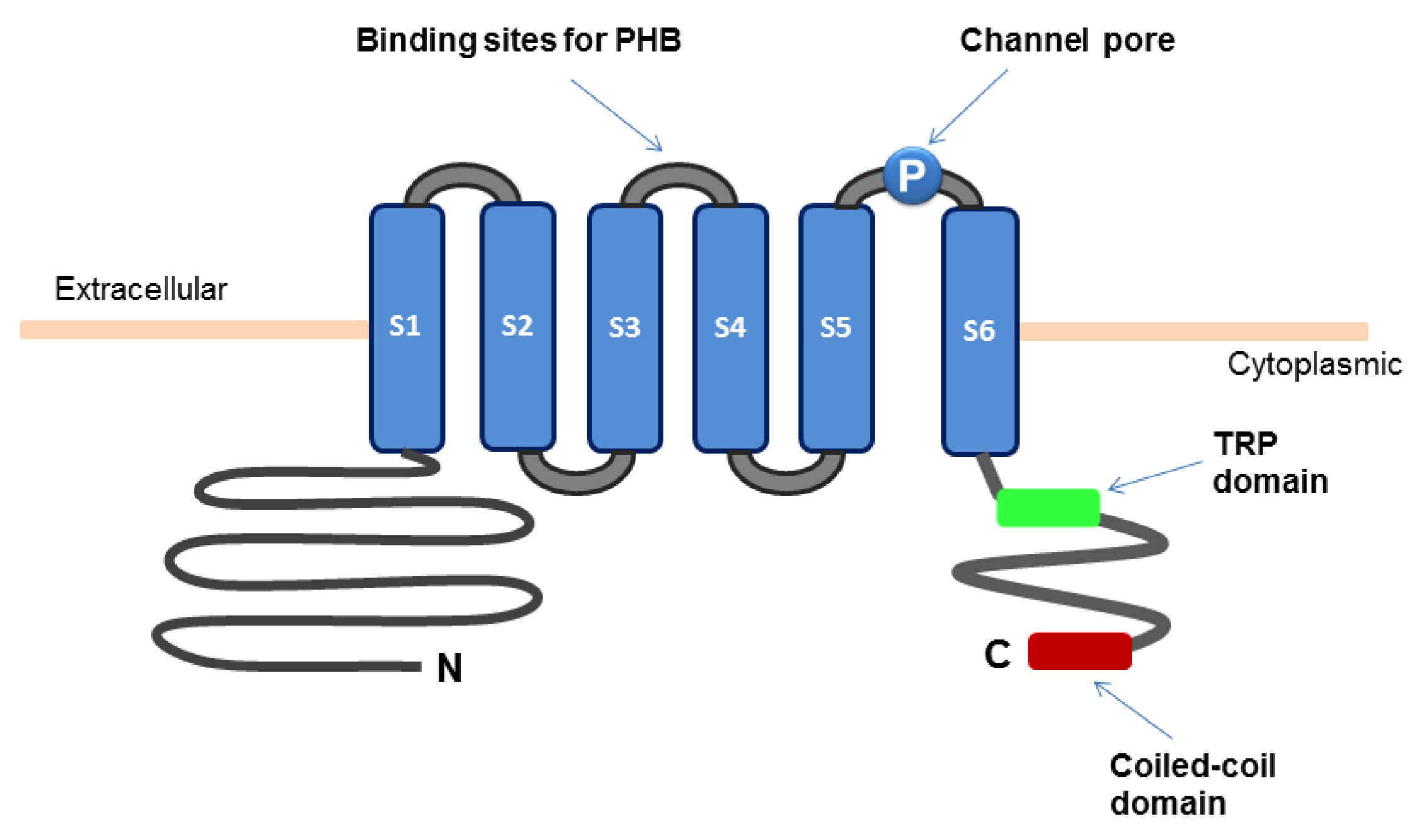
2. Structure and Functions of TRPM8 Ion Channels

3. TRPM8 Channels in Cancers
3.1. Expression of TRPM8 Ion Channels in Cancers

{kind=link}
{kind=link}
| Cancer | Expression | Functional Role | References |
|---|---|---|---|
| Prostatic carcinoma | Up-regulated in tissues and androgen receptor-expressing cell lines (LNCaP, VcaP, C4-2B, NCI-H660). | Cell proliferation, survival, migration, hypoxic growth, xenograft growth, angiogenesis | [31,32,35,36,37,38,39,40,41,42,43,44,45,46] |
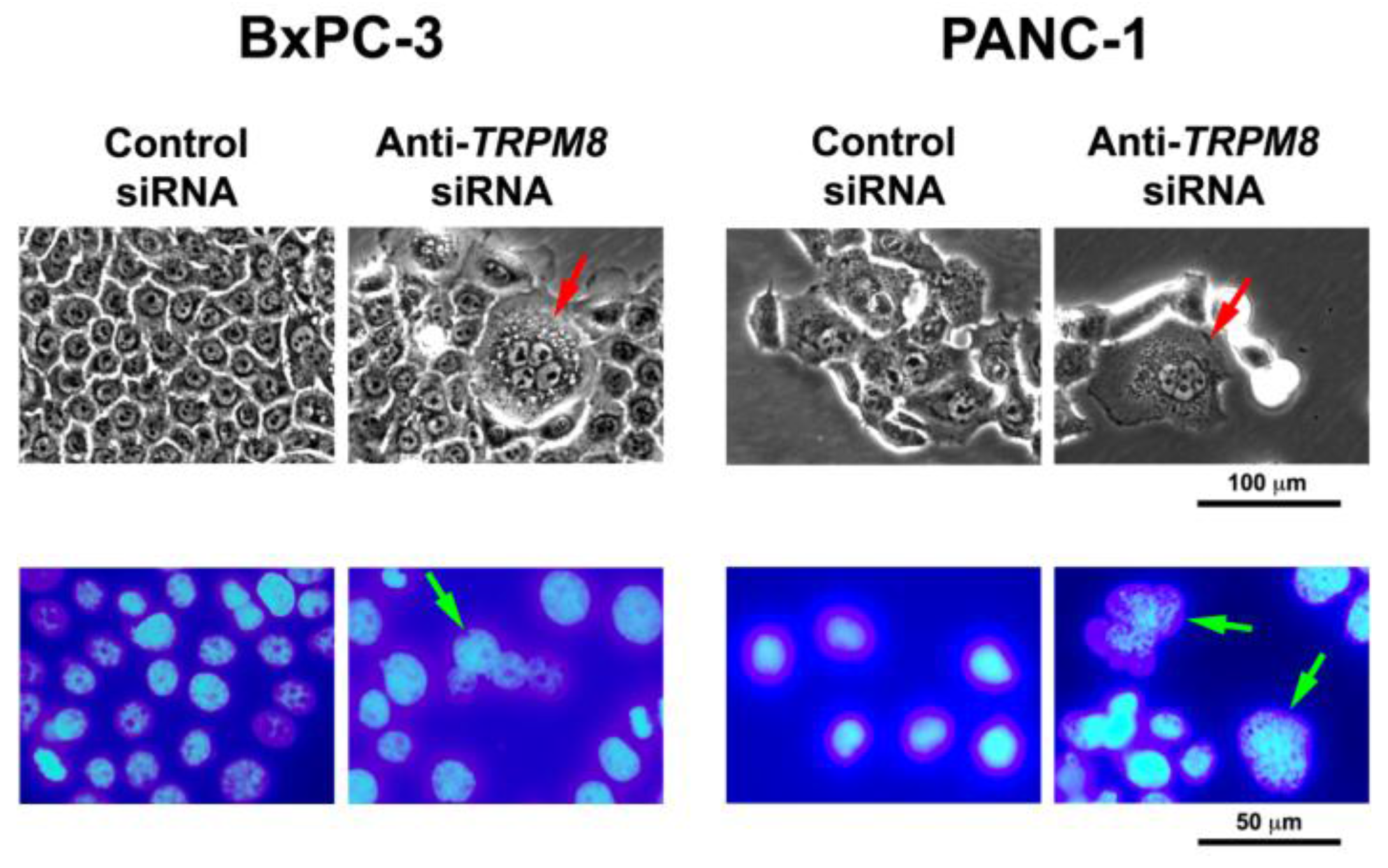
| Pancreatic carcinoma | Up-regulated in cell lines (PL45, MIA PaCa-2, PANC-1, HPAF-II, BxPC-3, Capan-1, Panc 02.03). Over-expressed in pancreatic adenocarcinoma. Also aberrantly expressed in chronic pancreatitis, pancreatic intraepithelial neoplasm, intraductal papillary mucinous neoplasm, solid pseudopapillary neoplasm, adenosquamous carcinoma, and neuroendocrine tumor. | Cell proliferation, cell cycle progression, replicative senescence, survival, migration, invasion. | [47,48,49,50,51] |
| Breast adenocarcinoma | Over-expressed in cell line (MCF-7, T47D, MDA-MB231, BT549, SKBR3, ZR-75-30). Over-expressed in breast adenocarcinoma tissues. | Cell migration, invasion | [40,52,53,54] |
| Lung carcinoma | Expressed in tissues and cell lines (LLC-1, LLC-2, LLC-3). | Cell proliferation, adhesion, migration, invasion, resistance to hypothermia. | [40,55] |
| Colorectal adenocarcinoma | Expressed in tissues and cell lines (Caco-2, HCT 116). | Cell growth, survival, xenograft tumor growth, chemically-induced cancer growth. | [40,56] |
| Melanoma | Expressed in tissues and cell lines (G-361, A-375, Mel 202, Mel 270, 92.1, omm 2.3). | Cell survival | [40,57,58,59] |
| Urinary bladder carcinoma | Expressed in cell line (T24). Over-expressed in urothelial carcinoma tissues. | Cell survival | [60,61] |
| Neuroblastoma | Up-regulated expression in cell line (IMR-32) in response to 5-bromo-2-deoxyuridine induced differentiation. | Not reported | [62] |
| Glioblastoma multiforme | Expressed in cell line (DBTRG) and tissues. | Cell migration, survival | [63,64] |
| Neuroendocrine tumor | Expressed in neuroendocrine tumor cell line (BON) and tissues. | Secretion of neurotensin. | [50,65] |
| Oral squamous cell carcinoma | Expressed in cell lines derived from tongue (HSC3 and HSC4). | Cell migration and invasion. | [66] |
| Osteosarcoma | Expression in osteosarcoma cell lines (U2OS, MG-63, SaOS2, HOS); increased expression in osteosarcoma as compared to osteochondroma. | Cell proliferation, cell cycle progression, survival, migration, and invasion. | [67] |
3.2. Roles of TRPM8 Ion Channels in Cancers
3.2.1. Role of TRPM8 in Cancer Cells Proliferation

3.2.2. Role of TRPM8 in Survival of Cancer Cells
3.2.3. Role of TRPM8 in Cancer Cells Migration and Invasion
3.2.4. Mechanisms of TRPM8-Mediated Biological Processes in Cancer
4. Conclusions and Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- Fonfria, E.; Murdock, P.R.; Cusdin, F.S.; Benham, C.D.; Kelsell, R.E.; McNulty, S. Tissue distribution profiles of the human TRPM cation channel family. J. Recept. Signal Transduc. Res. 2006, 26, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Almaraz, L.; Manenschijn, J.-A.; de la Pena, E.; Viana, F. TRPM8. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Nilius, B., Flockerzi, V., Eds.; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 547–579. [Google Scholar]
- Bautista, D.M.; Siemens, J.; Glazer, J.M.; Tsuruda, P.R.; Basbaum, A.I.; Stucky, C.L.; Jordt, S.E.; Julius, D. The menthol receptor TRPM8 is the principal detector of environment cold. Nature 2007, 448, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, A.; Murray, A.N.; Mathur, J.; Earley, T.J.; Petrus, M.J.; Patapoutian, A. TRPM8 is required for cold sensation in mice. Neuron 2007, 54, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Colburn, R.W.; Lubin, M.L.; Stone, D.J., Jr.; Wang, Y.; Lawrence, D.; D’Andrea, M.R.; Brandt, M.R.; Liu, Y.; Flores, C.M.; Qin, N.; et al. Attenuated cold sensitivity in TRPM8 null mice. Neuron 2007, 54, 379–386. [Google Scholar] [CrossRef] [PubMed]
- McCoy, D.D.; Zhou, L.; Nguyen, A.-K.; Watts, A.G.; Donovan, C.M.; McKemy, D.D. Enhanced insulin clearance in mice lacking TRPM8 channels. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E78–E88. [Google Scholar] [CrossRef] [PubMed]
- Voets, T.; Owsianik, G.; Nilius, B. TRPM8. Handb. Exp. Pharmacol. 2007, 179, 329–344. [Google Scholar] [PubMed]
- Dragoni, I.; Guida, E.; McIntyre, P. The cold and menthol receptor TRPM8 contains a functionally important double cysteine motif. J. Biol. Chem. 2006, 281, 37353–37360. [Google Scholar] [CrossRef] [PubMed]
- Phelps, C.B.; Gaudet, R. The role of the N terminus and transmembrane domain of TRPM8 in channel localization and tetramerization. J. Biol. Chem. 2007, 282, 36474–36480. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.P.; Egressy, K.; Lim, A.; Edwardson, J.M. AFM imaging reveals the tetrameric structure of the TRPM8 channel. Biochem. Biophys. Res. Commun. 2010, 394, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Janssens, A.; Voets, T. Ligand stoichiometry of the cold- and menthol-activated activated channel TRPM8. J. Physiol. 2011, 589, 4827–4835. [Google Scholar] [CrossRef] [PubMed]
- Tsuruda, P.R.; Julius, D.; Minor, D.L., Jr. Coiled coils direct assembly of a cold-activated TRP channel. Neuron 2006, 51, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Erler, I.; Al-Ansary, D.M.M.; Wissenbach, U.; Wagner, T.F.J.; Flockerzi, V.; Niemeyer, B.A. Trafficking and assembly of the cold-sensitive TRPM8 channel. J. Biol. Chem. 2006, 281, 38396–38404. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Qin, F. Functional control of cold- and menthol-sensitive TRPM8 ion channels by phosphatidylinositol 4,5-bisphosphate. J. Neurosci. 2005, 25, 1674–1681. [Google Scholar] [CrossRef] [PubMed]
- Rohács, T.; Lopes, C.M.B.; Michailidis, I.; Logothetis, D.E. PI(4,5)P2 regulates the activation and desensitization of TRPM8 channels through the TRP domain. Nat. Neurosci. 2005, 8, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Bandell, M.; Dubin, A.E.; Petrus, M.J.; Orth, A.; Mathur, J.; Hwang, S.W.; Patapoutian, A. High-throughput random mutagenesis screen reveals TRPM8 residues specifically required for activation by menthol. Nat. Neurosci. 2006, 9, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Bavencoffe, A.; Gkika, D.; Kondratskyi, A.; Beck, B.; Borowiec, A.-S.; Bidaux, G.; Busserolles, J.; Eschalier, A.; Shuba, Y.; Skryma, R.; et al. The transient receptor potential channel TRPM8 is inhibited via the α2A adrenoreceptor signaling pathway. J. Biol. Chem. 2010, 285, 9410–9419. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Yudin, Y.; Bikard, Y.; Chen, W.; Liu, T.; Li, H.; Jendrossek, D.; Cohen, A.; Pavlov, E.; Rohács, T.; et al. Polyester modification of the mammalian TRPM8 channel protein: Implications for structure and function. Cell Rep. 2013, 4, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Gkika, D.; Prevarskaya, N. Molecular mechanisms of TRP regulation in tumor growth and metastasis. Biochim. Biophys. Acta 2009, 1793, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Bidaux, G.; Beck, B.; Zholos, A.; Gordienko, D.; Lemonnier, L.; Flourakis, M.; Roudbaraki, M.; Borowiec, A.S.; Fernandez, J.; Delcourt, P.; et al. Regulation of activity of transient receptor potential melastatin 8 (TRPM8) channel by its short isoforms. J. Biol. Chem. 2012, 287, 2948–2962. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.A.; Skryma, R.; Bidaux, G.; Magleby, K.L.; Scholfield, C.N.; McGeown, J.G.; Prevarskaya, N.; Zholos, A.V. Short iosforms of the cold receptor TRPM8 inhibit channel gating by mimicking heat action rather than chemical inhibitors. J. Biol. Chem. 2012, 287, 2963–2970. [Google Scholar] [CrossRef] [PubMed]
- Voets, T.; Droogmans, G.; Wissenbach, U.; Janssens, A.; Flockerzi, V.; Nilius, B. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature 2004, 430, 748–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malkia, A.; Madrid, R.; Meseguer, V.; de la Pena, E.; Valero, M.; Belmonte, C.; Viana, F. Bidirectional shifts of TRPM8 channel gating by temperature and chemical agents modulate the cold sensitivity of mammalian thermoreceptors. J. Physiol. 2007, 581, 155–174. [Google Scholar] [CrossRef] [PubMed]
- Chuang, H.H.; Neuhausser, W.M.; Julius, D. The super-cooling agent icilin reveals a mechanism of coincidence detection by a temperature-sensitive TRP channel. Neuron 2004, 43, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.A.; Chase, H.W.N.; Bevan, S. TRPM8 activation by menthol, icilin, and cold is differentially modulated by intracellular pH. J. Neurosci. 2004, 24, 5364–5369. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.S.; Raisinghani, M.; Pingle, S.C.; Long, C.; Pimentel, F. Downregulation of transient receptor potential melastatin 8 by protein kinase C-mediated dephosphorylation. J. Neurosci. 2005, 25, 11322–11329. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Starowicz, K.; Moriello, A.S.; Vivese, M.; Orlando, P.; di Marzo, V. Regulation of transient receptor potential channels of melastatic type 8 (TRPM8): Effect of camp, cannabinoid CB1 receptors and endovanilloids. Exp. Cell Res. 2007, 313, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Gkika, D.; Flourakis, M.; Lemonnier, L.; Prevarskaya, N. PSA reduces prostate cancer cell motility by stimulating TRPM8 activity and plasma membrane expression. Oncogene 2010, 29, 4611–4616. [Google Scholar] [CrossRef] [PubMed]
- Gkika, D.; Lemonnier, L.; Shapovalov, G.; Gordienko, D.; Poux, C.; Bernardini, M.; Bokhobza, A.; Bidaux, G.; Degerny, C.; Verreman, K.; et al. TRP channel-associated factors are a novel protein family that regulates TRPM8 trafficking and activity. J. Cell Biol. 2015, 208, 89–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asuthkar, S.; Elustondo, P.A.; Demirkhanyan, L.; Sun, X.; Baskaran, P.; Velpula, K.K.; Thyagarajan, B.; Pavlov, E.V.; Zakharian, E. The TRPM8 protein is a testosterone receptor. I. Biochemical evidence for direct TRPM8-testosterone interactions. J. Biol. Chem. 2015, 290, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Asuthkar, S.; Demirkhanyan, L.; Sun, X.; Elustondo, P.A.; Krishnan, V.; Baskaran, P.; Velpula, K.K.; Thyagarajan, B.; Pavlov, E.V.; Zakharian, E.; et al. The TRPM8 protein is a testosterone receptor. II. Functional evidence for an ionotropic effect of testosterone on TRPM8. J. Biol. Chem. 2015, 290, 2670–2688. [Google Scholar] [CrossRef] [PubMed]
- Asuthkar, S.; Velpula, K.K.; Elustondo, P.A.; Demirkhanyan, L.; Zakharian, E. TRPM8 channel as a novel molecular target in androgen-regulated prostate cancer cells. Oncotarget 2015, 6, 17221–17236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Barritt, G.J. Evidence that TRPM8 is an androgen-dependent Ca2+ channel required for the survival of prostate cancer cells. Cancer Res. 2004, 64, 8365–8373. [Google Scholar] [CrossRef] [PubMed]
- Bidaux, G.; Roudbaraki, M.; Merle, C.; Crepin, A.; Delcourt, P.; Slomianny, C.; Thebault, S.; Bonnal, J.-L.; Benahmed, M.; Cabon, F.; et al. Evidence for specific TRPM8 expression in human prostate secretory epithelial cells: Functional androgen receptor requirement. Endocr. Relat. Cancer 2005, 12, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Thebault, S.; Lemonnier, L.; Bidaux, G.; Flourakis, M.; Bavencoffe, A.; Gordienko, D.; Roudbaraki, M.; Delcourt, P.; Panchin, Y.; Shuba, Y.; et al. Novel role of cold/menthol-sensitive transient receptor potential melastatine family member 8 (TRPM8) in the activation of store-operated channels in LNCaP human prostate cancer epithelial cells. J. Biol. Chem. 2005, 280, 39423–39435. [Google Scholar] [CrossRef] [PubMed]
- Bidaux, G.; Flourakis, M.; Thebault, S.; Zholos, A.; Beck, B.; Gkika, D.; Roudbaraki, M.; Bonnal, J.-L.; Mauroy, B.; Shuba, Y.; et al. Prostate cell differentiation status determines transient receptor potential melastatin member 8 channel subcellular localization and function. J. Clin. Investig. 2007, 117, 1647–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsavaler, L.; Shapero, M.H.; Morkowski, S.; Laus, R. Trp-p8, a novel prostate-specific gene, is up-regulated in prostate cancer and other malignancies and shares high homology with transient receptor potential calcium channel proteins. Cancer Res. 2001, 61, 3760–3769. [Google Scholar] [PubMed]
- Fuessel, S.; Sickert, D.; Meye, A.; Klenk, U.; Schmidt, U.; Schmitz, M.; Rost, A.K.; Weigle, B.; Kiessling, A.; Wirth, M.P.; et al. Multiple tumor marker analyses (PSA, hK2, PSCA, trp-p8) in primary prostate cancers using quantitative RT-PCR. Int. J. Oncol. 2003, 23, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Xu, Z.; Zou, C.; Wu, D.; Wang, Y.; Yao, X.; Ng, C.-F.; Chan, F.L. Ion channel TRPM8 promotes hypoxic growth fo prostate cancer cells via an O2-independent and RACK1-mediated mechanism of HIF-1α stabilization. J. Pathol. 2014, 234, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Bai, V.U.; Murthy, S.; Chinnakannu, K.; Muhlethaler, F.; Tejwani, S.; Barrack, E.R.; Kim, S.-H.; Menon, M.; Reddy, G.P.V. Androgen regulated TRPM8 expression: A potential mRNA marker for metastatic prostate cancer detection in body fluids. Int. J. Oncol. 2010, 36, 443–450. [Google Scholar] [PubMed]
- Valero, M.L.I.; Mello de Queiroz, F.; Stuhmer, W.; Viana, F.; Pardo, L.A. TRPM8 ion channels differentially modulate proliferation and cell cycle distribution of normal and cancer prostate cells. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.-H.; Wang, X.-H.; Wang, H.-P.; Hu, H.-Q. Effects of TRPM8 on the proliferation and motility of prostate cancer PC-3 cells. Asian J. Androl. 2009, 11, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, X.; Yang, Z.; Cao, H.; Meng, Z.; Wang, Y.; Chen, D. Effects of TRPM8 on the proliferation and angiogenesis of prostate cancer PC-3 cells in vivo. Oncol. Lett. 2011, 2, 1213–1217. [Google Scholar] [PubMed]
- Yee, N.S.; Zhou, W.; Lee, M. Transient receptor potential channel TRPM8 is over-expressed and required for cellular proliferation in pancreatic adenocarcinoma. Cancer Lett. 2010, 297, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Yee, N.S.; Chan, A.S.; Yee, J.D.; Yee, R.K. TRPM7 and TRPM8 ion channels in pancreatic adenocarcinoma: Potential roles as cancer biomarkers and targets. Scientifica 2012, 2012, 415158. [Google Scholar] [CrossRef] [PubMed]
- Yee, N.S.; Brown, R.D.; Lee, M.S.; Zhou, W.; Jensen, C.; Gerke, H.; Yee, R.K. TRPM8 ion channel is aberrantly expressed and required for preventing replicative senescence in pancreatic adenocarcinoma: Potential role of TRPM8 as a biomarker and target. Cancer Biol. Ther. 2012, 13, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Yee, N.S.; Li, Q.; Kazi, A.A.; Yang, Z.; Berg, A.; Yee, R.K. Aberrantly over-expressed TRPM8 channels in pancreatic adenocarcinoma: Correlation with tumor size/stage and requirement for cancer cells invasion. Cells 2014, 3, 500–516. [Google Scholar] [CrossRef] [PubMed]
- Cucu, D.; Chirtoiu, G.; Petrescu, S.; Babes, A.; Stanica, L.; Duda, D.G.; Horii, A.; Dima, S.O.; Popescu, I. Characterization of functional transient receptor potential melastatin 8 channels in human pancreatic adenocarcinoma cells. Pancreas 2014, 43, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Dhennin-Duthille, I.; Gautier, M.; Faouzi, M.; Guilbert, A.; Brevet, M.; Vaudry, D.; Ahidouch, A.; Sevestre, H.; Ouadid-Ahidouch, H. High expression of transient receptor potential channels in human breast cancer epithelial cells and tissues: Correlation with pathological parameters. Cell. Physiol. Biochem. 2011, 28, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Chodon, D.; Guilbert, A.; Dhennin-Duthille, I.; Gautier, M.; Telliez, M.S.; Sevestre, H.; Ouadid-Ahidouch, H. Estrogen regulation of TRPM8 expression in breast cancer cells. BMC Cancer 2012, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, Y.; Shuai, S.; Ding, D.; Li, R.; Luo, R. TRPM8 promotes aggressiveness of breast cancer cells by regulating EMT via activating AKT/GSK-3β pathway. Tumor Biol. 2014, 35, 8969–8977. [Google Scholar] [CrossRef] [PubMed]
- Du, G.-J.; Li, J.-H.; Liu, W.-J.; Liu, Y.-H.; Zhao, B.; Li, H.-R.; Hou, X.-D.; Li, H.; Qi, X.-X.; Duan, Y.-J.; et al. The combination of TRPM8 and TRPA1 expression causes an invasive phenotype in lung cancer. Tumor Biol. 2014, 35, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, F.; Pagano, E.; Romano, B.; Panzera, S.; Maiello, F.; Coppola, D.; de Petrocellis, L.; Buono, L.; Orlando, P.; Izzo, A.A.; et al. Colon carcinogenesis is inhibited by the TRPM8 antagonist cannabigerol, a Cannabis-derived non-psychotropic cannabinoid. Carcinogenesis 2014, 35, 2787–2797. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, H.; Ugawa, S.; Ueda, T.; Morita, A.; Shimada, S. TRPM8 activation suppresses cellular viability in human melanoma. Am. J. Physiol. Cell Physiol. 2008, 295, C296–C301. [Google Scholar] [CrossRef] [PubMed]
- Kijpornyongpan, T.; Sereemaspun, A.; Chanchao, C. Dose-dependent cytotoxic effects of menthol on human malignant melanoma A-375 cells: Correlation with TRPM8 transcript expression. Asian Pac. J. Cancer Prev. 2014, 15, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Derckx, R.; Reinach, P.S.; Garreis, F.; Bohm, A.; Schmelzer, L.; Skosyrski, S.; Ramesh, N.; Abdelmessih, S.; Polat, O.K.; et al. Calcium regulation by temperature-sensitive transient receptor potential channels in human uveal melanoma cells. Cell. Signal. 2014, 26, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Jiang, L.M.; Ge, B.; Zhang, T.Y.; Zhao, X.K.; Zhou, X. Over-expression of TRPM8 is associated with poor prognosis in urothelial carcinoma of bladder. Tumor Biol. 2014, 35, 11499–11504. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, X.; Yang, Z.; Wang, B.; Li, S. Menthol induces cell death via the TRPM8 channel in the human bladder cancer cell line T24. Oncology 2009, 77, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Louhivuori, L.M.; Bart, G.; Larsson, K.P.; Louhivuori, V.; Nasman, J.; Nordstrom, T.; Koivisto, A.P.; Akerman, K.E. Differentiation dependent expression of TRPA1 and TRPM8 channels in IMR-32 human neuroblastoma cells. J. Cell. Physiol. 2009, 221, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Wondergem, R.; Bartley, J.W. Menthol increases human glioblastoma intracellular Ca2+, BK channel activity and cell migration. J. Biomed. Sci. 2009, 16, 90. [Google Scholar] [CrossRef] [PubMed]
- Alptekin, M.; Eroglu, S.; Tutar, E.; Sencan, S.; Geyik, M.A.; Ulasli, M.; Demiryurek, A.T.; Camci, C. Gene expressions of TRP channels in glioblastoma multiforme and relation with survival. Tumor Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Strowski, M.Z.; Kaiser, S.; Plath, T.; Giesecke, Y.; Neumann, M.; Hosokawa, H.; Kobayashi, S.; Langrehr, J.; Neuhaus, P.; et al. Transient receptor potential channel TRPM8 agonists stimulate calcium influx and neurotensin secretion in neuroendocrine tumor cells. Neuroendocrinology 2007, 85, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, Y.; Ohkubo, T.; Ikebe, T.; Yamazaki, J. Blockade of TRPM8 activity reduces the invasion potential of oral squamous carcinoma cell lines. Int. J. Oncol. 2012, 40, 1431–1440. [Google Scholar] [PubMed]
- Wang, Y.; Yang, Z.; Meng, Z.; Cao, H.; Zhu, G.; Liu, T.; Wang, X. Knockdown of TRPM8 suppresses cancer malignancy and enhances epirubicin-induced apoptosis in human osteosarcoma cells. Int. J. Biol. Sci. 2014, 10, 90–102. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yee, N.S. Roles of TRPM8 Ion Channels in Cancer: Proliferation, Survival, and Invasion. Cancers 2015, 7, 2134-2146. https://doi.org/10.3390/cancers7040882
Yee NS. Roles of TRPM8 Ion Channels in Cancer: Proliferation, Survival, and Invasion. Cancers. 2015; 7(4):2134-2146. https://doi.org/10.3390/cancers7040882
Chicago/Turabian StyleYee, Nelson S. 2015. "Roles of TRPM8 Ion Channels in Cancer: Proliferation, Survival, and Invasion" Cancers 7, no. 4: 2134-2146. https://doi.org/10.3390/cancers7040882
APA StyleYee, N. S. (2015). Roles of TRPM8 Ion Channels in Cancer: Proliferation, Survival, and Invasion. Cancers, 7(4), 2134-2146. https://doi.org/10.3390/cancers7040882