
Production of Ethylene Glycol from Glycerol Using an In Vitro Enzymatic Cascade

Abstract
:1. Introduction
2. Results and Discussion
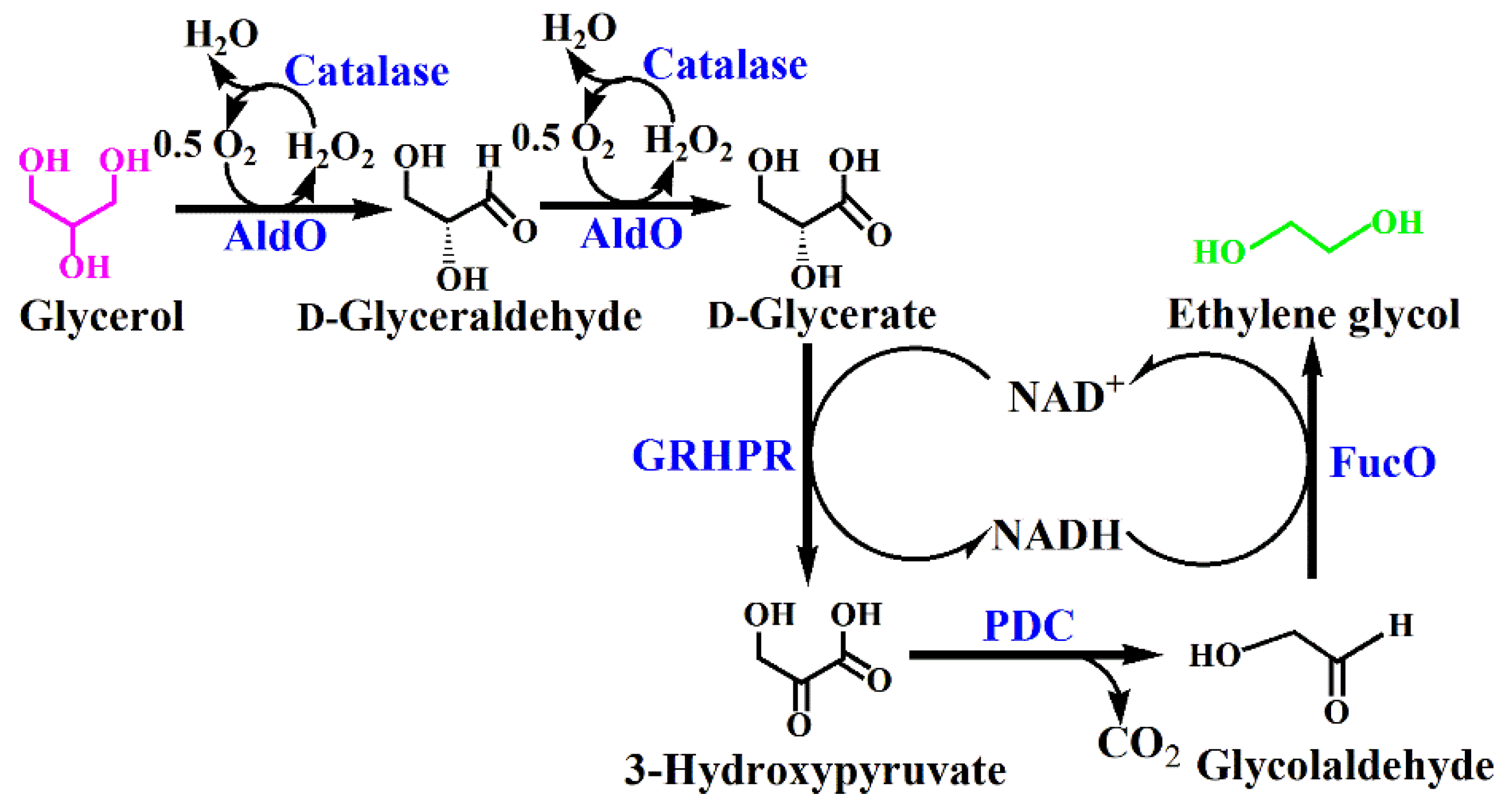
2.1. Design of the In Vitro Biosystem for Ethylene Glycol Production
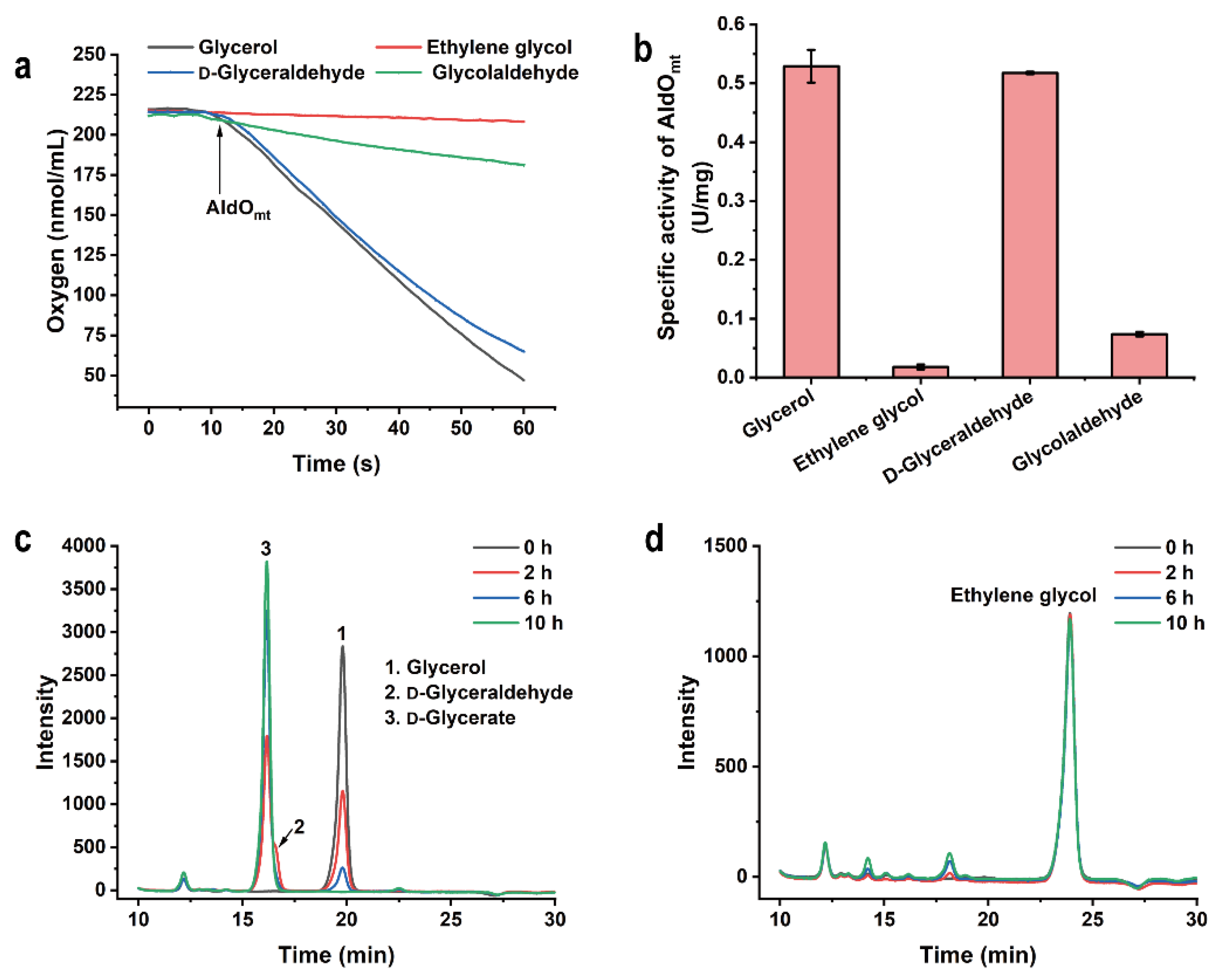
2.2. Substrate Specificity of AldOmt
2.3. 3-Hydroxypyruvate Decarboxylation Catalyzed by PDC
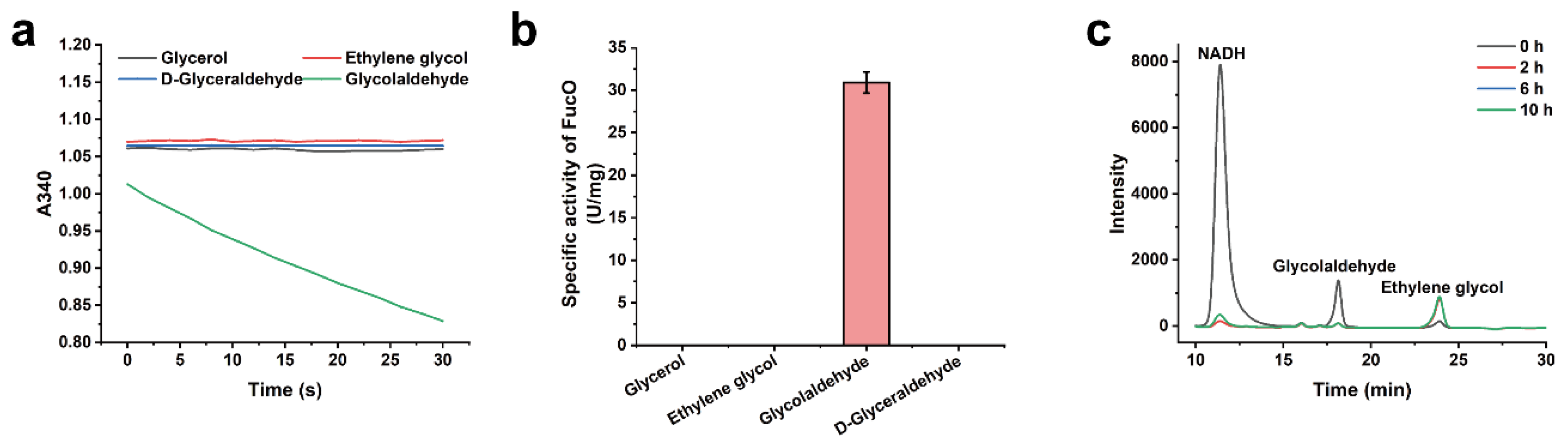
2.4. Substrate Specificity of FucO
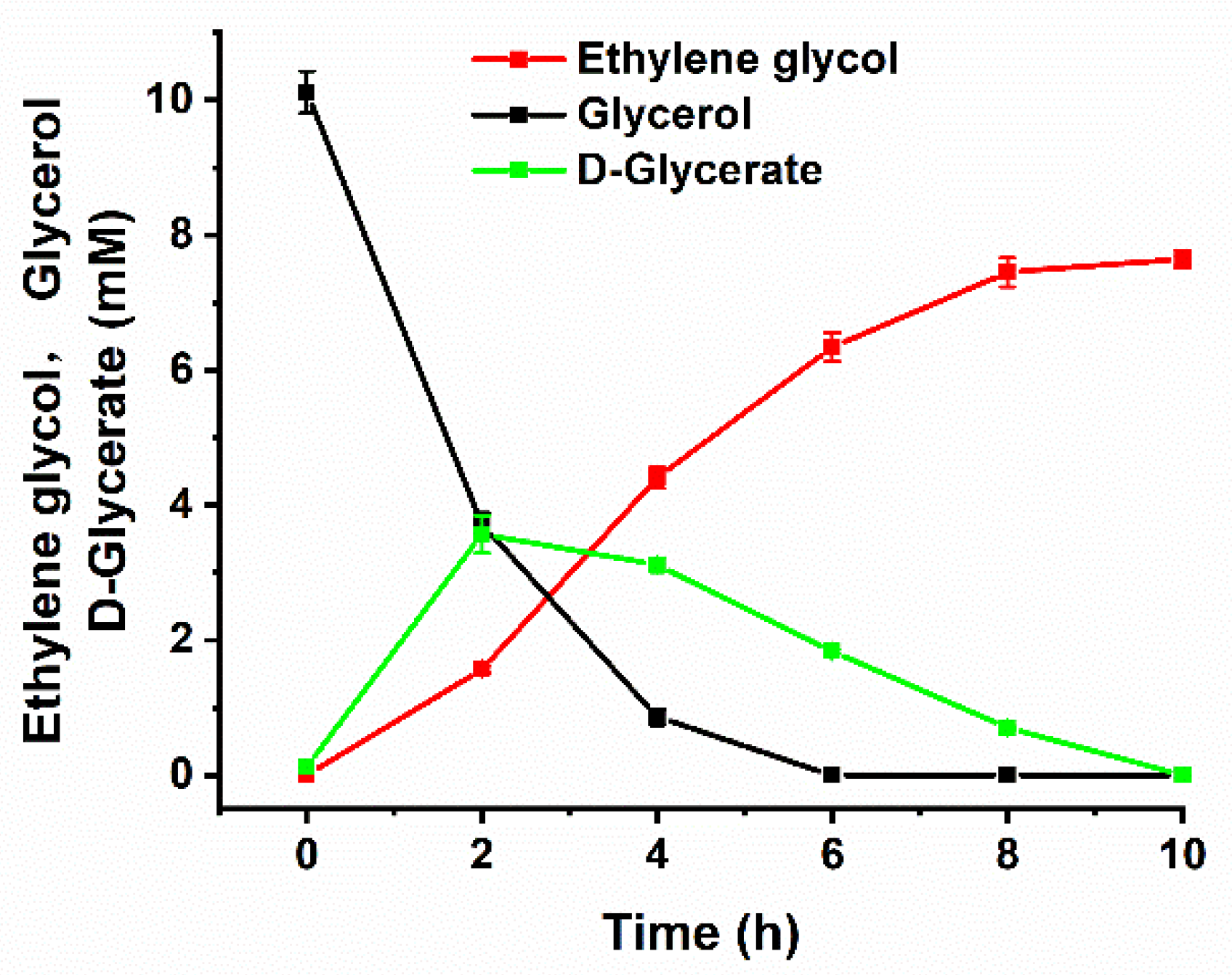
2.5. Production of Ethylene Glycol from Glycerol In Vitro
3. Materials and Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mahmood, W.M.A.; Theodoropoulos, C.; Gonzalez-Miquel, M. Enhanced microalgal lipid extraction using bio-based solvents for sustainable biofuel production. Green Chem. 2017, 19, 5723–5733. [Google Scholar] [CrossRef]
- Hill, J.; Nelson, E.; Tilman, D.; Polasky, S.; Tiffany, D. Environmental, economic, and energetic costs and benefits of biodiesel and ethanol biofuels. Proc. Natl. Acad. Sci. USA 2006, 103, 11206–11210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heeres, A.S.; Picone, C.S.; van der Wielen, L.A.; Cunha, R.L.; Cuellar, M.C. Microbial advanced biofuels production: Over-coming emulsification challenges for large-scale operation. Trends Biotechnol. 2014, 32, 221–229. [Google Scholar] [CrossRef]
- Tan, H.W.; Abdul Aziz, A.; Aroua, M. Glycerol production and its applications as a raw material: A review. Renew. Sustain. Energy Rev. 2013, 27, 118–127. [Google Scholar] [CrossRef]
- Lu, X.; Vora, H.; Khosla, C. Overproduction of free fatty acids in E. coli: Implications for biodiesel production. Metab. Eng. 2008, 10, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Zhao, Y.; Ma, X.; Gong, J. Ethylene glycol: Properties, synthesis, and applications. Chem. Soc. Rev. 2012, 41, 4218–4244. [Google Scholar] [CrossRef]
- Kawabe, K. Development of highly selective process for mono-ethylene glycol production from ethylene oxide via ethylene carbonate using phosphonium salt catalyst. Catal. Surv. Asia 2010, 14, 111–115. [Google Scholar] [CrossRef]
- Li, Y.; Yue, B.; Yan, S.; Yang, W.; Xie, Z.; Chen, Q.; He, H. Preparation of ethylene glycol via catalytic hydration with highly efficient supported niobia catalyst. Catal. Lett. 2004, 95, 163–166. [Google Scholar] [CrossRef]
- Salusjärvi, L.; Havukainen, S.; Koivistoinen, O.; Toivari, M. Biotechnological production of glycolic acid and ethylene glycol: Current state and perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 2525–2535. [Google Scholar] [CrossRef] [Green Version]
- Ueda, N.; Nakagawa, Y.; Tomishige, K. Conversion of glycerol to ethylene glycol over Pt-modified Ni catalyst. Chem. Lett. 2010, 39, 506–507. [Google Scholar] [CrossRef]
- Yin, A.-Y.; Guo, X.-Y.; Dai, W.-L.; Fan, K.-N. The synthesis of propylene glycol and ethylene glycol from glycerol using Raney Ni as a versatile catalyst. Green Chem. 2009, 11, 1514–1516. [Google Scholar] [CrossRef]
- Liu, H.; Ramos, K.R.M.; Valdehuesa, K.N.G.; Nisola, G.M.; Lee, W.-K.; Chung, W.-J. Biosynthesis of ethylene glycol in Escherichia coli. Appl. Microbiol. Biotechnol. 2012, 97, 3409–3417. [Google Scholar] [CrossRef]
- Cam, Y.; Alkim, C.; Trichez, D.; Trebosc, V.; Vax, A.; Bartolo, F.; Besse, P.; François, J.-M.; Walther, T. Engineering of a synthetic metabolic pathway for the assimilation of (d)-Xylose into value-added chemicals. ACS Synth. Biol. 2015, 5, 607–618. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, J.; Wu, Y.; Liu, D. Metabolic engineering of Corynebacterium glutamicum for the de novo production of ethylene glycol from glucose. Metab. Eng. 2016, 33, 12–18. [Google Scholar] [CrossRef]
- Rollin, J.A.; Tam, T.K.; Zhang, Y.-H.P. New biotechnology paradigm: Cell-free biosystems for biomanufacturing. Green Chem. 2013, 15, 1708–1719. [Google Scholar] [CrossRef]
- Shi, T.; Han, P.; You, C.; Zhang, Y.P.J. An in vitro synthetic biology platform for emerging industrial biomanufacturing: Bot-tom-up pathway design. Synth. Syst. Biotechnol. 2018, 3, 186–195. [Google Scholar] [CrossRef]
- Gao, C.; Li, Z.; Zhang, L.; Wang, C.; Li, K.; Ma, C.; Xu, P. An artificial enzymatic reaction cascade for a cell-free bio-system based on glycerol. Green Chem. 2015, 17, 804–807. [Google Scholar] [CrossRef]
- Li, Z.; Yan, J.; Sun, J.; Xu, P.; Ma, C.; Gao, C. Production of value-added chemicals from glycerol using in vitro enzymatic cascades. Commun. Chem. 2018, 1, 1–7. [Google Scholar] [CrossRef]
- Jia, X.; Kelly, M.R.; Han, Y. Simultaneous biosynthesis of (R)-acetoin and ethylene glycol from D-xylose through in vitro met-abolic engineering. Metab. Eng. Commun. 2018, 7, e00074. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, M.; Jiang, T.; Sheng, B.; Ma, C.; Xu, P.; Gao, C. Enzymatic cascades for efficient biotransformation of racemic lactate derived from corn steep water. ACS Sustain. Chem. Eng. 2017, 5, 3456–3464. [Google Scholar] [CrossRef]
- Heuts, D.P.H.M.; Van Hellemond, E.W.; Janssen, D.B.; Fraaije, M.W. Discovery, characterization, and kinetic analysis of an alditol oxidase from Streptomyces coelicolor. J. Biol. Chem. 2007, 282, 20283–20291. [Google Scholar] [CrossRef] [Green Version]
- Forneris, F.; Heuts, D.P.H.M.; Delvecchio, M.; Rovida, S.; Fraaije, M.W.; Mattevi, A. Structural analysis of the catalytic mechanism and stereoselectivity in Streptomyces coelicolor alditol oxidase. Biochemistry 2008, 47, 978–985. [Google Scholar] [CrossRef] [Green Version]
- Gerstenbruch, S.; Wulf, H.; Mußmann, N.; O’Connell, T.; Maurer, K.-H.; Bornscheuer, U.T.; Mussmann, N. Asymmetric synthesis of d-glyceric acid by an alditol oxidase and directed evolution for enhanced oxidative activity towards glycerol. Appl. Microbiol. Biotechnol. 2012, 96, 1243–1252. [Google Scholar] [CrossRef]
- Sheng, B.; Xu, J.; Ge, Y.; Zhang, S.; Wang, D.; Gao, C.; Ma, C.; Xu, P. Enzymatic resolution by a d-lactate oxidase catalyzed reaction for (S)-2-hydroxycarboxylic acids. ChemCatChem 2016, 8, 2630–2633. [Google Scholar] [CrossRef]
- Montella, C.; Bellsolell, L.; Pérez-Luque, R.; Badía, J.; Baldoma, L.; Coll, M.; Aguilar, J. Crystal structure of an iron-dependent group III dehydrogenase that interconverts l-lactaldehyde and l-1,2-propanediol in Escherichia coli. J. Bacteriol. 2005, 187, 4957–4966. [Google Scholar] [CrossRef] [Green Version]
- Iding, H.; Siegert, P.; Mesch, K.; Pohl, M. Application of α-keto acid decarboxylases in biotransformations. Biochim. Biophys. Acta Protein Struct. Mol. Enzym. 1998, 1385, 307–322. [Google Scholar] [CrossRef]
- Wang, X.; Miller, E.N.; Yomano, L.P.; Zhang, X.; Shanmugam, K.T.; Ingram, L.O. Increased furfural tolerance due to overex-pression of NADH-dependent oxidoreductase FucO in Escherichia coli strains engineered for the production of ethanol and lactate. Appl. Environ. Microbiol. 2011, 77, 5132–5140. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Lin, E.C. L-1,2-propanediol exits more rapidly than l-lactaldehyde from Escherichia coli. J. Bacteriol. 1989, 171, 862–867. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xian, M.; Feng, X.; Liu, M.; Zhao, G. Biosynthesis of ethylene glycol from d-xylose in recombinant Escherichia coli. Bioengineered 2018, 9, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Zhang, Y.; Xiong, W.; Ding, H.; He, B. Hydrogenolysis of glycerol to 1,2-propanediol and ethylene glycol over Ru-Co/ZrO2 catalysts. Catalysts 2016, 6, 51. [Google Scholar] [CrossRef] [Green Version]
- Pereira, B.; Zhang, H.; De Mey, M.; Lim, C.G.; Li, Z.; Stephanopoulos, G. Engineering a novel biosynthetic pathway in Escherichia coli for production of renewable ethylene glycol. Biotechnol. Bioeng. 2016, 113, 376–383. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Wang, Y.; Gu, J.; Lu, X.; Liao, X.; Shi, J.; Kim, C.H.; Lye, G.; Baganz, F.; et al. Ethylene glycol and glycolic acid production from xylonic acid by Enterobacter cloacae. Microb. Cell Fact. 2020, 19, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Pereira, B.; Li, Z.-J.; De Mey, M.; Lim, C.G.; Zhang, H.; Hoeltgen, C.; Stephanopoulos, G. Efficient utilization of pentoses for bioproduction of the renewable two-carbon compounds ethylene glycol and glycolate. Metab. Eng. 2016, 34, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Chae, T.U.; Choi, S.Y.; Ryu, J.Y.; Lee, S.Y. Production of ethylene glycol from xylose by metabolically engineered Escherichia coli. AICHE J. 2018, 64, 4193–4200. [Google Scholar] [CrossRef]
- Cabulong, R.B.; Chung, W.-J.; Ramos, K.R.M.; Nisola, G.M.; Lee, W.-K.; Lee, C.R.; Chung, W.-J. Enhanced yield of ethylene glycol production from d-xylose by pathway optimization in Escherichia coli. Enzym. Microb. Technol. 2017, 97, 11–20. [Google Scholar] [CrossRef]
- You, C.; Shi, T.; Li, Y.; Han, P.; Zhou, X.; Zhang, Y.-H.P. An in vitro synthetic biology platform for the industrial biomanufacturing of myo-inositol from starch. Biotechnol. Bioeng. 2017, 114, 1855–1864. [Google Scholar] [CrossRef]
- Rollin, J.A.; Del Martin del Campo, J.; Myung, S.; Sun, F.; You, C.; Bakovic, A.; Castro, R.; Chandrayan, S.K.; Wu, C.-H.; Adams, M.W.; et al. High-yield hydrogen production from biomass by in vitro metabolic engineering: Mixed sugars coutilization and kinetic modeling. Proc. Natl. Acad. Sci. USA 2015, 112, 4964–4969. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Enzymes | Substrate | Ethylene Glycol Yield (g/g) | Reference |
|---|---|---|---|
| E. coli BW25113(DE3) ΔxylAΔaldA/pACYC-xdh-yqhD | d-Xylose | 0.29 | [12] |
| E. coli MG1655 ΔxylB/pEXT20-khkC-aldoB | d-Xylose | 0.19 | [13] |
| E. coli BL21(DE3) ΔarcAΔaldA/pETDuet1-yjhH-xdh- xylC/pACYCDuet1-fucO-yjhG | d-Xylose | 0.40 | [29] |
| E. coli MG1655(DE3) ΔendAΔrecA ΔxylBΔaldA/p10_T5-Pc.dte-fucA-fucO-fucK | d-Xylose | 0.35 | [33] |
| E. coli W3110 ΔlacI/pTacxylBC-yqhD-P1-anti-xylB | d-Xylose | 0.36 | [34] |
| E. coli W3110 ΔxylABΔaldAΔyjgBΔyqhD/pKMX & pTrcHis2A-yqhD | d-Xylose | 0.39 | [35] |
| Enterobacter cloacae S1 | Xylonic acid | 0.288 | [32] |
| E. coli MG1655(DE3) ΔendAΔrecAΔxylBΔaraBΔaldA/p5_T7-Pc.dte-rhaB-rhaD-fucO | l-Arabinose | 0.38 | [33] |
| E. coli MG1655(DE3) ΔaldΔAeutBC/p10_T7-serA:317-serB-serC-fucO & pCDFDuet_T7-At.sdc.t+T7-aao | d-Glucose | 0.14 | [31] |
| C. glutamicum PABS1/pEC-P1-GsLdh-PpMdlc-yqhD | d-Glucose | 0.09 | [14] |
| d-Xylose dehydrogenase, xylonolactonase, xylonate dehydratase, 2-keto-3-deoxy-d-xylonate aldolase, lactaldehyde reductase, α-acetolactate synthase, α-acetolactate decarboxylase | d-Xylose | 0.227 | [19] |
| AldOmt, catalase, GRHPR, PDC, FucO | Glycerol | 0.515 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.; Sun, W.; Meng, W.; Yan, J.; Zhang, Y.; Guo, S.; Lü, C.; Ma, C.; Gao, C. Production of Ethylene Glycol from Glycerol Using an In Vitro Enzymatic Cascade. Catalysts 2021, 11, 214. https://doi.org/10.3390/catal11020214
Li K, Sun W, Meng W, Yan J, Zhang Y, Guo S, Lü C, Ma C, Gao C. Production of Ethylene Glycol from Glycerol Using an In Vitro Enzymatic Cascade. Catalysts. 2021; 11(2):214. https://doi.org/10.3390/catal11020214
Chicago/Turabian StyleLi, Kai, Weikang Sun, Wensi Meng, Jinxin Yan, Yipeng Zhang, Shiting Guo, Chuanjuan Lü, Cuiqing Ma, and Chao Gao. 2021. "Production of Ethylene Glycol from Glycerol Using an In Vitro Enzymatic Cascade" Catalysts 11, no. 2: 214. https://doi.org/10.3390/catal11020214
APA StyleLi, K., Sun, W., Meng, W., Yan, J., Zhang, Y., Guo, S., Lü, C., Ma, C., & Gao, C. (2021). Production of Ethylene Glycol from Glycerol Using an In Vitro Enzymatic Cascade. Catalysts, 11(2), 214. https://doi.org/10.3390/catal11020214