
Differential miRNA-Expression as an Adjunctive Diagnostic Tool in Neuroendocrine Tumors of the Lung

Abstract
:1. Introduction
2. Results
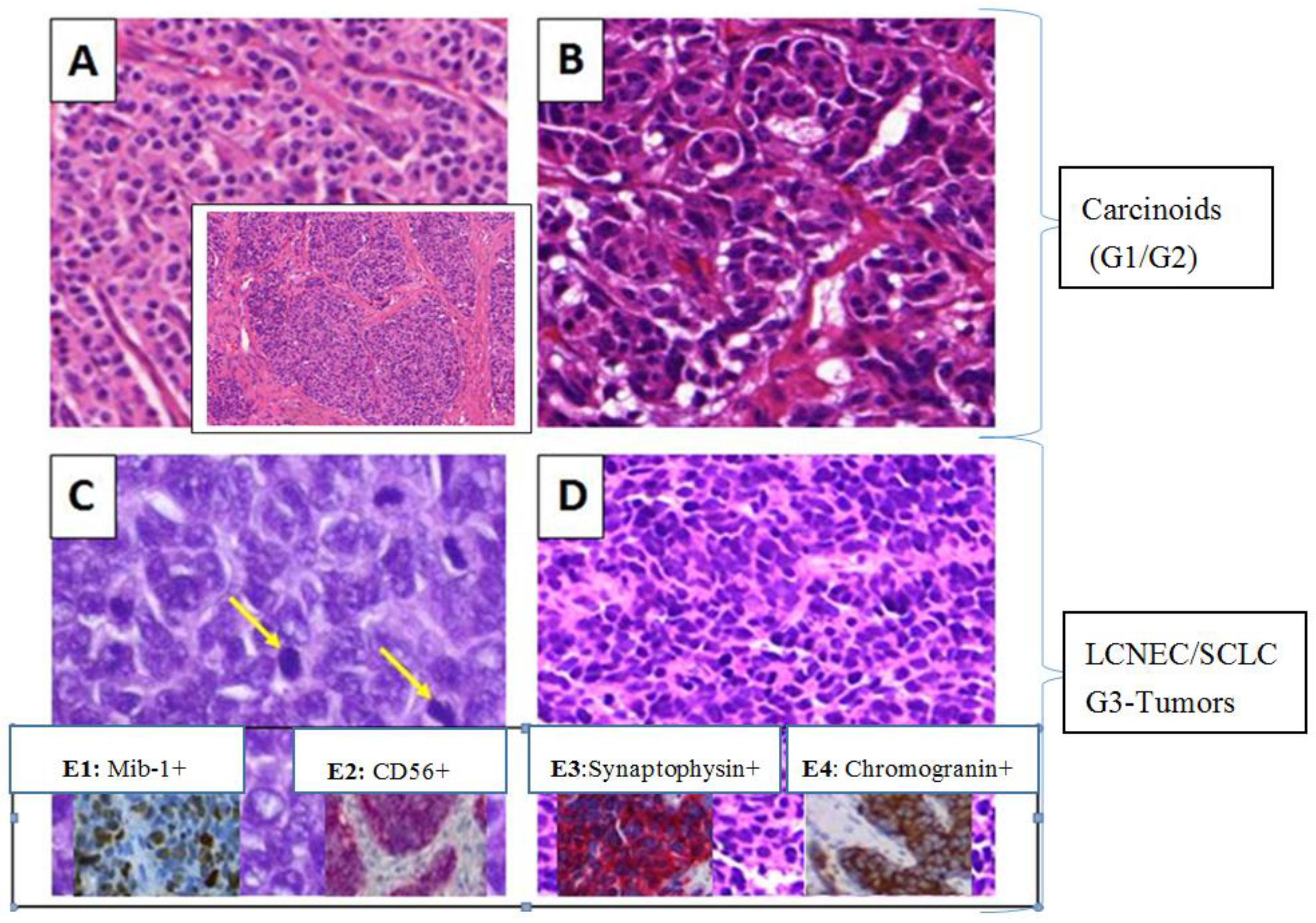
2.1. Baseline Characteristics and Pathologic Assessment
- 20 carcinoids (G1/G2), 52.6%
- 18 LCNECs/SCLCs (G3), 47.4%
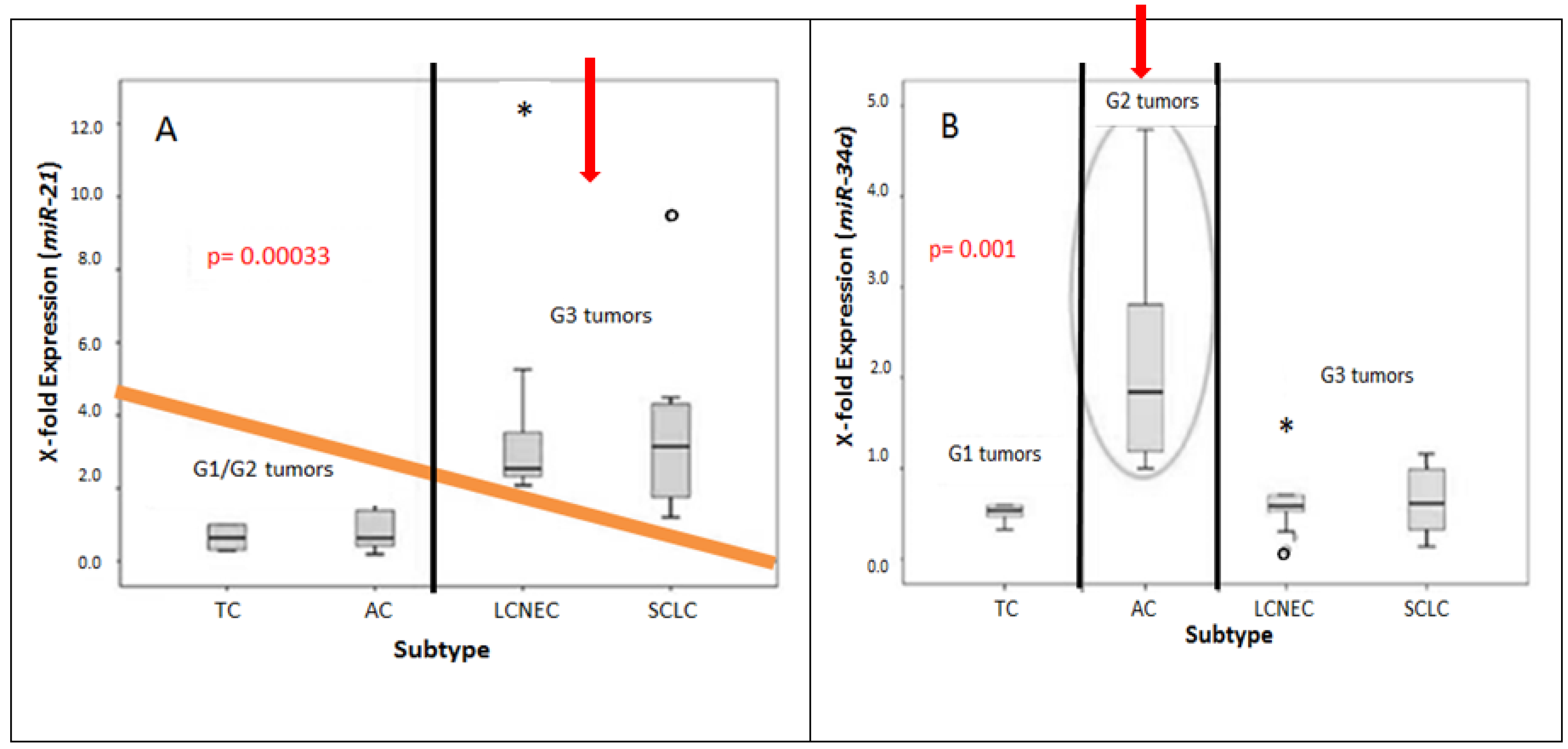
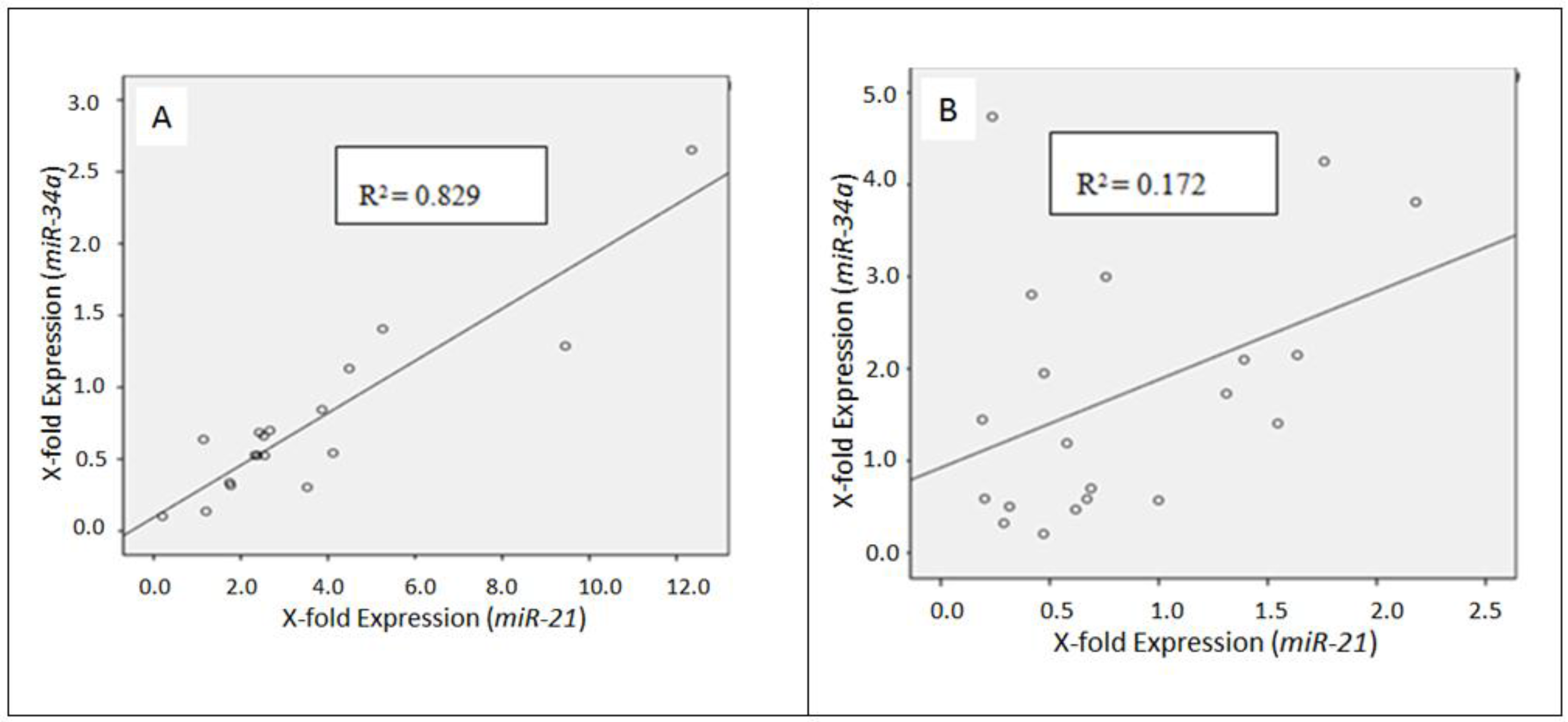
2.2. Differential miRNA Expression in Pulmonary Subtypes of Neuroendocrine Tumors
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. RNA Isolation
4.3. Primer Sequences
4.4. Quantitative Analysis of miRNAs in Neuroendocrine Tumors of the Lung
4.5. Statistics and Mathematics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Faggiano, A.; Ferella, P.; Grimaldi, F.; Campana, D.; Manzoni, M.; Davi, M.V.; Bianchi, A.; Valcavi, R.; Papini, E.; Giuffrida, D. Natural history of gastro-entero-pancreatic and thoracic neuroendocrine tumors. Data from a large prospective and retrospective Italian Epidemiological study: The NET management study. J. Endocrinol. Investig. 2012, 35, 817–823. [Google Scholar]
- Bertino, E.M. Pulmonary neuroendocrine/carcinoid tumors. Cancer 2009, 115, 4434–4441. [Google Scholar] [CrossRef] [PubMed]
- Fisseler-Eckhoff, A.; Demes, M. Neuroendocrine Tumors of the Lung. Cancers 2012, 4, 777–798. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Junhua, Z.; Zuyao, Y.; Yafang, H.; Xinyin, W.; Hong, S.; Jinling, T.; Qing, C. KRAS, BRAF and PIK3CA mutations and the loss of PTEN expression in Chinese patients with colorectal cancer. PLoS ONE 2012, 7, e36653. [Google Scholar]
- Bonikos, D.S.; Bensch, K.G.; Jamplis, R.W. Peripheral pulmonary carcinoid tumors. Cancer 1997, 37, 1977–1998. [Google Scholar] [CrossRef]
- Suarez, Y.; Sessa, W.C. MicroRNAs as novel regulators of angiogenesis. Circ. Res. 2009, 104, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancers 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M. RNAi, microRNAs, and human disease. Cancer Chemother. Pharmacol. 2006, 58, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Hashimi, S.T.; Fulcher, J.A.; Chang, M.H.; Gov, L.; Wang, S.; Lee, B. MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the coordinate regulation of dendritic cell differentiation. Blood 2009, 114, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Hwang, S.J.; Bae, Y.C.; Jung, J.S. MiR-21 regulates adipogenic differentiation through the modulation of TGF-beta signaling in mesenchymal stem cells derived from human adipose tissue. Stem Cells 2009, 27, 3093–3102. [Google Scholar] [PubMed]
- Meng, F.; Henson, R.; Wehbe-Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Papagiannakopoulos, T.; Shapiro, A.; Kosik, K.S. MicroRNA-21 targets a network of key tumor-suppressive pathways in glioblastoma cells. Cancer Res. 2008, 68, 8164–8172. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, N.S.; Manavalan, T.T.; Dougherty, S.M.; Riggs, K.A.; Li, Y.; Klinge, C.M. Estradiol downregulates miR-21 expression and increases miR-21 target gene expression in MCF-7 breast cancer cells. Nucleic Acids Res. 2009, 37, 2584–2595. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Ha, S.Y.; Roh, M.S. Altered expression of PTEN and its major regulator microRNA-21 in pulmonary neuroendocrine tumors. Korean J. Pathol. 2014, 48, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J. miR-34a repression of SIRT1 regulates apoptosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13421–13426. [Google Scholar] [CrossRef] [PubMed]
- DeVita, V.T.; Hellman, S.; Rosenberg, S.A. Cancer, Principles & Practice of Oncology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2006. [Google Scholar]
- Raghavan, D. Textbook of Uncommon Cancer; Wiley: Chichester, UK, 2006. [Google Scholar]
- Travis, W.D.; Linnoila, R.I.; Tsokos, M.G.; Hitchcock, C.L.; Cutler, G.B.; Nieman, L.; Chrousos, G.; Pass, H.; Doppman, J. Neuroendocrine tumors of the lung with proposed criteria for large-cell neuroendocrine carcinoma. An ultrastructural, immunohistochemical, and flow cytometric study of 35 cases. Am. J. Surg. 1991, 15, 529–553. [Google Scholar]
- Mezzetti, M.; Raveglia, F.; Panigalli, T.; Giuliani, L.; Lo Giudice, F.; Meda, S.; Conforti, S. Assessment of outcomes in typical and atypical carcinoids according to latest WHO classification. Ann. Thorac. Surg. 2003, 76, 1838–1842. [Google Scholar] [CrossRef]
- Rekhtman, N. Neuroendocrine tumors of the lung: An update. Arch. Pathol. Lab. Med. 2009, 134, 1628–1638. [Google Scholar]
- Travis, W.D.; Travis, L.B.; Devesa, S.S. Lung cancer. Cancer 1995, 75, 191–202. [Google Scholar] [CrossRef]
- Chen, L.C.; Travis, W.D.; Krug, L.M. Pulmonary neuroendocrine tumors: What (little) do we know? J. Natl. Compr. Canc. Netw. 2004, 4, 623–630. [Google Scholar]
- Demes, M.; Bartsch, H.; Scheil-Bertram, S.; Mücke, R.; Fisseler-Eckhoff, A. Real-Time PCR Data Processing Shown by the Analysis of Colorectal Specific Candidate Genes, ERCC1, RRM1 and TS in Relation to β2M as Endogenous Control. Appl. Sci. 2012, 2, 139–159. [Google Scholar] [CrossRef]
- Perez-Novo, C.A.; Claeys, C.; Speleman, F.; van Cauwenberge, P.; Bachert, C.; Vandesompele, J. Impact of RNA quality on reference gene expression stability. Biotechniques 2005, 39, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Fleige, S.; Pfaffl, M.W. RNA integrity and the effect on the real-time qRT-PCR performance. Mol. Asp. Med. 2006, 27, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A. A-Z of Quantitative PCR, 1st ed.; International University Line: San Diego, CA, USA, 2004. [Google Scholar]
- Wang, T.; Brown, M.J. mRNA quantification by real time TaqMan polymerase chain reaction: Validation and comparison with RNase protection. Anal. Biochem. 1999, 269, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, J.L.; Seah, C.; Brown, S.; Tang, P.; Jamieson, F.; Drews, S.J. Use of Bordetella pertussis BP3385 to establish a cutoff value for an IS481-targeted real-time PCR assay. J. Clin. Microbiol. 2008, 46, 3798–3799. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Yin, Z.; Li, X.; Wu, W.; Zhou, B. Meta-analysis of human lung cancer microRNA expression profiling studies comparing cancer tissues with normal tissues. J. Exp. Clin. Cancer Res. 2012, 31, 54. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Lin, C.Y.; Yuan, H.Y.; Xiong, B. Overexpression of miR-21 promotes proliferation and reduces apoptosis in non-small cell lung cancer. Zhonghua Zhong Liu Za Zhi (Chin. J. Oncol.) 2009, 33, 742–746. [Google Scholar]
- Wang, Z.X.; Bian, H.B.; Wang, J.R.; Cheng, Z.X.; Wang, K.M.; De, W. Prognostic significance of serum miRNA-21 expression in human non-small cell lung cancer. J. Surg. Oncol. 2004, 104, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Mairinger, F.D.; Ting, S.; Werner, R.; Walter, R.F.H.; Hager, T.; Vollbrecht, C.; Christoph, D.; Worm, K.; Mairinger, T.; Grabellus, S.Y.S.; et al. Different micro-RNA expression profiles distinguish subtypes of neuroendocrine tumors of the lung: Results of a profiling study. Mod. Pathol. 2014, 27, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Lee, E.H.; Ha, S.Y.; Lee, C.H.; Chang, H.K.; Chang, S.; Kwon, K.Y.; Hwang, I.S.; Roh, M.S.; Seo, J.W. Altered expression of microRNA miR-21, miR-155, and let-7a and their roles in pulmonary neuroendocrine tumors. Pathol. Int. 2012, 62, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, D.; Sha, J.; Sun, P.; Huang, Y. MicroRNA-21 directly targets MARCKS and promotes apoptosis resistance and invasion in prostate cancer cells. Biochem. Biophys. Res. Commun. 2009, 383, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Oudai, H.; Aamir, A.; Seema, S.; Fazlul, H.S. Recent updates on the role of microRNAs in prostate cancer. J. Hematol. 2012, 5, 5–9. [Google Scholar]
- Asangani, I.A.; Rasheed, S.A.; Nikolova, D.A.; Leupold, J.H.; Colburn, N.H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008, 27, 2128–2136. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Visone, R.; di Leva, G.; Donati, V.; Petrocca, F.; Casalini, P.; Taccioli, C.; Volinia, S.; Liu, C.G.; Alder, H.; et al. MicroRNA signatures in human ovarian cancer. Cancer Res. 2007, 67, 8699–8707. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Voortman, J.; Dingemans, A.M.; Voeller, D.M.; Pham, T.; Wang, Y.; Giaccone, G. MicroRNA expression and clinical outcome of small cell lung cancer. PLoS ONE 2011, 6, e21300. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.; Goldstraw, P.; Nicholson, A.G.; Travis, W.D.; Jett, J.R.; Ferolla, P.; Bomanji, J.; Rusch, V.W.; Asamura, H.; Skogseid, B.; et al. Proceedings of the IASLC international workshop on advances in pulmonary neuroendocrine tumors. J. Thorac. Oncol. 2007, 3, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Li, L.; Yu, T.; Zheng, L.L.; Liu, W.; Zuo, X.Y.; He, Y.M.; Zhang, S.Y.; Zhu, W.; Cao, J.P.; et al. miR-34a regulated apoptosis in liver cells by targeting the KLF4 gene. Cell. Mol. Biol. 2014, 19, 52–64. [Google Scholar]
- Applied Biosystems. microRNA TaqMan Assay Protocol. Available online: https://genome.med.harvard.edu/documents/qpcr/microRNATaqManAssayProtocol.pdf (accessed on 16 July 2015).




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Absolute Number | Percentage (%) | |
|---|---|---|---|
| Cases | 38 | 100 | |
| CANCER SUBTYPE 1 | AC | 6 | 15.8 |
| TC | 14 | 36.8 | |
| LCNEC | 10 | 26.3 | |
| SCLC | 8 | 21.1 | |
| LOCATION 2 | ML | 5 | 13.2 |
| LU | 13 | 34.2 | |
| RU | 4 | 10.5 | |
| LL | 8 | 21.1 | |
| RL | 8 | 21.1 | |
| HISTOLOGICAL GRADING (G) | G1 | 14 | 36.8 |
| G2 | 6 | 15.8 | |
| G3 | 18 | 47.4 | |
| TUMOR (T) STAGE | T1a | 15 | 39.5 |
| T1b | 7 | 18.4 | |
| T2a | 7 | 18.4 | |
| T2b | 1 | 2.6 | |
| pT3 | 7 | 18.4 | |
| n.a. 3 | 1 | 2.6 | |
| NODAL (N) STAGE | N0 | 27 | 71.1 |
| N1 | 5 | 13.2 | |
| N2 | 3 | 7.9 | |
| N3 | 2 | 5.3 | |
| n.a. | 1 | 2.6 |
| miRNA | Mature miRNA Sequence (5′–3′) | Length | Species | Melting Temperature |
|---|---|---|---|---|
| 21 | UAG CUU AUC AGA CUG AUG UUG A | 22 bp | Homo sapiens | 49.2 °C |
| 34a | UGG CAG UGU CUU AGC UGG UUG U | 22 bp | Homo sapiens | 53.0 °C |
| U47 (endogenous control) | TAA TGA TTC TGC CAA ATG AAA TAT AAT GAT ATC ACT GTA AAA CCG TTC CAT TTT GAT TCT GAG GT | 65 bp | Homo sapiens | 67.2 °C |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demes, M.; Aszyk, C.; Bartsch, H.; Schirren, J.; Fisseler-Eckhoff, A. Differential miRNA-Expression as an Adjunctive Diagnostic Tool in Neuroendocrine Tumors of the Lung. Cancers 2016, 8, 38. https://doi.org/10.3390/cancers8040038
Demes M, Aszyk C, Bartsch H, Schirren J, Fisseler-Eckhoff A. Differential miRNA-Expression as an Adjunctive Diagnostic Tool in Neuroendocrine Tumors of the Lung. Cancers. 2016; 8(4):38. https://doi.org/10.3390/cancers8040038
Chicago/Turabian StyleDemes, Melanie, Christoph Aszyk, Holger Bartsch, Joachim Schirren, and Annette Fisseler-Eckhoff. 2016. "Differential miRNA-Expression as an Adjunctive Diagnostic Tool in Neuroendocrine Tumors of the Lung" Cancers 8, no. 4: 38. https://doi.org/10.3390/cancers8040038
APA StyleDemes, M., Aszyk, C., Bartsch, H., Schirren, J., & Fisseler-Eckhoff, A. (2016). Differential miRNA-Expression as an Adjunctive Diagnostic Tool in Neuroendocrine Tumors of the Lung. Cancers, 8(4), 38. https://doi.org/10.3390/cancers8040038