Development and Characterization of Two Types of Surface Displayed Levansucrases for Levan Biosynthesis


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Construction of Two Yeast Cell Surface Display Systems
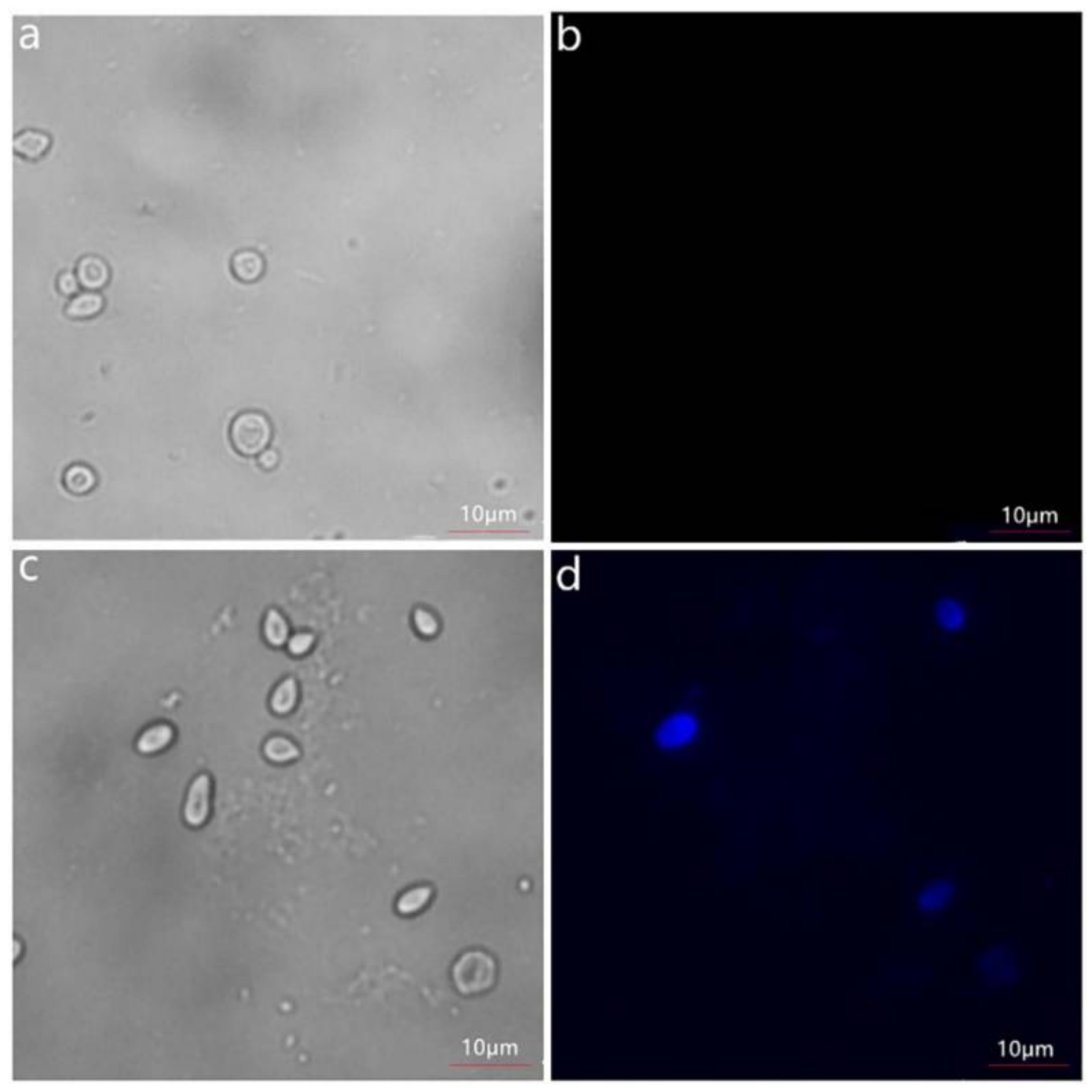
2.2. Verification of the Recombinant Levansucrase Localization on S. cerevisiae EBY100
2.3. Characterization of Levansucrase
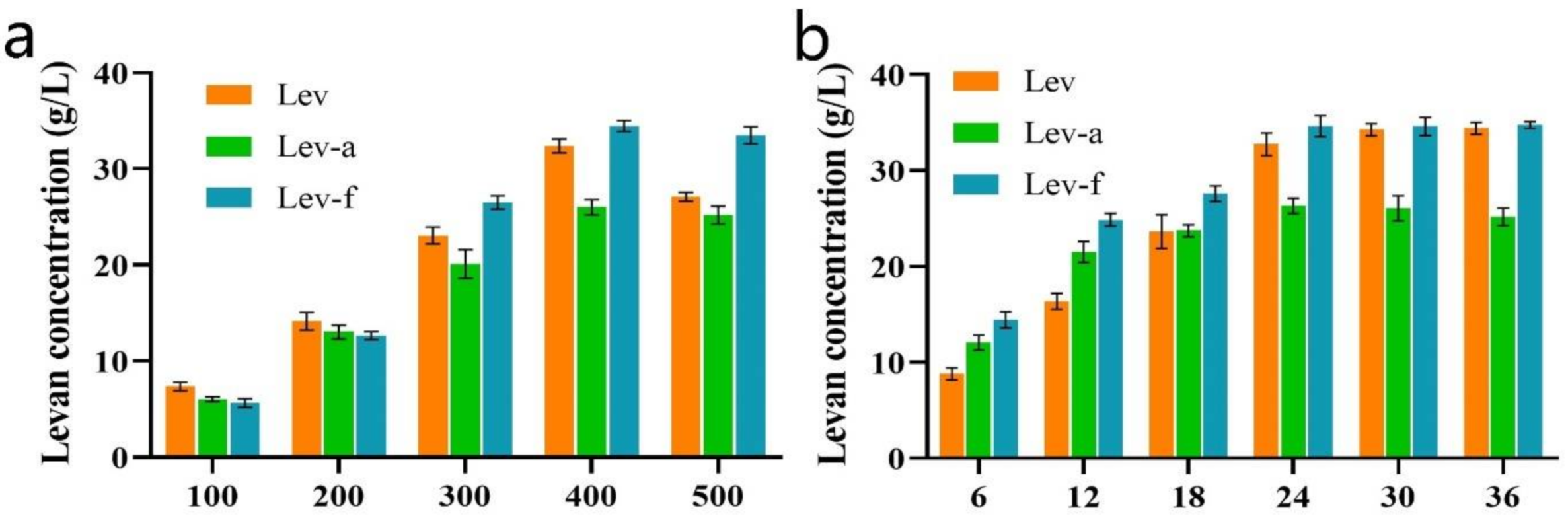
2.4. Levan Biosynthesis
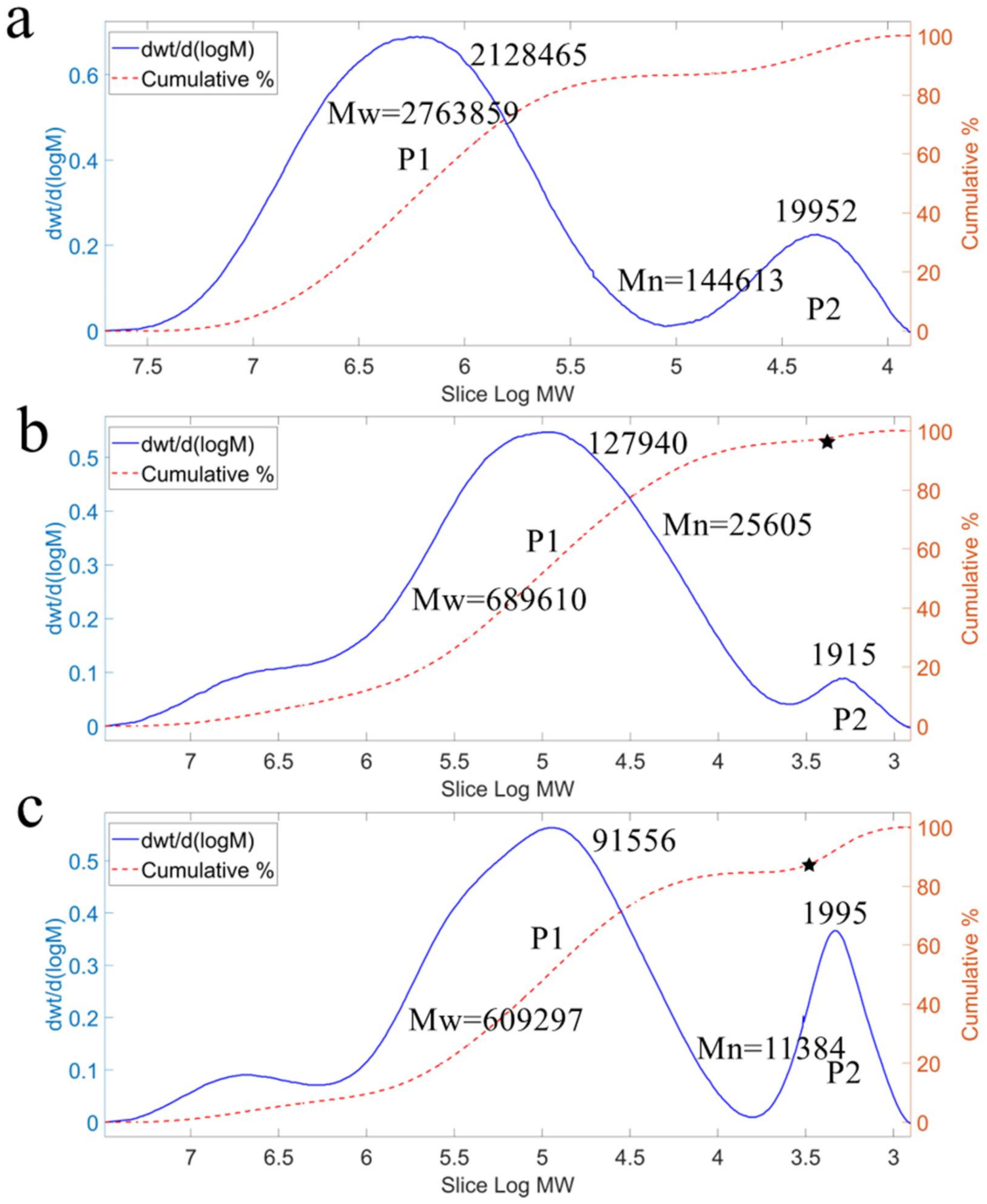
2.5. Polymerization of Levan Synthesized by Different Forms of Levansucrase
3. Materials and Methods
3.1. Strains, Plasmids and Culture Conditions
3.2. Construction of Cell-Surface Display Plasmids
3.3. Yeast Transformation and Surface Display of Levansucrase
3.4. Indirect Yeast Immunofluorescence
3.5. Enzyme Activity Assay of Levansucrase
3.6. Characterization of Surface Displayed Levansucrases
3.7. Levan Synthesis
3.8. Molecular Weight Measurement
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Smeekens, J.C.M.; Ebskamp, M.J.M.; Weisbeek, P.J. Method for Obtaining Transgenic Plants Showing a Modified Fructan Pattern. U.S. Patent US5986173A, 16 November 1999. [Google Scholar]
- Nadzir, M.M.; Nurhayati, R.W.; Idris, F.N.; Nguyen, M.H. Biomedical Applications of Bacterial Exopolysaccharides: A Review. Polymers 2021, 13, 530. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, R.; Reddy, C.H.; Siddartha, G.; Ramaiah, M.J.; Uppuluri, K.B. Review on production, characterization and applications of microbial levan. Carbohydr. Polym. 2015, 120, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yu, S.; Zhang, T.; Jiang, B.; Mu, W. Recent novel applications of levansucrases. Appl. Microbiol. Biotechnol. 2015, 99, 6959–6969. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.H.; Song, K.B.; Park, B.S. Levan production by use of the recombinant levansucrase immobilized on titanium-activated magnetite. Process. Biochem. 2001, 37, 339–343. [Google Scholar] [CrossRef]
- Chambert, R.; Petit-Glatron, M.F. Immobilisation of levansucrase on calcium phosphate gel strongly increases its polymerase activity. Carbohydr. Res. 1993, 244, 129–136. [Google Scholar] [CrossRef]
- Iizuka, M.; Yamaguchi, H.; Ono, S.; Minamiura, N. Production and Isolation of Levan by Use of Levansucrase Immobilized on the Ceramic Support SM-10. Biosci. Biotechnol. Biochem. 1993, 57, 322–324. [Google Scholar] [CrossRef]
- Chiang, C.J.; Wang, J.Y.; Chen, P.T.; Chao, Y.P. Enhanced levan production using chitin-binding domain fused levansucrase immobilized on chitin beads. Appl. Microbiol. Biotechnol. 2009, 82, 445–451. [Google Scholar] [CrossRef]
- Lozani, M.; Hossain, A.S.; Mra, V.; Te Pari, R. Surface Display—An Alternative to Classic Enzyme Immobilization. Catalysts 2019, 9, 728. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.-F.; Lin, S.-C.; Juang, T.-Y.; Liu, Y.-C. Shaking Rate during Production Affects the Activity of Escherichia coli Surface-Displayed Candida antarctica Lipase A. Catalysts 2020, 10, 382. [Google Scholar] [CrossRef]
- Kim, D.; Ku, S. Bacillus Cellulase Molecular Cloning, Expression, and Surface Display on the Outer Membrane of Escherichia coli. Molecules 2018, 23, 503. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Katahira, S.; Ueda, M.; Tanaka, A.; Okada, H.; Morikawa, Y.; Fukuda, H.; Kondo, A. Construction of whole-cell biocatalyst for xylan degradation through cell-surface xylanase display in Saccharomyces cerevisiae. J. Mol. Catal. B Enzym. 2002, 17, 189–195. [Google Scholar] [CrossRef]
- Wang, J.K.; He, B.; Du, W.; Luo, Y.; Yu, Z.; Liu, J.X. Yeast with surface displayed xylanase as a new dual purpose delivery vehicle of xylanase and yeast. Anim. Feed. Sci. Technol. 2015, 208, 44–52. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Y.; Pang, H.; Yuan, S.; Wang, X.; Hu, Z.; Zhou, Q.; He, Y.; Yan, Y.; Xu, L. Codisplay of Rhizopus oryzae and Candida rugosa Lipases for Biodiesel Production. Catalysts 2021, 11, 421. [Google Scholar] [CrossRef]
- Zhang, L.; An, J.; Li, L.; Wang, H.; Liu, D.; Li, N.; Cheng, H.; Deng, Z. Highly Efficient Fructooligosaccharides Production by an Erythritol-Producing Yeast Yarrowia lipolytica Displaying Fructosyltransferase. J. Agric. Food Chem. 2016, 64, 3828–3837. [Google Scholar] [CrossRef]
- Jung, H.C.; Lebeault, J.M.; Pan, J.G. Surface display of Zymomonas mobilis levansucrase by using the ice-nucleation protein of Pseudomonas syringae. Nat. Biotechnol. 1998, 16, 576–580. [Google Scholar] [CrossRef]
- Bo, Y.B.; Lee, S.J.; Mah, J.H. Antipathogenic activity and preservative effect of levan (β-2,6-fructan), a multifunctional polysaccharide. Int. J. Food Sci. Technol. 2014, 49, 238–245. [Google Scholar]
- Benigar, E.; Dogsa, I.; Stopar, D.; Jamnik, A.; Kralj Cigic, I.; Tomsic, M. Structure and dynamics of a polysaccharide matrix: Aqueous solutions of bacterial levan. Langmuir 2014, 30, 4172–4182. [Google Scholar] [CrossRef]
- Hundschell, C.S.; Jakob, F.; Wagemans, A.M. Molecular weight dependent structure of the exopolysaccharide levan. Int. J. Biol. Macromol. 2020, 161, 398–405. [Google Scholar] [CrossRef]
- Liu, C.; Kolida, S.; Charalampopoulos, D.; Rastall, R.A. An evaluation of the prebiotic potential of microbial levans from Erwinia sp. 10119. J. Funct. Foods 2019, 64, 103668. [Google Scholar] [CrossRef]
- Porras-Dominguez, J.R.; Avila-Fernandez, A.; Miranda-Molina, A.; Rodriguez-Alegria, M.E.; Munguia, A.L. Bacillus subtilis 168 levansucrase (SacB) activity affects average levan molecular weight. Carbohydr. Polym. 2015, 132, 338–344. [Google Scholar] [CrossRef]
- Hou, Y.; Huang, F.; Yang, H.; Cong, H.; Zhang, X.; Xie, X.; Yang, H.; Tong, Q.; Luo, N.; Zhu, P. Factors affecting the production and molecular weight of levan in enzymatic synthesis by recombinant Bacillus subtilis levansucrase SacB-T305A. Polym. Int. 2020, 70, 185–192. [Google Scholar] [CrossRef]
- Jang, K.H.; Song, K.B.; Kim, C.H.; Chung, B.H.; Kang, S.A.; Chun, U.H.; Choue, R.W.; Rhee, S.K. Comparison of characteristics of levan produced by different preparations of levansucrase from Zymomonas mobilis. Biotechnol. Lett. 2001, 23, 339–344. [Google Scholar] [CrossRef]
- Santos-Moriano, P.; Monsalve-Ledesma, L.; Ortega-Mu?Oz, M.; Fernandez-Arrojo, L.; Ballesteros, A.O.; Santoyo-Gonzalez, F.; Plou, F.J. Vinyl sulfone-activated silica for efficient covalent immobilization of alkaline unstable enzymes: Application to levansucrase for fructooligosaccharide synthesis. RSC Adv. 2016, 6, 64175–64181. [Google Scholar] [CrossRef]
- Song, K.B.; Joo, H.K.; Rhee, S.K. Nucleotide sequence of levansucrase gene (levU) of Zymomonas mobilis ZM1 (ATCC10988). Biochim. Biophys. Acta 1993, 1173, 320–324. [Google Scholar] [CrossRef]
- Wen, S.; Mao, T.; Yao, D.; Li, T.; Wang, F. Yeast Surface Display of Antheraea pernyi Lysozyme Revealed α-Helical Antibacterial Peptides in Its N-Terminal Domain. J. Agric. Food Chem. 2018, 66, 9138–9146. [Google Scholar] [CrossRef]
- Oner, E.T.; Hernandez, L.; Combie, J. Review of Levan polysaccharide: From a century of past experiences to future prospects. Biotechnol. Adv. 2016, 34, 827–844. [Google Scholar] [CrossRef]
- Goldman, D.; Lavid, N.; Schwartz, A.; Shoham, G.; Danino, D.; Shoham, Y. Two active forms of Zymomonas mobilis levansucrase. An ordered microfibril structure of the enzyme promotes levan polymerization. J. Biol. Chem. 2008, 283, 32209. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, P.M.; Rupp, R.A. Inhibition of Sucrose: Sucrose Fructosyl Transferase by Cations and Ionic Strength. Plant Physiol. 1993, 101, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.; Chen, L.; Mariage, A.; Petit, J.L.; Berardinis, V.D.; Karboune, S. Discovery of new levansucrase enzymes with interesting properties and improved catalytic activity to produce levan and fructooligosaccharides. Catal. Sci. Technol. 2019, 9, 2931–2944. [Google Scholar] [CrossRef]
- Ko, H.; Bae, J.H.; Sung, B.H.; Kim, M.J.; Sohn, J.H. Efficient production of levan using a recombinant yeast Saccharomyces cerevisiae hypersecreting a bacterial levansucrase. J. Ind. Microbiol. Biotechnol. 2019, 46, 1611–1620. [Google Scholar] [CrossRef]
- Xavier, J.R.; Ramana, K.V. Optimization of Levan Production by Cold-Active Bacillus licheniformis ANT 179 and Fructooligosaccharide Synthesis by Its Levansucrase. Appl. Biochem. Biotechnol. 2017, 181, 986–1006. [Google Scholar] [CrossRef]
- Hvels, M.; Kosciow, K.; Kniewel, J.; Jakob, F.; Deppenmeier, U. High yield production of levan-type fructans by Gluconobacter japonicus LMG 1417. Int. J. Biol. Macromol. 2020, 164, 295–303. [Google Scholar] [CrossRef]
- van Hijum, S.A.; Szalowska, E.; van der Maarel, M.J.; Dijkhuizen, L. Biochemical and molecular characterization of a levansucrase from Lactobacillus reuteri. Microbiology 2004, 150, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yu, S.; Zhang, T.; Jiang, B.; Mu, W. Efficient biosynthesis of levan from sucrose by a novel levansucrase from Brenneria goodwinii. Carbohydr. Polym. 2017, 157, 1732–1740. [Google Scholar] [CrossRef]
- Khudair, A.Y.; Salman, J.; Ajah, H.A. Production of levan from locally leuconostoc mesensteroides isolates. J. Pharm. Sci. Res. 2018, 10, 3372–3378. [Google Scholar]
- Gmt, C.; Lima, R.C.; de França, F.P.; Lopes, C.E. Molecular weight and antitumour activity of Zymomonas mobilis levans. Int. J. Biol. Macromol. 2000, 27, 245. [Google Scholar]
- Ortizsoto, M.E.; Rivera, M.; Rudiñopiñera, E.; Olvera, C.; Lópezmunguía, A. Selected mutations in Bacillus subtilis levansucrase semi-conserved regions affecting its biochemical properties. Protein Eng. Des. Sel. 2008, 21, 589. [Google Scholar] [CrossRef]
- Dyatlov, V.A.; Kruppa, I.S.; Mamaeva, S.A.; Kireev , V.V. Change of Polysaccharide Molecular-Weight Distribution and Fraction Homogeneity After Periodate Oxidation. Chem. Nat. Compd. 2014, 50, 973–977. [Google Scholar]
- Marx, S.P.; Winkler, S.; Hartmeier, W. Metabolization of beta-(2,6)-linked fructose-oligosaccharides by different bifidobacteria. FEMS Microbiol. Lett. 2000, 182, 163–169. [Google Scholar]
- Gietz, D.; Jean, A.S.; Woods, R.A.; Schiestl, R.H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Res. 1992, 20, 1425. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Biocatalyst | Yield (g/L) | Time (h) | Productivity 1 (g/L/h) | References |
|---|---|---|---|---|---|
| Z.mobilis | Displayed enzyme | 34 | 24 | 1.42 | This study |
| Z.mobilis | Displayed enzyme | 44 | 216 | 0.20 | [16] |
| Bacillus licheniformis | Purified enzyme | 16.35 | 48 | 0.34 | [32] |
| Gluconobacter japonicus | Recombinant enzyme | 148 | 260 | 0.57 | [33] |
| Lactobacillus reuteri | Recombinant enzyme | 1.4 | 16 | 0.09 | [34] |
| Brenneria goodwinii | Purified enzyme | 185 | 12 | 15.4 | [35] |
| Leuconostoc mesenteroides | Crude enzyme | 14.82 | 24 | 0.62 | [36] |
| Bacillus natto | Immobilized enzyme | 18.75 | 24 | 0.78 | [7] |
| PCR Product | Primer Name | Primer Sequence (5′-3′) | Template | Restriction Enzyme Site |
|---|---|---|---|---|
| levU | levU-F | CGGGATCCCGATGTTGAATAAAGCAGGC | Z.mobilis genome | BamH I |
| levU-R | CCGCCTCGAGGTTTATTCAATAAAGACAGG | Xho I | ||
| α-factor | αF1 | CGGGATCCCGATGAGATTTCCTTCAATTTTTTAC | EBY100 genome | BamH I |
| αR1 | GCCTGCTTTATTCAACATAGCTTCAGCCTC | Overlapping | ||
| levU2 | levU-F2 | GAGAGGCTGAAGCTATGTTGAATAAAGCAGGC | levU | Overlapping |
| levU-R2 | GGGGGCCCCCTTTATTCAATAAAGACAGGGC | Apa I | ||
| α-factor-levU | αF1 | CGGGATCCCGATGAGATTTCCTTCAATTTTTTAC | α-factor/levU2 | BamH I |
| levU-R2 | GGGGGCCCCCTTTATTCAATAAAGACAGGGC | Apa I | ||
| FLO1 | LF-F | GCCTCGAGGTATCCTAGCAATGGAACTTCTGTG | EBY100 genome | Xho I |
| LF-R | CGGCTAGCCGTTAAATAATTGCCAGCAATAAG | Nhe I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shang, H.; Yang, D.; Qiao, D.; Xu, H.; Cao, Y. Development and Characterization of Two Types of Surface Displayed Levansucrases for Levan Biosynthesis. Catalysts 2021, 11, 757. https://doi.org/10.3390/catal11070757
Shang H, Yang D, Qiao D, Xu H, Cao Y. Development and Characterization of Two Types of Surface Displayed Levansucrases for Levan Biosynthesis. Catalysts. 2021; 11(7):757. https://doi.org/10.3390/catal11070757
Chicago/Turabian StyleShang, Huiyi, Danni Yang, Dairong Qiao, Hui Xu, and Yi Cao. 2021. "Development and Characterization of Two Types of Surface Displayed Levansucrases for Levan Biosynthesis" Catalysts 11, no. 7: 757. https://doi.org/10.3390/catal11070757
APA StyleShang, H., Yang, D., Qiao, D., Xu, H., & Cao, Y. (2021). Development and Characterization of Two Types of Surface Displayed Levansucrases for Levan Biosynthesis. Catalysts, 11(7), 757. https://doi.org/10.3390/catal11070757