High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism

Abstract
:1. Introduction
2. Results
2.1. Reference Cultivation with Pure Glycerol
2.2. Cultivation with Crude Glycerol
2.2.1. Cultivation with Crude Glycerol in 100 mL Laboratory Glass Bottles
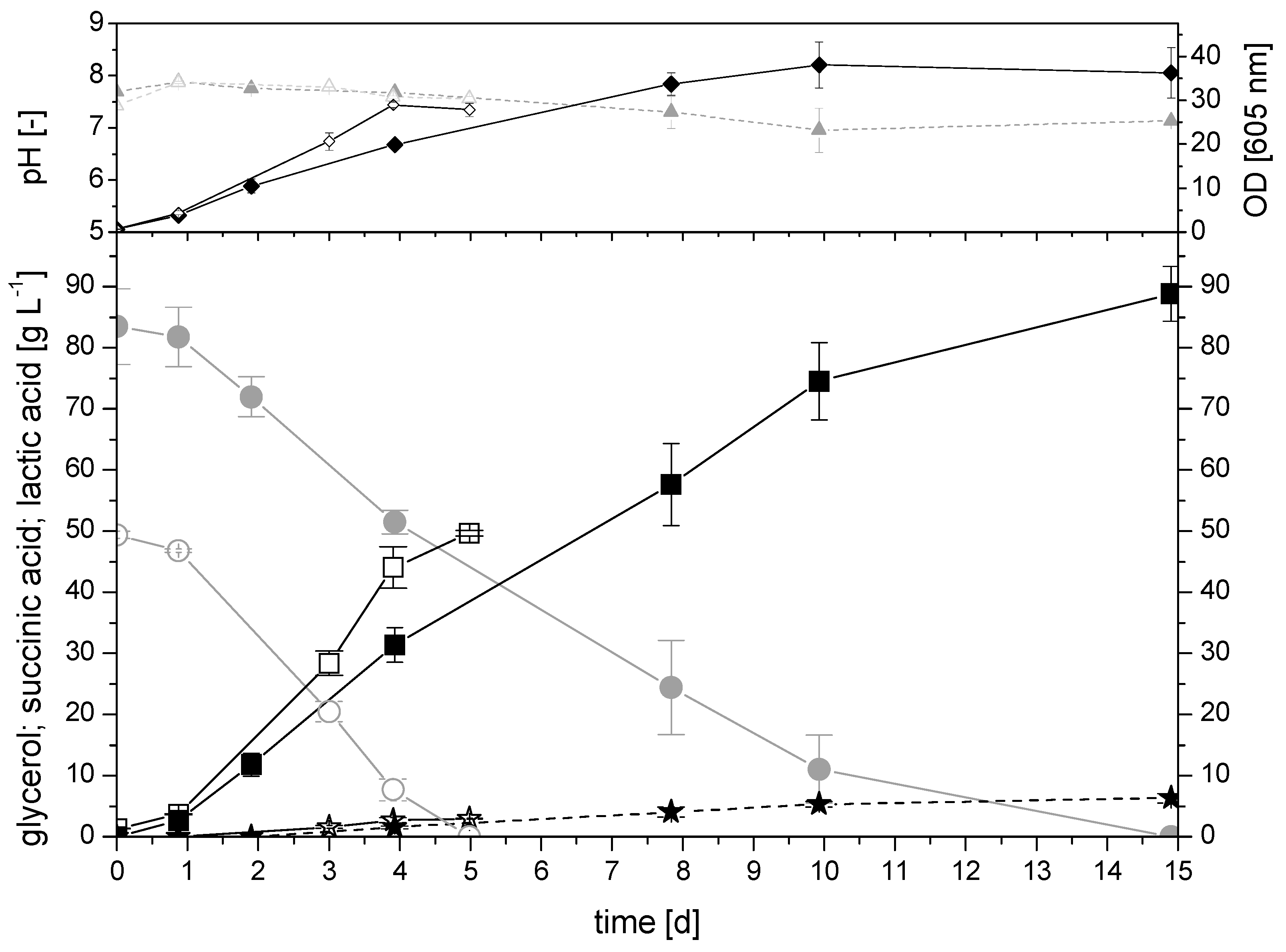
2.2.2. Cultivation with Crude Glycerol in the 350 mL Bioreactor System Compared to Pure Glycerol
2.3. Crystallization and Purity Determination
3. Discussion
4. Materials and Methods
4.1. Microorganism
4.2. Media composition and Chemicals
4.3. Cultivation
4.4. Analytical Methods
4.5. Crystallization and Purity Determination
Author Contributions
Funding
Conflicts of Interest
References
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—The us department of energy’s “top 10” revisited. Green Chem. 2010, 12, 539. [Google Scholar] [CrossRef]
- Ferone, M.; Raganati, F.; Ercole, A.; Olivieri, G.; Salatino, P.; Marzocchella, A. Continuous succinic acid fermentation by Actinobacillus succinogenes in a packed-bed biofilm reactor. Biotechnol. Biofuels 2018, 11, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinlay, J.B.; Vieille, C.; Zeikus, J.G. Prospects for a bio-based succinate industry. Appl. Microbiol. Biotechnol. 2007, 76, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Bechthold, I.; Bretz, K.; Kabasci, S.; Kopitzky, R.; Springer, A. Succinic acid: A new platform chemical for biobased polymers from renewable resources. Chem. Eng. Technol. 2008, 31, 647–654. [Google Scholar] [CrossRef]
- Bradfield, M.F.; Mohagheghi, A.; Salvachua, D.; Smith, H.; Black, B.A.; Dowe, N.; Beckham, G.T.; Nicol, W. Continuous succinic acid production by Actinobacillus succinogenes on xylose-enriched hydrolysate. Biotechnol. Biofuels 2015, 8, 181. [Google Scholar] [CrossRef] [Green Version]
- Cukalovic, A.; Stevens, C.V. Feasibility of production methods for succinic acid derivatives: A marriage of renewable resources and chemical technology. Biofuels Bioprod. Biorefin. 2008, 2, 505–529. [Google Scholar] [CrossRef]
- Cornils, B.; Lappe, P. Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag: Weinheim, Germany, 2010; pp. 287–304. [Google Scholar]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2019. Safeguarding Against Economic Slowdowns and Downturns; Licence: CC BY-NC-SA 3.0 IGO; FAO: Rome, Italy, 2019. [Google Scholar]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef]
- Pateraki, C.; Patsalou, M.; Vlysidis, A.; Kopsahelis, N.; Webb, C.; Koutinas, A.A.; Koutinas, M. Actinobacillus succinogenes: Advances on succinic acid production and prospects for development of integrated biorefineries. Biochem. Eng. J. 2016, 112, 285–303. [Google Scholar] [CrossRef]
- Herselman, J.; Bradfield, M.F.A.; Vijayan, U.; Nicol, W. The effect of carbon dioxide availability on succinic acid production with biofilms of Actinobacillus succinogenes. Biochem. Eng. J. 2017, 117, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzyme Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Cheng, K.-K.; Zhao, X.-B.; Zeng, J.; Zhang, J.-A. Biotechnological production of succinic acid: Current state and perspectives. Biofuels Bioprod. Biorefin. 2012, 6, 302–318. [Google Scholar] [CrossRef]
- Chen, Y.; Nielsen, J. Advances in metabolic pathway and strain engineering paving the way for sustainable production of chemical building blocks. Curr. Opin. Biotechnol. 2013, 24, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Guo, F.; Zhang, S.; Zhang, W.; Yang, Q.; Dong, W.; Jiang, M.; Ma, J.; Xin, F. Bio-based succinic acid: An overview of strain development, substrate utilization, and downstream purification. Biofuels Bioprod. Biorefin. 2020. [Google Scholar] [CrossRef]
- Tan, J.P.; Jahim, J.M.; Harun, S.; Wu, T.Y. Overview of the potential of bio-succinic acid production from oil palm fronds. J. Phys. Sci. 2017, 28, 53–72. [Google Scholar]
- Ahn, J.H.; Jang, Y.-S.; Lee, S.Y. Production of succinic acid by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 2016, 42, 54–66. [Google Scholar] [CrossRef]
- Li, J.; Jiang, M.; Chen, K.; Shang, L.; Wei, P.; Ying, H.; Ye, Q.; Ouyang, P.; Chang, H. Enhanced production of succinic acid by Actinobacillus succinogenes with reductive carbon source. Process Biochem. 2010, 45, 980–985. [Google Scholar] [CrossRef]
- Biobasierte Chemikalien|chemanager-online.com—Chemie und Life Science. Available online: https://www.chemanager-online.com/themen/chemikalien-distribution/biobasierte-chemikalien (accessed on 12 August 2019).
- Nghiem, N.P.; Kleff, S.; Schwegmann, S. Succinic acid: Technology development and commercialization. Fermentation 2017, 3, 26. [Google Scholar] [CrossRef]
- Li, Q.; Wang, D.; Wu, Y.; Li, W.L.; Zhang, Y.J.; Xing, J.M.; Su, Z.G. One step recovery of succinic acid from fermentation broths by crystallization. Sep. Purif. Technol. 2010, 72, 294–300. [Google Scholar] [CrossRef]
- Tan, J.P.; Md Jahim, J.; Wu, T.Y.; Harun, S.; Kim, B.H.; Mohammad, A.W. Insight into bio-mass as a renewable carbon source for the production of succinic acid and the factors affecting the metabolic flux toward higher succinate yield. Industrial & Engineering. Chem. Res. 2014, 53, 16123–16134. [Google Scholar]
- Blankschien, M.D.; Clomburg, J.M.; Gonzalez, R. Metabolic engineering of Escherichia coli for the production of succinate from glycerol. Metab. Eng. 2010, 12, 409–419. [Google Scholar] [CrossRef]
- Li, C.; Yang, X.; Gao, S.; Wang, H.; Lin, C.S.K. High efficiency succinic acid production from glycerol via in situ fibrous bed bioreactor with an engineered Yarrowia lipolytica. Bioresour. Technol. 2017, 225, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.J.; Yang, X.F.; Wang, H.M.; Rivero, C.P.; Li, C.; Cui, Z.Y.; Qi, Q.S.; Lin, C.S.K. Robust succinic acid production from crude glycerol using engineered Yarrowia lipolytica. Biotechnol. Biofuels 2016, 9, 179. [Google Scholar] [CrossRef]
- Yocum, R.R.; Hermann, T.; Hu, X. Fermentation of Glycerol to Organic Acids. U.S. Patent 2014 0234923 A1, 21 August 2014. [Google Scholar]
- Wang, C.; Zhang, H.; Cai, H.; Zhou, Z.; Chen, Y.; Chen, Y.; Ouyang, P. Succinic acid production from corn cob hydrolysates by genetically engineered Corynebacterium glutamicum. Appl. Biochem. Biotechnol. 2014, 172, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.; Haefner, S.; Von Abendroth, G.; Hollman, R.; Raddatz, A.; Ernst, H.; Gurski, H. Novel Microbial Succinic Acid Producers and Purification of Succinic Acid. U.S. Patent 2014 0127764 A1, 8 Jan 2014. [Google Scholar]
- Okino, S.; Noburyu, R.; Suda, M.; Jojima, T.; Inui, M.; Yukawa, H. An efficient succinic acid production process in a metabolically engineered Corynebacterium glutamicum strain. Appl. Microbiol. Biotechnol. 2008, 81, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Succinate production in dual-phase Escherichia coli fermentations depends on the time of transition from aerobic to anaerobic conditions. J. Ind. Microbiol. Biotechnol. 2002, 28, 325–332. [Google Scholar] [CrossRef]
- Li, Q.; Huang, B.; He, Q.; Lu, J.; Li, X.; Li, Z.; Wu, H.; Ye, Q. Production of succinate from simply purified crude glycerol by engineered Escherichia coli using two-stage fermentation. Bioresour. Bioprocess. 2018, 5, 41. [Google Scholar] [CrossRef]
- Guettler, M.V.; Rumler, D.; Jain, M.K. Actinobacillus succinogenes sp. Nov., a novel succinic-acid-producing strain from the bovine rumen. Int. J. Syst. Bacteriol. 1999, 49, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Bradfield, M.F.A.; Nicol, W. Continuous succinic acid production from xylose by Actinobacillus succinogenes. Bioproc. Biosyst. Eng. 2016, 39, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Stellmacher, R.; Hangebrauk, J.; von Abendroth, G.; Scholten, E.; Wittmann, C. Fermentative production of succinic acid with Basfia succiniciproducens. Chem. Ing. Tech. 2010, 82, 1223–1229. [Google Scholar] [CrossRef]
- Scholten, E.; Dagele, D. Succinic acid production by a newly isolated bacterium. Biotechnol. Lett. 2008, 30, 2143–2146. [Google Scholar] [CrossRef]
- Vlysidis, A.; Binns, M.; Webb, C.; Theodoropoulos, C. Glycerol utilisation for the production of chemicals: Conversion to succinic acid, a combined experimental and computational study. Biochem. Eng. J. 2011, 58–59, 1–11. [Google Scholar] [CrossRef]
- Shen, N.; Qin, Y.; Wang, Q.; Liao, S.; Zhu, J.; Zhu, Q.; Mi, H.; Adhikari, B.; Wei, Y.; Huang, R. Production of succinic acid from sugarcane molasses supplemented with a mixture of corn steep liquor powder and peanut meal as nitrogen sources by Actinobacillus succinogenes. Lett. Appl. Microbiol. 2015, 60, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Bromann, S.; Goy, K.; Kuenz, A.; Prüße, U.; Willke, T. Schlussbericht zum Vorhaben Biobst—Biotechnische Herstellung von Bernsteinsäure aus Nachwachsenden Rohstoffen (Biotechnological Production of Succinic Acid from Renewable Resources); Laufzeit: 01.09.2015 bis 15.10.2018; Johann Heinrich von Thünen-Institut: Braunschweig, Germany, 2019. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Reference Cultivation in 100 mL Laboratory Glass Bottle | ||
|---|---|---|
| Initial 50 g L−1 Substrate | Initial 85 g L−1 Substrate | |
| CEnd, SA / g L−1 | 49.4 | 88.8 |
| CEnd, LA / g L−1 | 3.2 | 6.3 |
| Pmax / g L−1 h−1 | 0.76 | 0.31 |
| P / g L−1 h−1 | 0.37 | 0.22 |
| Y1 / g g−1 | 0.9 | 0.94 |
| Pure Glycerol | Crude Glycerol | |
|---|---|---|
| CEnd, SA / g L−1 | 49.4 | 46.8 |
| CEnd, LA / g L−1 | 3.2 | 2.3 |
| Pmax / g L−1 h−1 | 0.76 | 0.62 |
| P / g L−1 h−1 | 0.37 | 0.35 |
| Y1 / g g−1 | 0.9 | 0.88 |
| Pure Glycerol | Crude Glycerol | |
|---|---|---|
| CEnd, SA / g L−1 | 117 | 86.9 |
| CEnd, LA / g L−1 | 8.6 | 6 |
| Pmax / g L−1 h−1 | 0.69 | 0.55 |
| P / g L−1 h−1 | 0.34 | 0.33 |
| Y1 / g g−1 | 1.3 | 0.9 |
| Microorganism | Substrate | Succinic Acid | Yield | Productivity | Reference |
|---|---|---|---|---|---|
| (g L−1) | (g g−1) | (g L−1h−1) | |||
| Actinobacillus succinogenes 130Z | Glucose | 106 | 0.8 | 1.34 | [32] |
| Xylose1 | 29.4 | 0.68 | 3.41 | [33] | |
| Basfia succiniciproducens DD1 | Glucose | 31.7 | 0.63 | 1.32 | [34] |
| Basfia succiniciproducens DD1 | Crude glycerol | 8.4 | 1.2 | 0.9 | [35] |
| Actinobacillus succinogenes ATCC 55618 | Glycerol | 29.3 | 1.23 | 0.27 | [36] |
| Actinobacillus succinogenes GXAS 137 | Sugar cane molasses | 64.7 | 0.86 | 1.4 | [37] |
| Isolate AKR177 | Glycerol | 117 | 1.3 | 0.34 | This study |
| Isolate AKR177 | Crude glycerol | 86.9 | 0.9 | 0.33 | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuenz, A.; Hoffmann, L.; Goy, K.; Bromann, S.; Prüße, U. High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism. Catalysts 2020, 10, 470. https://doi.org/10.3390/catal10050470
Kuenz A, Hoffmann L, Goy K, Bromann S, Prüße U. High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism. Catalysts. 2020; 10(5):470. https://doi.org/10.3390/catal10050470
Chicago/Turabian StyleKuenz, Anja, Lisa Hoffmann, Katharina Goy, Sarah Bromann, and Ulf Prüße. 2020. "High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism" Catalysts 10, no. 5: 470. https://doi.org/10.3390/catal10050470
APA StyleKuenz, A., Hoffmann, L., Goy, K., Bromann, S., & Prüße, U. (2020). High-Level Production of Succinic Acid from Crude Glycerol by a Wild Type Organism. Catalysts, 10(5), 470. https://doi.org/10.3390/catal10050470