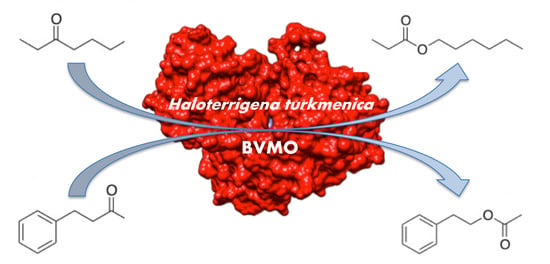
Unique Features of a New Baeyer–Villiger Monooxygenase from a Halophilic Archaeon

 ,
,  ,
,
Abstract
:
1. Introduction
2. Results
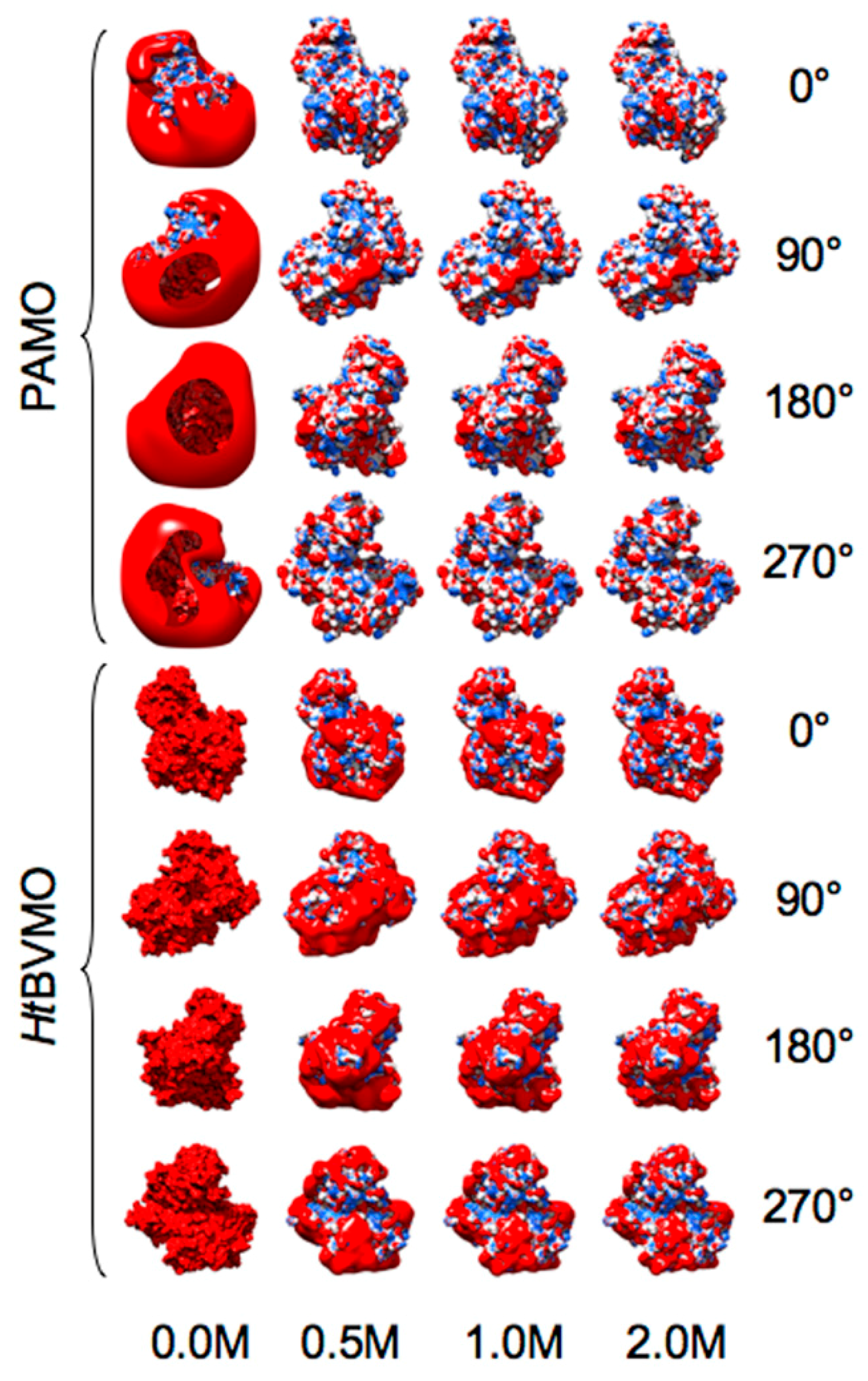
2.1. Identification, Structural Modeling and Electrostatic Analysis of HtBVMO
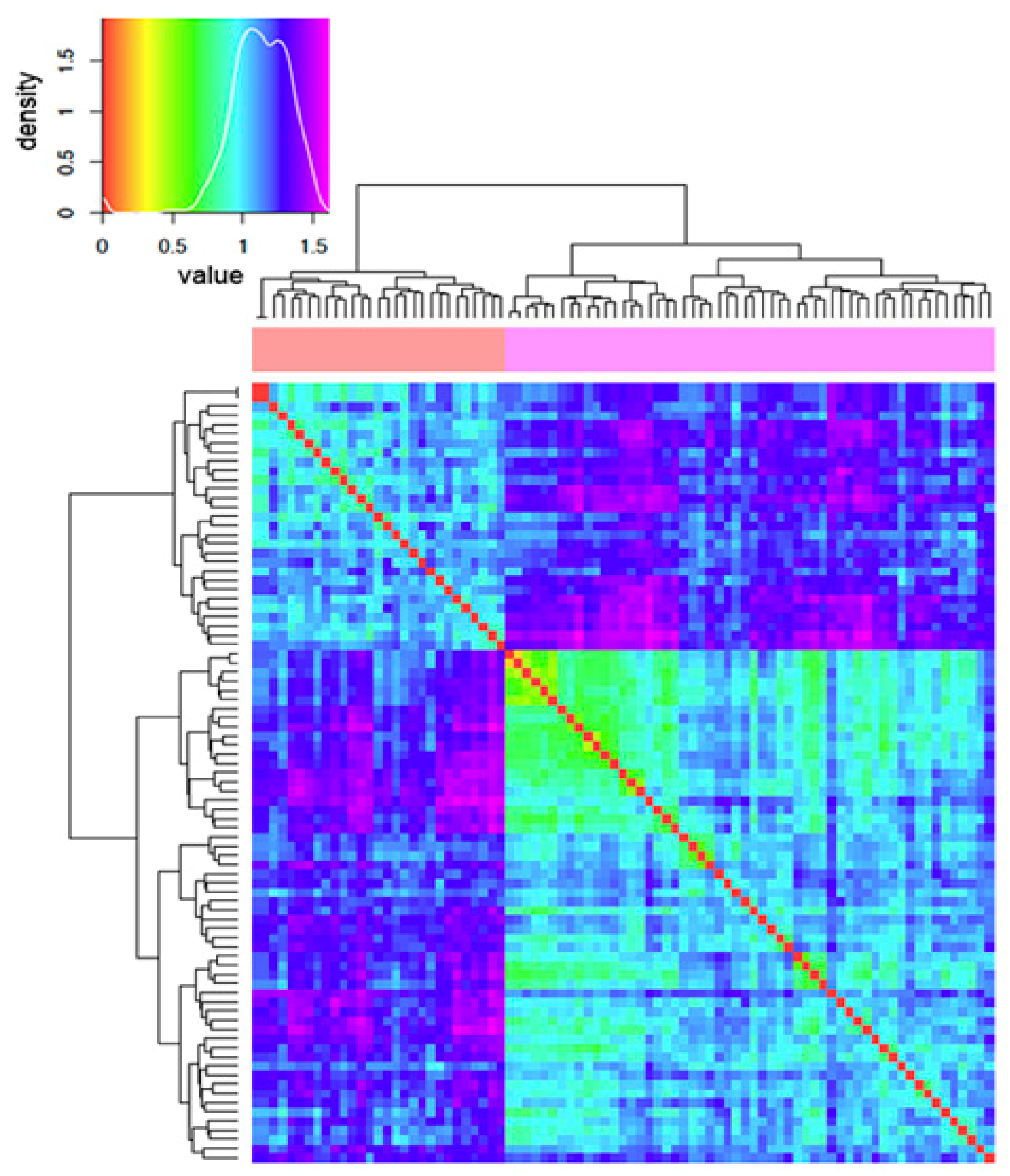
2.2. Comparative Electrostatic and Normal Modes Analysis of HtBVMO
2.3. Normal Modes Analysis of HtBVMO and Soluble/Insoluble Enzymes
2.4. Cloning, Expression, and Purification of HtBVMO
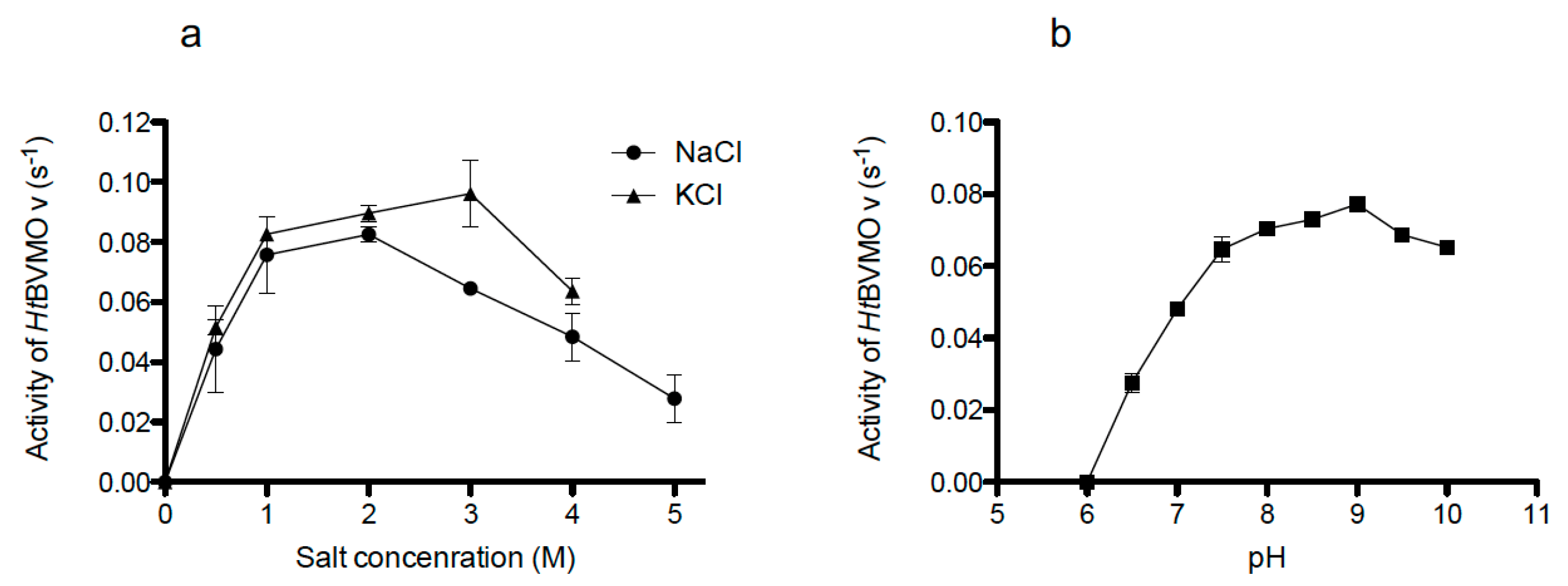
2.5. Salt and pH-Dependent Activity of the Recombinant Enzyme
2.6. Salt-Dependent Folding of HtBVMO
2.7. Steady-State Kinetics
2.8. Enzymatic Conversions
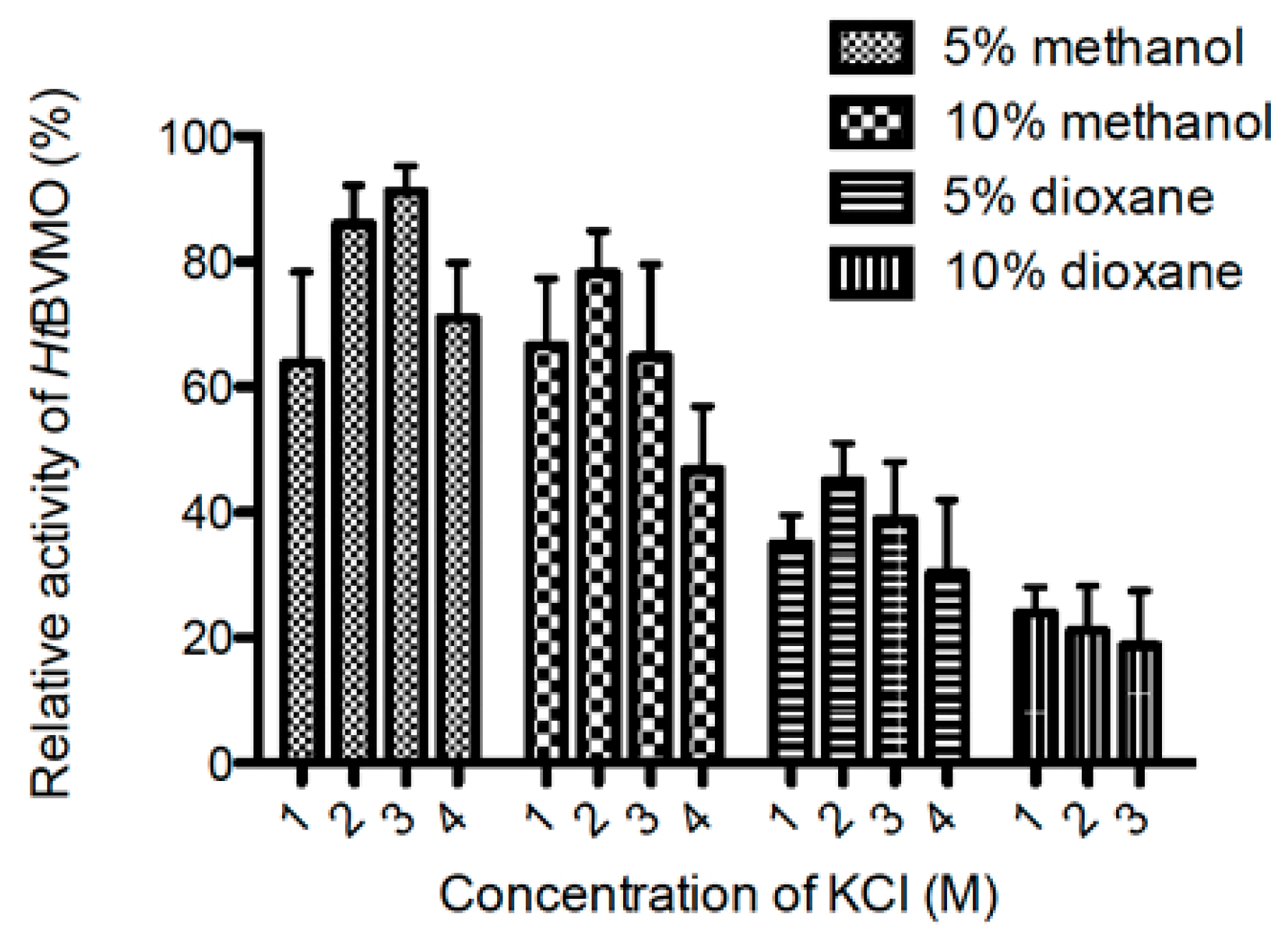
2.9. Activity of HtBVMO in Aqueous–Organic Media
3. Discussion
4. Materials and Methods
4.1. Bioinformatic Identification, Structural Modeling and Electrostatics Analysis
4.2. Single and Comparative Normal Mode Analysis
4.3. Cloning, Expression, and Purification
4.4. In-Gel Digestion and Extraction of Peptides for ESI MS
4.5. Enzymatic Activity Measurements and Kinetic Characterization
4.6. Circular Dichroism
4.7. ThermoFluor
4.8. Conversions and GC-MS Analysis
4.9. Biotransformations and GC Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krow, G.R. The Baeyer–Villiger oxidation of ketones and aldehydes. Org. React. 2004, 43, 251–798. [Google Scholar]
- Ten Brink, G.-J.; Arends, I.; Sheldon, R.A. The Baeyer−Villiger reaction: New developments toward greener procedures. Chem. Rev. 2004, 104, 4105–4124. [Google Scholar] [CrossRef]
- Kamerbeek, N.M.; Olsthoorn, A.J.; Fraaije, M.W.; Janssen, D.B. Substrate specificity and enantioselectivity of 4-hydroxyacetophenone monooxygenase. Appl. Environ. Microbiol. 2003, 69, 419–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento, F.; Peralta, R.; Blamey, J.M. Cold and Hot Extremozymes: Industrial Relevance and Current Trends. Front. Bioeng. Biotechnol. 2015, 3, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malito, E.; Alfieri, A.; Fraaije, M.W.; Mattevi, A. Crystal structure of a Baeyer-Villiger monooxygenase. Proc. Natl. Acad. Sci. USA 2004, 101, 13157–13162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, E.; Castellanos, J.; Mattevi, A.; Fraaije, M.W. Characterization and Crystal Structure of a Robust Cyclohexanone Monooxygenase. Angew. Chemi. Int. Ed. 2016, 55, 15852–15855. [Google Scholar] [CrossRef] [PubMed]
- Fürst, M.J.L.J.; Savino, S.; Dudek, H.M.; Gómez Castellanos, J.R.; Gutiérrez de Souza, C.; Rovida, S.; Fraaije, M.W.; Mattevi, A. Polycyclic Ketone Monooxygenase from the Thermophilic Fungus Thermothelomyces thermophila: A Structurally Distinct Biocatalyst for Bulky Substrates. J. Am. Chem. Soc. 2017, 139, 627–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, E.; Tindall, B.J.; Fähnrich, R.; Lapidus, A.; Copeland, A.; Del Rio, T.G.; Lucas, S.; Chen, F.; Tice, H.; Cheng, J.F.; et al. Complete genome sequence of Haloterrigena turkmenica type strain (4k T). Stand. Genom. Sci. 2010, 2, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Burg, B. Extremophiles as a source for novel enzymes. Curr. Opin. Microbiol. 2003, 6, 213–218. [Google Scholar] [CrossRef]
- Elleuche, S.; Schröder, C.; Sahm, K.; Antranikian, G. Extremozymes—Biocatalysts with unique properties from extremophilic microorganisms. Curr. Opin. Biotechnol. 2014, 29, 116–123. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Siroosi, M.; Atashgahi, S.; Smidt, H.; Ventosa, A. Systematics of haloarchaea and biotechnological potential of their hydrolytic enzymes. Microbiology 2017, 163, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Singh, A.K. Haloarchaea: Worth exploring for their biotechnological potential. Biotechnol. Lett. 2017, 39, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Sellek, G.A.; Chaudhuri, J.B. Biocatalysis in organic media using enzymes from extremophiles. Enzym. Microb. Technol. 1999, 25, 471–482. [Google Scholar] [CrossRef]
- Manikandan, K.; Bhardwaj, A.; Gupta, N.; Lokanath, N.K.; Ghosh, A.; Reddy, V.S.; Ramakumar, S. Crystal structures of native and xylosaccharide-bound alkali thermostable xylanase from an alkalophilic Bacillus sp. NG-27: Structural insights into alkalophilicity and implications for adaptation to polyextreme conditions. Protein Sci. 2006, 15, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Fraaije, M.W.; Kamerbeek, N.M.; van Berkel, W.J.H.; Janssen, D.B. Identification of a Baeyer–Villiger monooxygenase sequence motif. FEBS Lett. 2002, 518, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Vallon, O. New sequence motifs in flavoproteins: Evidence for common ancestry and tools to predict structure. Proteins 2000, 38, 95–114. [Google Scholar] [CrossRef]
- Riebel, A.; Dudek, H.M.; de Gonzalo, G.; Stepniak, P.; Rychlewski, L.; Fraaije, M.W. Expanding the set of rhodococcal Baeyer–Villiger monooxygenases by high-throughput cloning, expression and substrate screening. Appl. Microbiol. Biotechnol. 2012, 95, 1479–1489. [Google Scholar] [CrossRef] [Green Version]
- Beier, A.; Bordewick, S.; Genz, M.; Schmidt, S.; van den Bergh, T.; Peters, C.; Joosten, H.J.; Bornscheuer, U.T. Switch in Cofactor Specificity of a Baeyer-Villiger Monooxygenase. Chembiochem 2016, 17, 2312–2315. [Google Scholar] [CrossRef]
- Bordoli, L.; Kiefer, F.; Arnold, K.; Benkert, P.; Battey, J.; Schwede, T. Protein structure homology modeling using SWISS-MODEL workspace. Nat. Protoc. 2009, 4, 1–13. [Google Scholar] [CrossRef]
- Wang, Q.; Canutescu, A.A.; Dunbrack, R.L., Jr. SCWRL and MolIDE: Computer programs for side-chain conformation prediction and homology modeling. Nat. Protoc. 2008, 3, 1832–1847. [Google Scholar] [CrossRef] [Green Version]
- Benkert, P.; Kunzli, M.; Schwede, T. QMEAN server for protein model quality estimation. Nucleic Acids Res. 2009, 37, W510–W514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, K.L.; Baker, P.J.; Fisher, M.; Ruzheinikov, S.; Gilmour, D.J.; Bonete, M.J.; Ferrer, J.; Pire, C.; Esclapez, J.; Rice, D.W. Analysis of protein solvent interactions in glucose dehydrogenase from the extreme halophile Haloferax mediterranei. Proc. Natl. Acad. Sci. USA 2006, 103, 4846–4851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dym, O.; Mevarech, M.; Sussman, J.L. Structural features that stabilize halophilic malate dehydrogenase from an archaebacterium. Science 1995, 267, 1344–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frolow, F.; Harel, M.; Sussman, J.L.; Mevarech, M.; Shoham, M. Insights into protein adaptation to a saturated salt environment from the crystal structure of a halophilic 2Fe-2S ferredoxin. Nat. Struct. Mol. Biol. 1996, 3, 452–458. [Google Scholar] [CrossRef]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.T.; Dutta, C. Molecular signature of hypersaline adaptation: Insights from genome and proteome composition of halophilic prokaryotes. Genome Biol. 2008, 9, R70. [Google Scholar] [CrossRef] [Green Version]
- Tadeo, X.; López-Méndez, B.; Trigueros, T.; Laín, A.; Castaño, D.; Millet, O. Structural basis for the aminoacid composition of proteins from halophilic archaea. PLoS Biol. 2009, 7, e1000257. [Google Scholar] [CrossRef] [Green Version]
- Chakravorty, D.; Khan, M.F.; Patra, S. Multifactorial level of extremostability of proteins: Can they be exploited for protein engineering? Extremophiles 2017, 21, 419–444. [Google Scholar] [CrossRef]
- Ceccoli, R.D.; Bianchi, D.A.; Fink, M.J.; Mihovilovic, M.D.; Rial, D.V. Cloning and characterization of the Type I Baeyer-Villiger Monooxygenase from Leptospira biflexa. AMB Express 2017, 7, 87. [Google Scholar] [CrossRef] [Green Version]
- Righetto, I.; Milani, A.; Cattoli, G.; Filippini, F. Comparative structural analysis of haemagglutinin proteins from type A influenza viruses: Conserved and variable features. BMC Bioinform. 2014, 15, 363. [Google Scholar] [CrossRef] [Green Version]
- Heidari, A.; Righetto, I.; Filippini, F. Electrostatic Variation of Haemagglutinin as a Hallmark of the Evolution of Avian Influenza Viruses. Sci. Rep. 2018, 8, 1929. [Google Scholar] [CrossRef] [Green Version]
- Meeuwsen, S.M.; Hodac, A.N.; Adams, L.M.; McMunn, R.D.; Anschutz, M.S.; Carothers, K.J.; Egdorf, R.E.; Hanneman, P.M.; Kitzrow, J.P.; Keonigsberg, C.K.; et al. Investigation of intrinsic dynamics of enzymes involved in metabolic pathways using coarse-grained normal mode analysis. Cogent Biol. 2017, 3, 1. [Google Scholar] [CrossRef]
- Tiwari, S.P.; Fuglebakk, E.; Hollup, S.M.; Skjærven, L.; Cragnolini, T.; Grindhaug, S.H.; Tekle, K.M.; Reuter, N. WEBnm@ v2.0: Web server and services for comparing protein flexibility. BMC Bioinform. 2014, 15, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, D.D.; Ciulla, R.A.; Roberts, M.F. Osmoadaptation in archaea. Appl. Environ. Microbiol. 1999, 65, 1815–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madern, D.; Zaccai, G. Molecular adaptation: The malate dehydrogenase from the extreme halophilic bacterium Salinibacter ruber behaves like a non-halophilic protein. Biochimie 2004, 86, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Eddy, M.L.; Jablonski, P.E. Purification and characterization of a membrane-associated ATPase from Natronococcus occultus, a haloalkaliphilic archaeon. FEMS Microbiol. Lett. 2000, 189, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Müller-Santos, M.; de Souza, E.M.; Pedrosa, F.O.; Mitchell, D.A.; Longhi, S.; Carrière, F.; Canaan, S.; Krieger, N. First evidence for the salt-dependent folding and activity of an esterase from the halophilic archaeon Haloarcula marismortui. Biochim. Biophys. Acta 2009, 1791, 719–729. [Google Scholar] [CrossRef]
- Ellis, K.J.; Morrison, J.F. Buffers of constant ionic strength for studying pH-dependent processes. Methods Enzymol. 1982, 87, 405–426. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef]
- Costas, A.M.; White, A.K.; Metcalf, W.W. Purification and characterization of a novel phosphorus-oxidizing enzyme from Pseudomonas stutzeri WM88. J. Biol. Chem. 2001, 276, 17429–17436. [Google Scholar] [CrossRef] [Green Version]
- Ebel, C.; Faou, P.; Kernel, B.; Zaccai, G. Relative role of anions and cations in the stabilization of halophilic malate dehydrogenase. Biochemistry 1999, 38, 9039–9047. [Google Scholar] [CrossRef]
- Beneventi, E.; Niero, M.; Motterle, R.; Fraaije, M.; Bergantino, E. Discovery of Baeyer-Villiger monooxygenases from photosynthetic eukaryotes. J. Mol. Catal. B Enzym. 2013, 98, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Zaccai, G.; Eisenberg, H. Halophilic proteins and the influence of solvent on protein stabilization. Trends Biochem. Sci. 1990, 15, 333–337. [Google Scholar] [CrossRef]
- Castle, A.M.; Macnab, R.M.; Shulman, R.G. Measurement of intracellular sodium concentration and sodium transport in Escherichia coli by 23Na nuclear magnetic resonance. J. Biol. Chem. 1986, 261, 3288–3294. [Google Scholar] [PubMed]
- Bonete, M.-J.; Pire, C.; Llorca, F.I.; Camacho, M.L. Glucose dehydrogenase from the halophilic Archaeon Haloferax mediterranei: Enzyme purification, characterisation and N-terminal sequence. FEBS Lett. 1996, 383, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Torres, S.; Castro, G.R. Non-aqueous biocatalysis in homogeneous solvent systems. Food Technol. Biotechnol. 2004, 42, 271–277. [Google Scholar]
- Ruiz, D.M.; De Castro, R.E. Effect of organic solvents on the activity and stability of an extracellular protease secreted by the haloalkaliphilic archaeon Natrialba magadii. J. Ind. Microbiol. Biotechnol. 2007, 34, 111–115. [Google Scholar] [CrossRef]
- Elbanna, K.; Ibrahim, I.M.; Revol-Junelles, A.-M. Purification and characterization of halo-alkali-thermophilic protease from Halobacterium. Extremophiles 2015, 19, 763–774. [Google Scholar] [CrossRef]
- Munawar, N.; Engel, P.C. Overexpression in a non-native halophilic host and biotechnological potential of NAD-dependent glutamate dehydrogenase from Halobacterium salinarum strain NRC-36014. Extremophiles 2012, 16, 463–476. [Google Scholar] [CrossRef]
- Alsafadi, D.; Paradisi, F. Effect of organic solvents on the activity and stability of halophilic alcohol dehydrogenase (ADH2) from Haloferax volcanii. Extremophiles 2013, 17, 115–122. [Google Scholar] [CrossRef]
- Fukushima, T.; Mizuki, T.; Echigo, A.; Inoue, A.; Usami, R. Organic solvent tolerance of halophilic α-amylase from a Haloarchaeon, Haloarcula sp. strain S-1. Extremophiles 2005, 9, 85–89. [Google Scholar] [CrossRef]
- Chang, J.; Lee, Y.-S.; Fang, S.-J. Recombinant expression and characterization of an organic-solvent-tolerant α-amylase from Exiguobacterium sp. DAU5. Appl. Biochem. Biotechnol. 2013, 169, 1870–1883. [Google Scholar] [CrossRef] [PubMed]
- Opperman, D.J.; Reetz, M.T. Towards Practical Baeyer–Villiger-Monooxygenases: Design of Cyclohexanone Monooxygenase Mutants with Enhanced Oxidative Stability. Chembiochem 2010, 11, 2589–2596. [Google Scholar] [CrossRef] [PubMed]
- Van Beek, H.L.; Wijma, H.J.; Fromont, L.; Janssen, D.B.; Fraaije, M.W. Stabilization of cyclohexanone monooxygenase by a computationally designed disulfide bond spanning only one residue. FEBS Open Biol. 2014, 4, 168–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, S.; Genz, M.; Balke, K.; Bornscheuer, U.T. The effect of disulfide bond introduction and related Cys/Ser mutations on the stability of a cyclohexanone monooxygenase. J. Biotechnol. 2015, 214, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- E. coli Codon Usage Analyzer 2.1. Available online: http://www.faculty.ucr.edu (accessed on 11 December 2019).
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools on the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- UCSF Chimera. Available online: http://www.cgl.ucsf.edu/chimera/ (accessed on 11 December 2019).
- Dolinsky, T.J.; Czodrowski, P.; Li, H.; Nielsen, J.E.; Jensen, J.H.; Klebe, G.; Baker, N.A. PDB2PQR: Expanding and upgrading automated preparation of biomolecular structures for molecular simulations. Nucleic Acids Res. 2007, 35, W522–W525. [Google Scholar] [CrossRef]
- Sitkoff, D.; Sharp, K.A.; Honig, B. Accurate calculation of hydration free energies using macroscopic solvent models. J. Phys. Chem. 1994, 98, 1978–1988. [Google Scholar] [CrossRef]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [Green Version]
- Opal Web Service. Available online: http://www.poissonboltzmann.org/apbs (accessed on 11 December 2019).
- Schutz, C.N.; Warshel, A. What are the dielectric “constants” of proteins and how to validate electrostatic models? Proteins 2001, 44, 400–417. [Google Scholar] [CrossRef] [PubMed]
- Gorham, R.D., Jr.; Kieslich, C.A.; Morikis, D. Electrostatic clustering and free energy calculations provide a foundation for protein design and optimization. Ann. Biomed. Eng. 2011, 39, 1252–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.K.; Fitch, C.A.; Garcìa-Moreno, E.B. Distance dependence and salt sensitivity of pairwise, coulombic interactions in a protein. Protein Sci. 2002, 11, 1004–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WebPIPSA Server. Available online: http://pipsa.eml.org/pipsa (accessed on 11 December 2019).
- Fraczkiewicz, R.; Braun, W. Exact and efficient analytical calculation of the accessible surface areas and their gradients for macromolecules. J. Comput. Chem. 1998, 19, 319–333. [Google Scholar] [CrossRef]
- Krissinel, E. On the relationship between sequence and structure similarities in proteomics. Bioinformatics 2007, 23, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraaije, M.W.; Wu, J.; Heuts, D.P.H.M.; van Hellemond, E.W.; Spelberg, J.H.; Janssen, D.B. Discovery of a thermostable Baeyer–Villiger monooxygenase by genome mining. Appl. Microbiol. Biotechnol. 2005, 66, 393–400. [Google Scholar] [CrossRef]
- K2D3. Available online: http://cbdm-01.zdv.uni-mainz.de/~andrade/k2d3// (accessed on 11 December 2019).
- Louis-Jeune, C.; Andrade-Navarro, M.A.; Perez-Iratxeta, C. Prediction of protein secondary structure from circular dichroism using theoretically derived spectra. Proteins 2012, 80, 374–381. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Halophilic | Thermophilic | Mesophilic | ||
|---|---|---|---|---|
| HtBVMO | HmMDH | PAMO | RhCHMO | |
| Amino acid frequencies (% of total residues) | ||||
| Asp+Glu | 21.66 | 20.39 | 13.84 | 14.44 |
| Arg+Lys | 7.84 | 7.57 | 10.30 | 8.89 |
| Hydrophobic residues | 41.70 | 49.67 | 47.20 | 51.85 |
| Amino acid ratios | ||||
| (Asp+Glu)/(Arg+Lys) | 2.73 | 2.70 | 1.34 | 1.63 |
| Charged/hydrophobic | 0.52 | 0.61 | 0.51 | 0.50 |
| Accessible surface (Å2) | ||||
| Total | 22,733.40 | 13,352.55 | 22,158.52 | 21,277.37 |
| Asp+Glu | 9648.94 | 4880.15 | 5403.37 | 6154.71 |
| Arg+Lys | 3040.31 | 1744.19 | 5143.93 | 4379.64 |
| Hydrophobic residues | 4536.05 | 3407.03 | 6413.00 | 6134.00 |
| Amino acid surface ratios | ||||
| (Asp+Glu)/(Arg+Lys) surface ratio | 2.88 | 2.80 | 1.05 | 1.41 |
| Charged/hydrophobic surface ratio | 2.73 | 1.94 | 1.64 | 1.41 |
| % of total surface | ||||
| Asp+Glu | 42.44 | 36.55 | 24.39 | 28.93 |
| Arg+Lys | 14.72 | 13.06 | 23.21 | 20.58 |
| Hydrophobic residues | 19.95 | 25.52 | 28.94 | 28.83 |
| Substrate | KM (mM) | kcat (s−1) | kcat/KM (s−1 mM−1) | Product |
|---|---|---|---|---|
heptan-2-one | 0.09 ± 0.01 | 0.277 ± 0.005 | 3.08 ± 0.35 | pentyl acetate |
octan-2-one | 0.7 ± 0.1 | 0.27 ± 0.02 | 0.38 ± 0.06 | hexyl acetate |
nonan-3-one | 4.5 ± 1.3 | 0.12 ± 0.01 | 0.260 ± 0.008 | hexyl propionate |
1-phenylpropan-2-one | no reaction | no reaction | no reaction | no product |
4-phenylbutan-2-one | 0.017 ± 0.002 | 0.155 ± 0.003 | 8.90 ± 0.89 | 2-phenylethyl acetate |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niero, M.; Righetto, I.; Beneventi, E.; Polverino de Laureto, P.; Fraaije, M.W.; Filippini, F.; Bergantino, E. Unique Features of a New Baeyer–Villiger Monooxygenase from a Halophilic Archaeon. Catalysts 2020, 10, 128. https://doi.org/10.3390/catal10010128
Niero M, Righetto I, Beneventi E, Polverino de Laureto P, Fraaije MW, Filippini F, Bergantino E. Unique Features of a New Baeyer–Villiger Monooxygenase from a Halophilic Archaeon. Catalysts. 2020; 10(1):128. https://doi.org/10.3390/catal10010128
Chicago/Turabian StyleNiero, Mattia, Irene Righetto, Elisa Beneventi, Patrizia Polverino de Laureto, Marco Wilhelmus Fraaije, Francesco Filippini, and Elisabetta Bergantino. 2020. "Unique Features of a New Baeyer–Villiger Monooxygenase from a Halophilic Archaeon" Catalysts 10, no. 1: 128. https://doi.org/10.3390/catal10010128
APA StyleNiero, M., Righetto, I., Beneventi, E., Polverino de Laureto, P., Fraaije, M. W., Filippini, F., & Bergantino, E. (2020). Unique Features of a New Baeyer–Villiger Monooxygenase from a Halophilic Archaeon. Catalysts, 10(1), 128. https://doi.org/10.3390/catal10010128