Penicillin Acylase from Streptomyces lavendulae and Aculeacin A Acylase from Actinoplanes utahensis: Two Versatile Enzymes as Useful Tools for Quorum Quenching Processes

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Substrate specificity of SlPA and AuAAC towards different AHLs
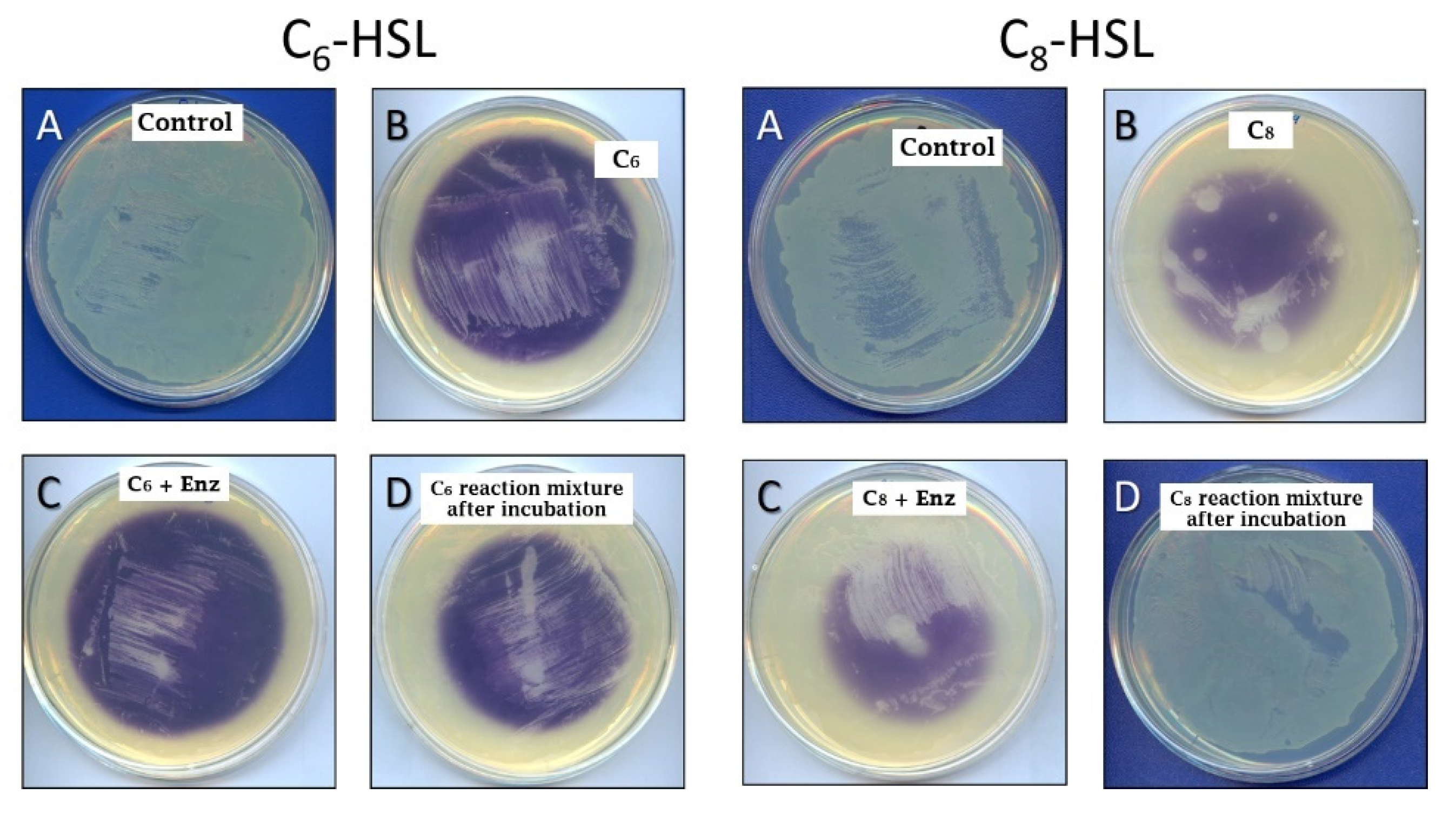
2.2. Quorum Quenching Role of SlPA and AuAAC
2.3. Phylogenetic Analysis of SlPA and AuAAC Acylases
3. Materials and Methods
3.1. Materials and Bacterial Strains
3.2. Synthesis of N-Acyl-Homoserine Lactones
3.3. Determination of Kinetic Parameters
3.4. Inhibition of Violacein Production by Chromobacterium Violaceum CV026
3.5. Inhibition of Biofilm Formation by Pseudomonas Aeruginosa on Glass Slides
3.6. Protein Sequence Alignments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- de Kievit, T.R.; Iglewski, B.H. Bacterial quorum sensing in pathogenic relationships. Infect. Immun. 2000, 68, 4839–4849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Gao, Y.X.; Chen, X.Y.; Yu, Z.M.; Li, X.Z. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing dependent infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef]
- Whitehead, N.A.; Welch, M.; Salmond, G.P.C. Silencing the majority. Nat. Biotechnol. 2001, 19, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Soler, A.; Arregui, L.; Arroyo, M.; Mendoza, J.A.; Muras, A.; Alvarez, C.; Garcia-Vera, C.; Marquina, D.; Santos, A.; Serrano, S. Quorum sensing versus quenching bacterial isolates obtained from mbr plants treating leachates from municipal solid waste. Int. J. Environ. Res. Public Health 2018, 15, 1019. [Google Scholar] [CrossRef] [Green Version]
- Dickschat, J.S. Quorum sensing and bacterial biofilms. Nat. Prod. Rep. 2010, 27, 343–369. [Google Scholar] [CrossRef]
- Whitehead, N.A.; Barnard, A.M.L.; Slater, H.; Simpson, N.J.L.; Salmond, G.P.C. Quorum sensing in Gram-negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [Green Version]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum quenching: Role in nature and applied developments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef]
- Fetzner, S. Quorum quenching enzymes. J. Biotechnol. 2015, 201, 2–14. [Google Scholar] [CrossRef]
- Czajkowski, R.; Jafra, S. Quenching of acyl-homoserine lactone-dependent quorum sensing by enzymatic disruption of signal molecules. Acta Biochim. Pol. 2009, 56, 1–16. [Google Scholar] [CrossRef]
- Yates, E.A.; Philipp, B.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y.; Cámara, M.; Smith, H.; et al. N-acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun. 2002, 70, 5635–5646. [Google Scholar] [CrossRef] [Green Version]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusada, H.; Zhang, Y.; Tamaki, H.; Kimura, N.; Kamagata, Y. Novel N-acyl Homoserine lactone-degrading bacteria isolated from penicillin-contaminated environments and their quorum-quenching activities. Front. Microbiol. 2019, 10, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bzdrenga, J.; Daude, D.; Remy, B.; Jacquet, P.; Plener, L.; Elias, M.; Chabriere, E. Biotechnological applications of quorum quenching enzymes. Chem. Biol. Interact. 2017, 267, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.W.; Koh, C.L.; Sam, C.K.; Yin, W.F.; Chan, K.G. Quorum quenching revisited—From signal decays to signalling confusion. Sensors 2012, 12, 4661–4696. [Google Scholar] [CrossRef] [Green Version]
- Oinonen, C.; Rouvinen, J. Structural comparison of Ntn-hydrolases. Protein. Sci. 2000, 9, 2329–2337. [Google Scholar] [CrossRef] [Green Version]
- Utari, P.D.; Vogel, J.; Quax, W.J. Deciphering physiological functions of AHL quorum quenching acylases. Front. Microbiol. 2017, 8, 1123. [Google Scholar] [CrossRef] [Green Version]
- Mukherji, R.; Varshney, N.K.; Panigrahi, P.; Suresh, C.G.; Prabhune, A. A new role for penicillin acylases: Degradation of acyl homoserine lactone quorum sensing signals by Kluyvera citrophila penicillin G acylase. Enzym. Microb. Technol. 2014, 56, 1–7. [Google Scholar] [CrossRef]
- Sunder, A.V.; Utari, P.D.; Ramasamy, S.; van Merkerk, R.; Quax, W.; Pundle, A. Penicillin V acylases from gram-negative bacteria degrade N-acylhomoserine lactones and attenuate virulence in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2017, 101, 2383–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Guzmán, R.; de la Mata, I.; Torres-Bacete, J.; Arroyo, M.; Castillón, M.P.; Acebal, C. Substrate specificity of penicillin acylase from Streptomyces lavendulae. Biochem. Biophys. Res. Commun. 2002, 291, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Torres-Bacete, J.; Hormigo, D.; Torres-Gúzman, R.; Arroyo, M.; Castillón, M.P.; García, J.L.; Acebal, C.; de la Mata, I. Overexpression of penicillin V acylase from Streptomyces lavendulae and elucidation of its catalytic residues. Appl. Environ. Microbiol. 2015, 81, 1225–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshima, H.; Inokoshi, J.; Takada, Y.; Tanaka, H.; Omura, S. A deacylation enzyme for aculeacin A, a neutral lipopeptide antibiotic; from Actinoplanes utahensis: Purification and characterization. J. Biochem. 1989, 105, 606–610. [Google Scholar] [CrossRef] [Green Version]
- Torres-Bacete, J.; Hormigo, D.; Stuart, M.; Arroyo, M.; Torres, P.; Castillón, M.P.; Acebal, C.; García, J.L.; de la Mata, I. Newly discovered penicillin acylase activity of aculeacin A acylase from Actinoplanes utahensis. Appl. Environ. Microbiol. 2007, 73, 5378–5381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, M.; Torres, R.; de la Mata, I.; Castillón, M.P.; Acebal, C. Interaction of penicillin V acylase with organic solvents: Catalytic activity modulation on the hydrolysis of penicillin V. Enzym. Microb. Technol. 1999, 25, 378–383. [Google Scholar] [CrossRef]
- Arroyo, M.; Torres-Guzman, R.; de la Mata, I.; Castillon, M.P.; Acebal, C. A kinetic examination of penicillin acylase stability in water-organic solvent systems at different temperatures. Biocatal. Biotransform. 2002, 20, 53–56. [Google Scholar] [CrossRef]
- Arroyo, M.; Torres-Guzman, R.; de la Mata, I.; Castillon, M.P.; Acebal, C. Prediction of penicillin V acylase stability in water-organic co-solvent monophasic systems as a function of solvent composition. Enzym. Microb. Technol. 2000, 27, 122–126. [Google Scholar] [CrossRef]
- Hormigo, D.; de la Mata, I.; Castillón, M.P.; Acebal, C.; Arroyo, M. Kinetic and microstructural characterization of immobilized penicillin acylase from Streptomyces lavendulae on Sepabeads EC-EP. Biocatal Biotransform. 2009, 27, 271–281. [Google Scholar] [CrossRef]
- Hormigo, D.; de la Mata, I.; Acebal, C.; Arroyo, M. Immobilized aculeacin A acylase from Actinoplanes utahensis: Characterization of a novel biocatalyst. Bioresour. Technol. 2010, 101, 4261–4268. [Google Scholar] [CrossRef]
- Hormigo, D.; López-Conejo, M.T.; Serrano-Aguirre, L.; García-Martín, A.; Saborido, A.; de la Mata, I.; Arroyo, M. Kinetically controlled acylation of 6-APA catalyzed by penicillin acylase from Streptomyces lavendulae: Effect of reaction conditions in the enzymatic synthesis of penicillin V. Biocatal. Biotransform. 2020, 38, 253–262. [Google Scholar] [CrossRef]
- Park, S.Y.; Kang, H.O.; Jang, H.S.; Lee, J.K.; Koo, B.T.; Yum, D.Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef] [Green Version]
- Morohoshi, T.; Nakazawa, S.; Ebata, A.; Kato, N.; Ikeda, T. Identification and characterization of N-acylhomoserine lactone-acylase from the fish intestinal Shewanella sp. strain MIB015. Biosci. Biotechnol. Biochem. 2008, 72, 1887–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochiai, S.; Yasumoto, S.; Morohoshi, T.; Ikeda, T. AmiE, a novel N-acylhomoserine lactone acylase belonging to the amidase family, from the activated-sludge isolate Acinetobacter sp. strain Ooi24. Appl. Environ. Microbiol. 2014, 80, 6919–6925. [Google Scholar] [CrossRef] [Green Version]
- Kusada, H.; Tamaki, H.; Kamagata, Y.; Hanada, S.; Kimura, N. A novel quorum quenching N-acylhomoserine lactone acylase from Acidovorax sp Strain MR-S7 mediates antibiotic resistance. Appl. Environ. Microbiol. 2017, 83, e00080–e00117. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Xu, J.L.; Hu, J.; Wang, L.H.; Ong, S.L.; Leadbetter, J.R.; Zhang, L.H. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum quenching enzymes. Mol. Microbiol. 2003, 47, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Sio, C.F.; Otten, L.G.; Cool, R.H.; Diggle, S.P.; Braun, P.G.; Bos, R.; Daykin, M.; Cámara, M.; Williams, P.; Quax, W.J. Quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas aeruginosa PAO1. Infect. Immun. 2006, 74, 1673–1682. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.J.; Petersen, A.; Whiteley, M.; Leadbetter, J.R. Identification of QuiP, the product of gene PA1032, as the second acyl-homoserine lactone acylase of Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2006, 72, 1190–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, R.W.; Lindow, S.E. Two dissimilar N-acyl-homoserine lactone acylases of Pseudomonas syringae influence colony and biofilm morphology. Appl. Environ. Microbiol. 2009, 75, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, M.; Diggle, S.P.; Heeb, S.; Cámara, M.; Otero, A. Quorum quenching activity in Anabaena sp. PCC 7120: Identification of AiiC, a novel AHL-acylase. FEMS Microbiol. Lett. 2008, 280, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.; Nadal-Jimenez, P.; Cool, R.H.; Quax, W.J. Deinococcus radiodurans can interfere with quorum sensing by producing an AHL-acylase and an AHL-lactonase. FEMS Microbiol. Lett. 2014, 356, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Wahjudi, M.; Papaioannou, E.; Hendrawati, O.; van Assen, A.H.G.; van Merkerk, R.; Cool, R.H.; Poelarends, G.J.; Quax, W.J. PA0305 of Pseudomonas aeruginosa is a quorum quenching acylhomoserine lactone acylase belonging to the Ntn hydrolase superfamily. Microbiology 2011, 157, 2042–2055. [Google Scholar] [CrossRef] [Green Version]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 1998, 280, 295–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohler, T.; Curty, L.K.; Barja, F.; van Delden, C.; Pechere, J.C. Swarming of Pseudomonas aeruginosa is dependent on cell-to-cell signaling and requires flagella and pili. J. Bacteriol. 2000, 182, 5990–5996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, S.Q.; Zhou, L.J.; Zhang, Z.Q.; Li, J.X. Influence and mechanism of N-(3-oxooctanoyl)-L-homoserine lactone (C8-oxo-HSL) on biofilm behaviors at early stage. J. Environ. Sci. 2012, 24, 2035–2040. [Google Scholar] [CrossRef]
- Kuipers, R.K.; Joosten, H.J.; van Berkel, W.J.; Leferink, N.G.; Rooijen, E.; Ittmann, E.; van Zimmeren, F.; Jochens, H.; Bornscheuer, U.; Vriend, G.; et al. 3DM: Systematic analysis of heterogeneous superfamily data to discover protein functionalities. Proteins Struct. Funct. Bioinform. 2010, 78, 2101–2113. [Google Scholar] [CrossRef] [PubMed]
- van den Bergh, T.; Tamo, G.; Nobili, A.; Tao, Y.; Tan, T.; Bornscheuer, U.T.; Kuipers, R.K.P.; Vroling, B.; de Jong, R.M.; Subramanian, K.; et al. CorNet: Assigning function to networks of co-evolving residues by automated literature mining. PLoS ONE 2017, 12, e0176427. [Google Scholar] [CrossRef]
- Yasutake, Y.; Kusada, H.; Ebuchi, T.; Hanada, S.; Kamagata, Y.; Tamura, T.; Kimura, N. Bifunctional quorum quenching and antibiotic acylase MacQ forms a 170-kDa capsule-shaped molecule containing spacer polypeptides. Sci. Rep. 2017, 7, 8946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, S.; Shibata, T.; Ito, K.; Oohata, N.; Yamashita, M.; Hino, M.; Yamada, M.; Isogai, Y.; Hashimoto, S. Cloning and expression of the FR901379 acylase gene from Streptomyces sp. no. 6907. J. Antibiot. 2011, 64, 169–175. [Google Scholar] [CrossRef]
- Zhang, D.; Koreishi, M.; Imanaka, H.; Imamura, K.; Nakanishi, K. Cloning and characterization of penicillin V acylase from Streptomyces mobaraensis. J. Biotechnol. 2007, 128, 788–800. [Google Scholar] [CrossRef]
- Terwagne, M.; Mirabella, A.; Lemaire, J.; Deschamps, C.; De Bolle, X.; Letesson, J.J. Quorum sensing and self-quorum quenching in the intracellular pathogen Brucella melitensis. PLoS ONE 2013, 8, e82514. [Google Scholar] [CrossRef]
- Liu, N.; Yu, M.; Zhao, Y.B.; Cheng, J.G.; An, K.; Zhang, X.H. PfmA, a novel quorum quenching N-acylhomoserine lactone acylase from Pseudoalteromonas flavipulchra. Microbiology 2017, 163, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Bokhove, M.; Jimenez, P.N.; Quax, W.J.; Dijkstra, B.W. The quorum quenching N-acyl homoserine lactone acylase PvdQ is an Ntn-hydrolase with an unusual substrate-binding pocket. Proc. Natl. Acad. Sci. USA 2010, 107, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.N.; Chen, C.J.; Liao, C.T.; Lee, C.Y. A probable aculeacin A acylase from the Ralstonia solanacearum GMI1000 is N-acyl-homoserine lactone acylase with quorum quenching activity. BMC Microbiol. 2009, 9, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pundle, A.; SivaRaman, H. Bacillus sphaericus penicillin V acylase: Purification, substrate specificity, and active-site characterization. Curr. Microbiol. 1997, 34, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Rathinaswamy, P.; Pundle, A.V.; Prabhune, A.A.; Sivaraman, H.; Brannigan, J.A.; Dodson, G.G.; Suresh, C.G. Cloning, purification, crystallization and preliminary structural studies of penicillin V acylase from Bacillus subtilis. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 680–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.X.; Lee, B.H. Bile salt hydrolases: Structure and function, substrate preference, and inhibitor development. Protein Sci. 2018, 27, 1742–1754. [Google Scholar] [CrossRef]
- Kumar, R.S.; Brannigan, J.A.; Prabhune, A.A.; Pundle, A.V.; Dodson, G.G.; Dodson, E.J.; Suresh, C.G. Structural and functional analysis of a conjugated bile salt hydrolase from Bifidobacterium longum reveals an evolutionary relationship with penicillin V acylase. J. Biol. Chem. 2006, 281, 32516–32525. [Google Scholar] [CrossRef] [Green Version]
- Chand, D.; Panigrahi, P.; Varshney, N.; Ramasamy, S.; Suresh, C.G. Structure and function of a highly active Bile Salt Hydrolase (BSH) from Enterococcus faecalis and post-translational processing of BSH enzymes. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2018, 1866, 507–518. [Google Scholar] [CrossRef]
- Koreishi, M.; Tani, K.; Ise, Y.; Imanaka, H.; Imamura, K.; Nakanishi, K. Enzymatic synthesis of β-lactam antibiotics and N-fatty-acylated amino compounds by the acyl-transfer reaction catalyzed by penicillin V acylase from Streptomyces mobaraensis. Biosci. Biotechnol. Biochem. 2007, 71, 1582–1586. [Google Scholar] [CrossRef] [Green Version]
- Isogai, Y.; Nakayama, K. Alteration of substrate selection of antibiotic acylase from β-lactam to echinocandin. Protein Eng. Des. Sel. 2016, 29, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.; Nadal-Jimenez, P.; Reis, C.R.; Muntendam, R.; Bokhove, M.; Melillo, E.; Dijkstra, B.W.; Cool, R.H.; Quax, W.J. Reducing virulence of the human pathogen Burkholderia by altering the substrate specificity of the quorum quenching acylase PvdQ. Proc. Natl. Acad. Sci. USA 2014, 111, 1568–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duggleby, H.J.; Tolley, S.P.; Hill, C.P.; Dodson, E.J.; Dodson, G.; Moody, P.C. Penicillin acylase has a single-amino-acid catalytic centre. Nature 1995, 373, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Brannigan, J.A.; Dodson, G.; Duggleby, H.J.; Moody, P.C.; Smith, J.L.; Tomchick, D.R.; Murzin, A.G. A protein catalytic framework with an N-terminal nucleophile is capable of self-activation. Nature 1995, 378, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, Y. Active site residues of cephalosporin acylase are critical not only for enzymatic catalysis but also for post-translational modification. J. Biol. Chem. 2001, 276, 48376–48381. [Google Scholar] [CrossRef] [Green Version]
- McVey, C.E.; Walsh, M.A.; Dodson, G.G.; Wilson, K.S.; Brannigan, J.A. Crystal structures of penicillin acylase enzyme-substrate complexes: Structural insights into the catalytic mechanism. J. Mol. Biol. 2001, 313, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, J.; Jiang, W.; Mao, X.; Zhao, G.; Wang, E. In vivo post-translational processing and subunit reconstitution of cephalosporin acylase from Pseudomonas sp. 130. Eur. J. Biochem. 1999, 262, 713–719. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, L.; Kasche, V.; Lummer, K.; Lewis, R.J.; Murshudov, G.N.; Verma, C.S.; Dodson, G.G.; Wilson, K.S. Structure of a slow processing precursor penicillin acylase from Escherichia coli reveals the linker peptide blocking the active-site cleft. J. Mol. Biol. 2000, 302, 887–898. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; The John Innes Foundation: Norwich, UK, 2000; ISBN 9780708406236. [Google Scholar]
- Clevenger, K.D.; Wu, R.; Er, J.A.V.; Liu, D.; Fast, W. Rational design of a transition state analogue with picomolar affinity for Pseudomonas aeruginosa PvdQ, a siderophore biosynthetic enzyme. ACS Chem. Biol. 2013, 8, 2192–2200. [Google Scholar] [CrossRef]
- Švedas, V.J.K.; Galaev, I.J.; Borisov, I.L.; Berezin, I.V. The interaction of amino acids with o-phthaldialdehyde: A kinetic study and spectrophotometric assay of the reaction product. Anal. Biochem. 1980, 101, 188–195. [Google Scholar] [CrossRef]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torabi, S.F.; Khajeh, K.; Ghasempur, S.; Ghaemi, N.; Siadat, S.O. Covalent attachment of cholesterol oxidase and horseradish peroxidase on perlite through silanization: Activity, stability and co-immobilization. J. Biotechnol. 2007, 131, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Saitou, N. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 9780199881222. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X. Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Km (mM) | kcat (s−1) | kcat/Km (mM−1 s−1) |
|---|---|---|---|
| C8-HSL | 1.19 ± 0.16 | 22.9 ± 1.2 | 19.2 |
| 3-oxo-C8-HSL | 0.93 ± 0.14 | 7.34 ± 0.42 | 7.9 |
| C10-HSL | 0.25 ± 0.04 | 21.9 ± 1.1 | 87.6 |
| 3-oxo-C10-HSL | 0.40 ± 0.07 | 14.8 ± 1.0 | 37.0 |
| C12-HSL | 0.13 ± 0.02 | 19.3 ± 1.1 | 148.5 |
| 3-oxo-C12-HSL | 0.22 ± 0.03 | 20.4 ± 1.3 | 92.7 |
| C14-HSL | 0.039 ± 0.007 | 3.80 ± 0.25 | 97.4 |
| 3-oxo-C14-HSL | 0.137 ± 0.032 | 10.8 ± 1.9 | 78.8 |
| Substrate | Km (mM) | kcat (s−1) | kcat/Km (mM−1 s−1) |
|---|---|---|---|
| C8-HSL | 2.70 ± 0.76 | 24.7 ± 3.6 | 9.2 |
| 3-oxo-C8-HSL | 0.45 ± 0.14 | 2.05 ± 0.20 | 4.6 |
| C10-HSL | 0.19 ± 0.03 | 13.6 ± 0.8 | 71.6 |
| 3-oxo-C10-HSL | 0.47 ± 0.10 | 4.45 ± 0.39 | 9.5 |
| C12-HSL | 0.10 ± 0.02 | 8.15 ± 0.56 | 81.5 |
| 3-oxo-C12-HSL | 0.17 ± 0.03 | 7.95 ± 0.57 | 46.8 |
| C14-HSL | 0.013 ± 0.003 | 1.19 ± 0.11 | 91.5 |
| 3-oxo-C14-HSL | 0.023 ± 0.006 | 1.79 ± 0.23 | 77.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velasco-Bucheli, R.; Hormigo, D.; Fernández-Lucas, J.; Torres-Ayuso, P.; Alfaro-Ureña, Y.; Saborido, A.I.; Serrano-Aguirre, L.; García, J.L.; Ramón, F.; Acebal, C.; et al. Penicillin Acylase from Streptomyces lavendulae and Aculeacin A Acylase from Actinoplanes utahensis: Two Versatile Enzymes as Useful Tools for Quorum Quenching Processes. Catalysts 2020, 10, 730. https://doi.org/10.3390/catal10070730
Velasco-Bucheli R, Hormigo D, Fernández-Lucas J, Torres-Ayuso P, Alfaro-Ureña Y, Saborido AI, Serrano-Aguirre L, García JL, Ramón F, Acebal C, et al. Penicillin Acylase from Streptomyces lavendulae and Aculeacin A Acylase from Actinoplanes utahensis: Two Versatile Enzymes as Useful Tools for Quorum Quenching Processes. Catalysts. 2020; 10(7):730. https://doi.org/10.3390/catal10070730
Chicago/Turabian StyleVelasco-Bucheli, Rodrigo, Daniel Hormigo, Jesús Fernández-Lucas, Pedro Torres-Ayuso, Yohana Alfaro-Ureña, Ana I. Saborido, Lara Serrano-Aguirre, José L. García, Fernando Ramón, Carmen Acebal, and et al. 2020. "Penicillin Acylase from Streptomyces lavendulae and Aculeacin A Acylase from Actinoplanes utahensis: Two Versatile Enzymes as Useful Tools for Quorum Quenching Processes" Catalysts 10, no. 7: 730. https://doi.org/10.3390/catal10070730
APA StyleVelasco-Bucheli, R., Hormigo, D., Fernández-Lucas, J., Torres-Ayuso, P., Alfaro-Ureña, Y., Saborido, A. I., Serrano-Aguirre, L., García, J. L., Ramón, F., Acebal, C., Santos, A., Arroyo, M., & de la Mata, I. (2020). Penicillin Acylase from Streptomyces lavendulae and Aculeacin A Acylase from Actinoplanes utahensis: Two Versatile Enzymes as Useful Tools for Quorum Quenching Processes. Catalysts, 10(7), 730. https://doi.org/10.3390/catal10070730