
Biological Hydrogen Production by Dark Fermentation in a Stirred Tank Reactor and Its Correlation with the pH Time Evolution


 ,
, 
Abstract
:1. Introduction
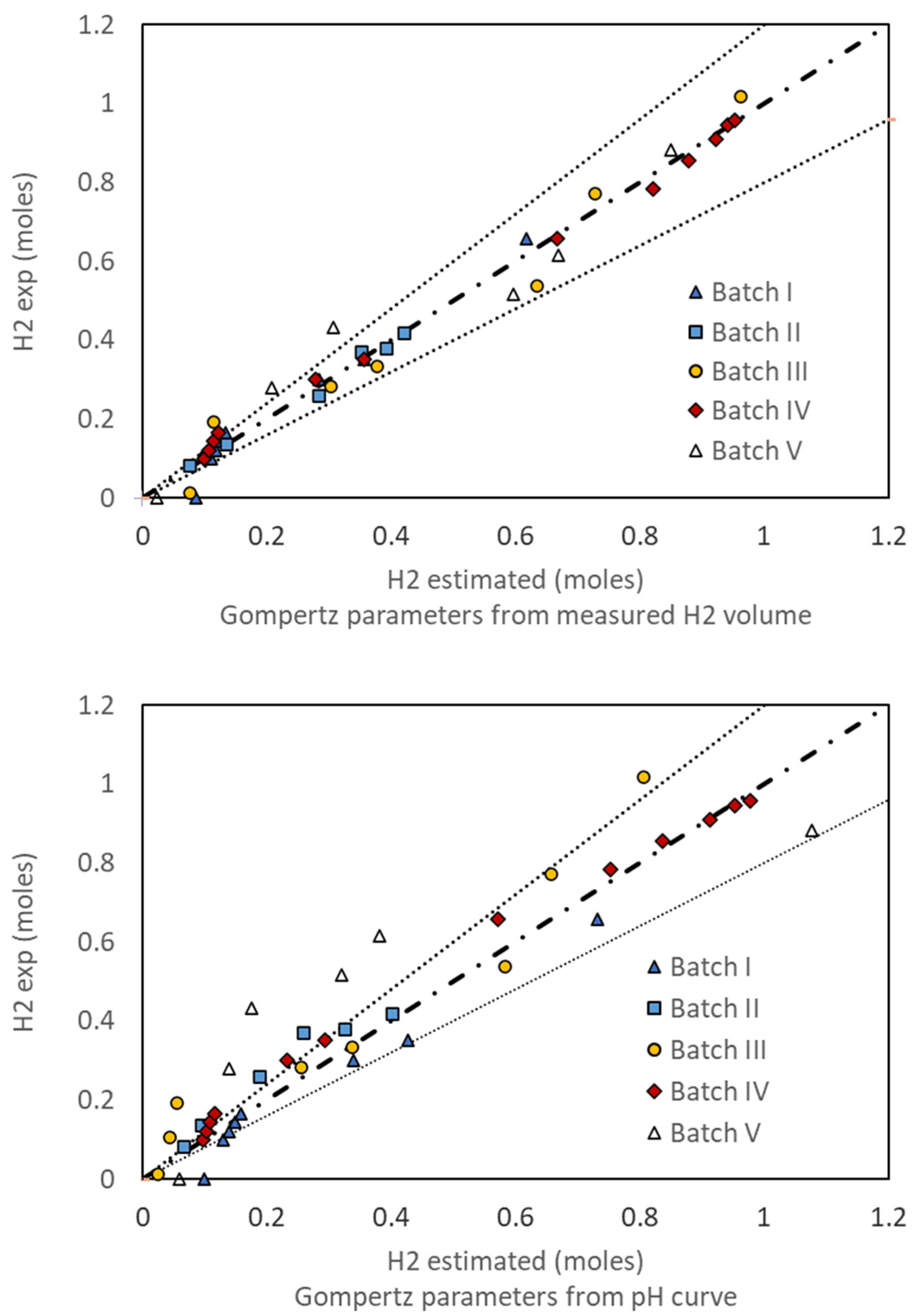
2. Results and Discussion
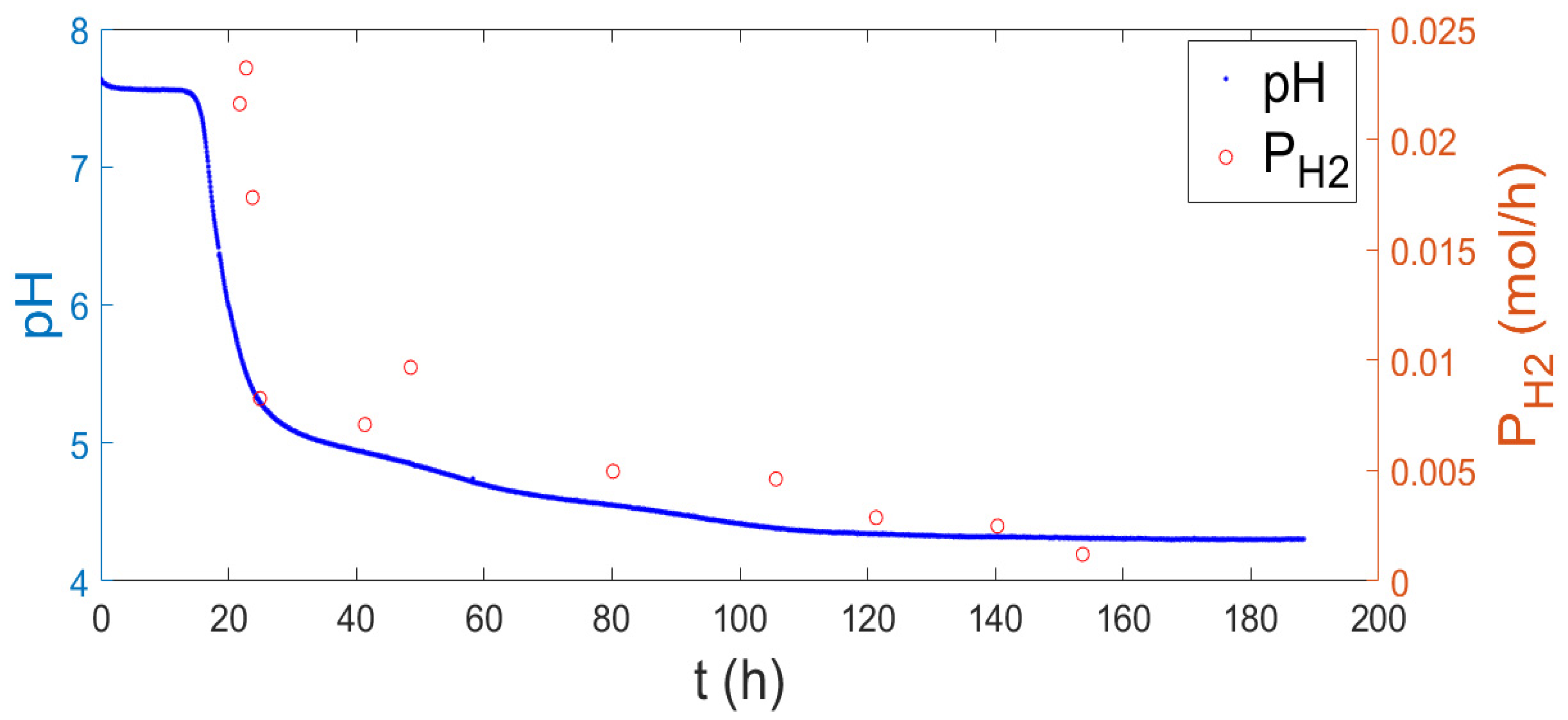
2.1. Hydrogen Measure and Production
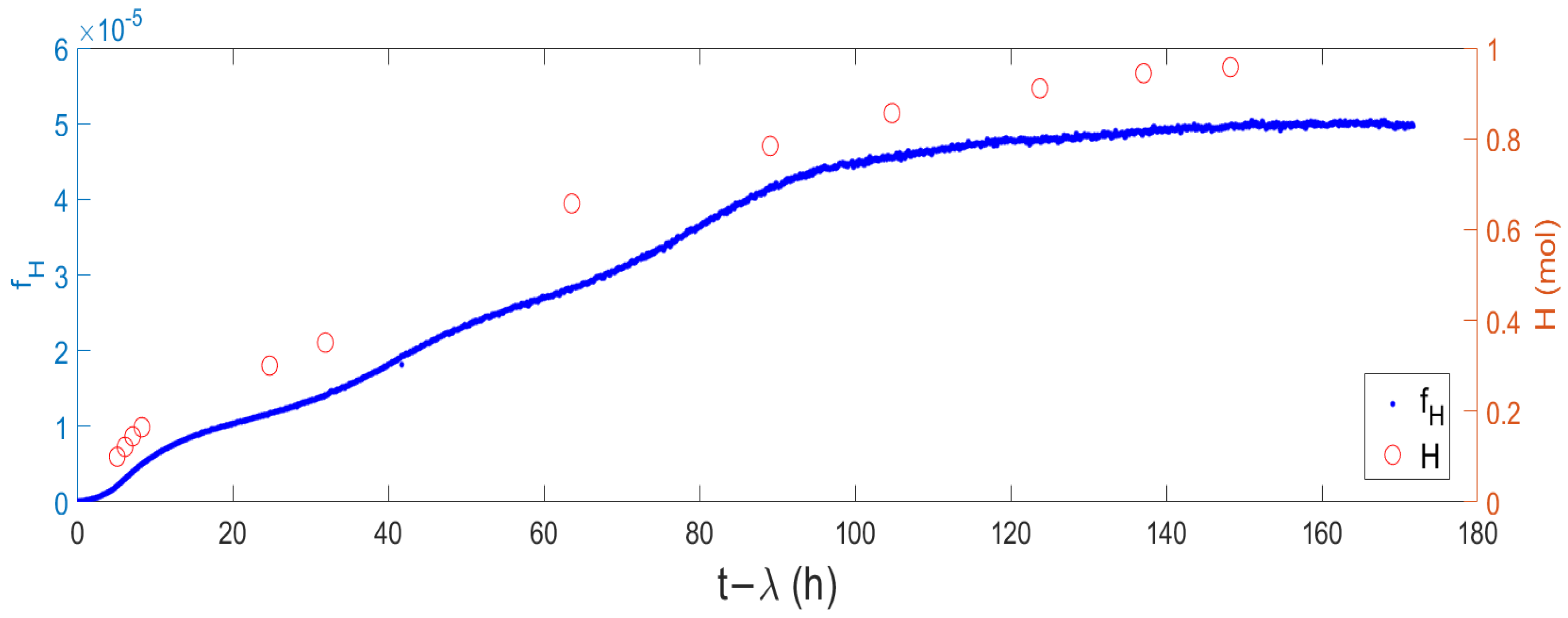
2.2. Relation between the Evolved Hydrogen and the pH Curve
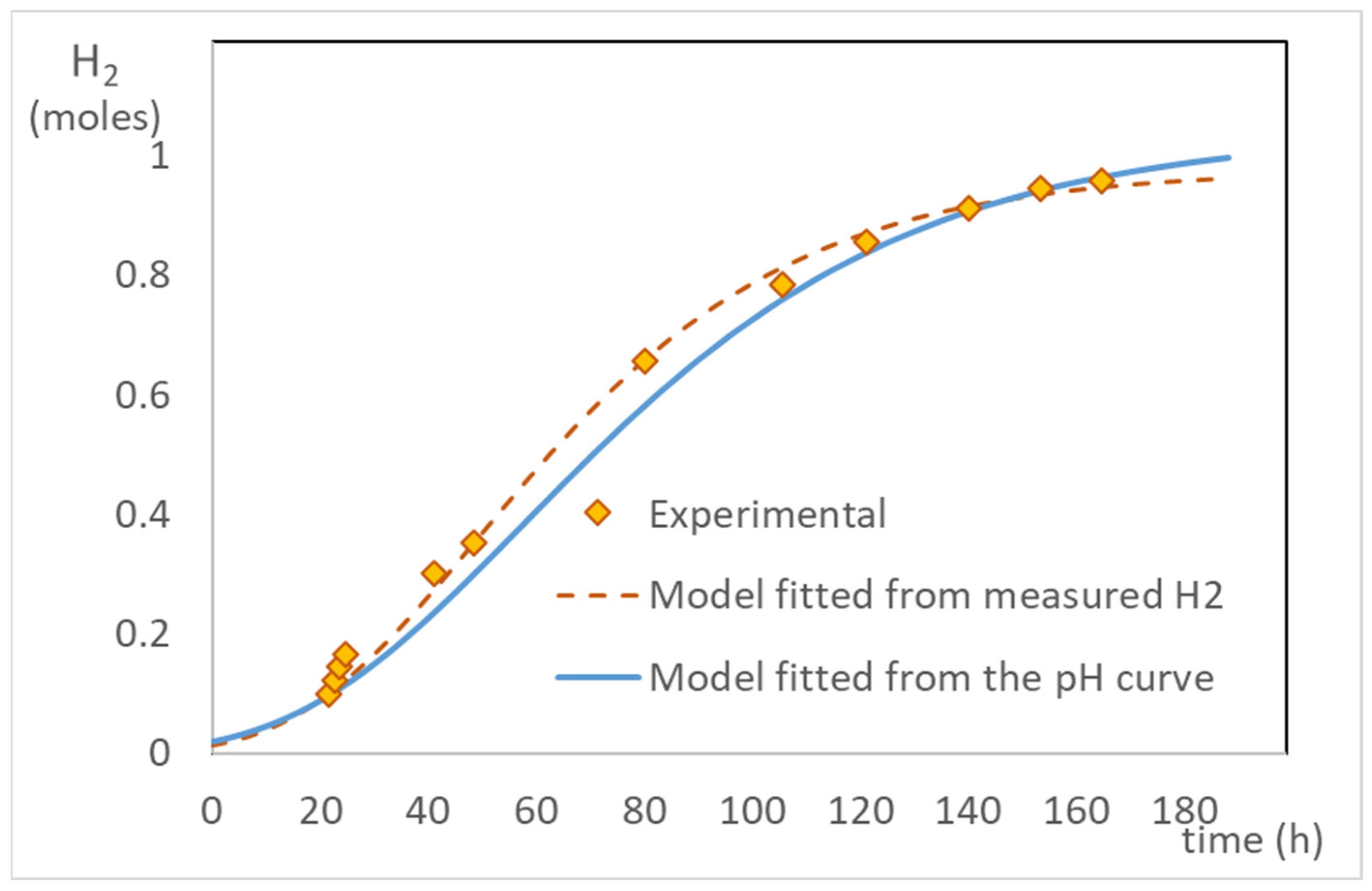
2.3. Correlation between pH Evolution and Hydrogen Production
2.4. Lag Time Calculation
2.5. A Measure of Hydrogen Production Efficiency
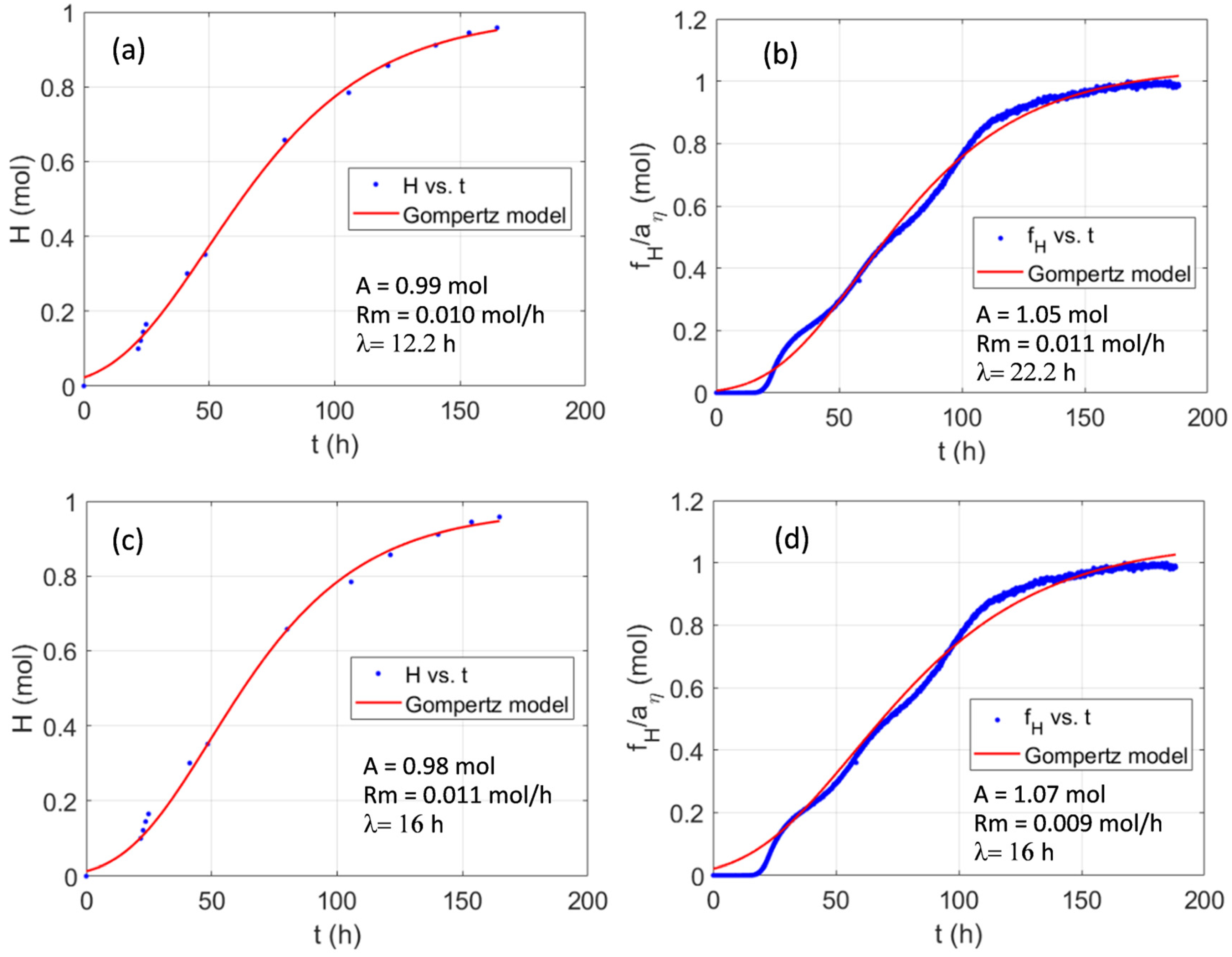
2.6. Kinetic Parameters—Modified Gompertz Model
3. Materials and Methods
3.1. Bacterial Sludge
3.2. Culture Medium and Fermentation Process
3.3. Hydrogen Measure and Production
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Name | Units |
|---|---|---|
| Ratio of H and fH according to Equation (13) | L−1 | |
| Rate parameter of ln(fH) vs. t non-linear fit | h−1 | |
| A | Affinity towards hydrogen production | Mol |
| Independent parameter of ln(fH) vs. t non-linear fit | - | |
| Base of natural logarithm | - | |
| Cumulated hydron conversion function | mol/L | |
| H | Cumulated hydrogen production | Mol |
| Hydron concentration | mol/L | |
| Acid dissociation equilibrium constant | ||
| n | Number of moles | Mol |
| ni | Electric charge of the i-th base anion | - |
| r | Correlation coefficient | - |
| Rm | Maximum hydrogen production rate | mol/h |
| pH | Negative base 10 logarithm of hydron concentration | - |
| Hydrogen production | mol/h | |
| Analytic phosphate concentration | mol/L | |
| t | Time | H |
| VR | Reactor working volume | L |
| Weak base | - | |
| Efficiency towards hydrogen production | - | |
| Lag phase time | H |
References
- Ritchie, H.; Roser, M.; Rosado, P. Energy Production and Consumption. Available online: https://ourworldindata.org/energy (accessed on 18 September 2022).
- Tranum, S. Powerless: India’s Energy Shortage and Its Impact; SAGE Publications India Pvt Ltd.: Los Angeles, CA, USA, 2013; ISBN 978-81-321-1314-0. [Google Scholar]
- Kalvelage, K.; Passe, U.; Rabideau, S.; Takle, E.S. Changing Climate: The Effects on Energy Demand and Human Comfort. Energy Build. 2014, 76, 373–380. [Google Scholar] [CrossRef]
- Höök, M.; Tang, X. Depletion of Fossil Fuels and Anthropogenic Climate Change—A Review. Energy Policy 2013, 52, 797–809. [Google Scholar] [CrossRef] [Green Version]
- Kartal, M.T. The Role of Consumption of Energy, Fossil Sources, Nuclear Energy, and Renewable Energy on Environmental Degradation in Top-Five Carbon Producing Countries. Renew. Energy 2022, 184, 871–880. [Google Scholar] [CrossRef]
- Auzanneau, M.; Reynolds, J.F.; Heinberg, R. Oil, Power, and War: A Dark History; Chelsea Green Publishing: Hartford, VT, USA; Post Carbon Institute: Corvallis, OR, USA, 2018; ISBN 978-1-60358-743-3. [Google Scholar]
- Conrad, R.; Reinhardt, T. Hydrogen Strategy: Enabling A Low-Carbon Economy; Office of Fossil Energy, US Department of Energy: Washington, DC, USA, 2020. Available online: https://www.energy.gov/fecm/downloads/hydrogen-strategy-enabling-low-carbon-economy (accessed on 18 September 2022).
- Łukajtis, R.; Hołowacz, I.; Kucharska, K.; Glinka, M.; Rybarczyk, P.; Przyjazny, A.; Kamiński, M. Hydrogen Production from Biomass Using Dark Fermentation. Renew. Sustain. Energy Rev. 2018, 91, 665–694. [Google Scholar] [CrossRef]
- Jørgensen, S.E.; Fath, B.D. Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2008; ISBN 978-0-08-045405-4. [Google Scholar]
- Ajanovic, A.; Sayer, M.; Haas, R. The Economics and the Environmental Benignity of Different Colors of Hydrogen. Int. J. Hydrogen Energy 2022, 47, 24136–24154. [Google Scholar] [CrossRef]
- Moussa, R.N.; Moussa, N.; Dionisi, D. Hydrogen Production from Biomass and Organic Waste Using Dark Fermentation: An Analysis of Literature Data on the Effect of Operating Parameters on Process Performance. Processes 2022, 10, 156. [Google Scholar] [CrossRef]
- García, R.E.; Martínez, V.L.; Franco, J.I.; Curutchet, G. Selection of Natural Bacterial Communities for the Biological Production of Hydrogen. Int. J. Hydrogen Energy 2012, 37, 10095–10100. [Google Scholar] [CrossRef]
- Islam, A.K.M.K.; Dunlop, P.S.M.; Hewitt, N.J.; Lenihan, R.; Brandoni, C. Bio-Hydrogen Production from Wastewater: A Comparative Study of Low Energy Intensive Production Processes. Clean Technol. 2021, 3, 156–182. [Google Scholar] [CrossRef]
- García, R.E.; Pin Viso, N.; Gerosa, F.A.; Nishinakamasu, V.; Puebla, A.F.; Farber, M.D.; Lavorante, M.J. Argentine Navy Icebreaker Ship “Almirante Irizar” Sludge Microbial Composition Analysis for Biohydrogen Production. Bioenerg. Res. 2022, 1–12. [Google Scholar] [CrossRef]
- Wang, J.; Yin, Y. Principle and Application of Different Pretreatment Methods for Enriching Hydrogen-Producing Bacteria from Mixed Cultures. Int. J. Hydrogen Energy 2017, 42, 4804–4823. [Google Scholar] [CrossRef]
- Martínez, V.L.; García, R.E.; Curutchet, G.; Sanguinetti, A.; Fasoli, H.J.; Franco, J.I. Demonstration of the Possibility to Power a Fuel Cell with Hydrogen Derived from the Fermentation of Sugar. Int. J. Hydrogen Energy 2012, 37, 14920–14925. [Google Scholar] [CrossRef]
- Mona, S.; Kumar, S.S.; Kumar, V.; Parveen, K.; Saini, N.; Deepak, B.; Pugazhendhi, A. Green Technology for Sustainable Biohydrogen Production (Waste to Energy): A Review. Sci. Total Environ. 2020, 728, 138481. [Google Scholar] [CrossRef] [PubMed]
- Sekoai, P.T.; Daramola, M.O.; Mogwase, B.; Engelbrecht, N.; Yoro, K.O.; Petrus du Preez, S.; Mhlongo, S.; Ezeokoli, O.T.; Ghimire, A.; Ayeni, A.O.; et al. Revising the Dark Fermentative H2 Research and Development Scenario—An Overview of the Recent Advances and Emerging Technological Approaches. Biomass Bioenergy 2020, 140, 105673. [Google Scholar] [CrossRef]
- Wei, X.; Wang, R.-Z.; Zhao, W.; Chen, G.; Chai, M.-R.; Zhang, L.; Zhang, J. Recent Research Progress in PEM Fuel Cell Electrocatalyst Degradation and Mitigation Strategies. EnergyChem 2021, 3, 100061. [Google Scholar] [CrossRef]
- Sun, Y.; He, J.; Yang, G.; Sun, G.; Sage, V. A Review of the Enhancement of Bio-Hydrogen Generation by Chemicals Addition. Catalysts 2019, 9, 353. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Hidalgo, A.M.; Smoliński, A.; Sanchez, A. A Meta-Analysis of Research Trends on Hydrogen Production via Dark Fermentation. Int. J. Hydrogen Energy 2022, 47, 13300–13339. [Google Scholar] [CrossRef]
- Hallenbeck, P.C.; Abo-Hashesh, M.; Ghosh, D. Strategies for Improving Biological Hydrogen Production. Bioresour. Technol. 2012, 110, 1–9. [Google Scholar] [CrossRef]
- García-Depraect, O.; Rene, E.R.; Gómez-Romero, J.; López-López, A.; León-Becerril, E. Enhanced Biohydrogen Production from the Dark Co-Fermentation of Tequila Vinasse and Nixtamalization Wastewater: Novel Insights into Ecological Regulation by PH. Fuel 2019, 253, 159–166. [Google Scholar] [CrossRef]
- Aghajani Delavar, M.; Wang, J. Numerical Investigation of PH Control on Dark Fermentation and Hydrogen Production in a Microbioreactor. Fuel 2021, 292, 120355. [Google Scholar] [CrossRef]
- Chen, Y.; Yin, Y.; Wang, J. Influence of Butyrate on Fermentative Hydrogen Production and Microbial Community Analysis. Int. J. Hydrogen Energy 2021, 46, 26825–26833. [Google Scholar] [CrossRef]
- Penniston, J.; Gueguim Kana, E.B. Impact of Medium PH Regulation on Biohydrogen Production in Dark Fermentation Process Using Suspended and Immobilized Microbial Cells. Biotechnol. Biotechnol. Equip. 2018, 32, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Zagrodnik, R.; Duber, A.; Seifert, K. Dark-Fermentative Hydrogen Production from Synthetic Lignocellulose Hydrolysate by a Mixed Bacterial Culture: The Relationship between Hydraulic Retention Time and PH Conditions. Bioresour. Technol. 2022, 358, 127309. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.J.; Fuentes, K.L.; Alberto Arias, C.; Chaparro, T.R. Production of Hydrogen from Beverage Wastewater by Dark Fermentation in an Internal Circulation Reactor: Effect on PH and Hydraulic Retention Time. Energy Convers. Manag. X 2022, 15, 100232. [Google Scholar] [CrossRef]
- Wiechmann, A.; Müller, V. Energy Conservation in the Acetogenic Bacterium Clostridium Aceticum. Microorganisms 2021, 9, 258. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Yu, H.-Q.; Wang, G. A Kinetic Approach to Anaerobic Hydrogen-Producing Process. Water Res. 2007, 41, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Nemestóthy, N.; Bakonyi, P.; Rózsenberszki, T.; Kumar, G.; Koók, L.; Kelemen, G.; Kim, S.-H.; Bélafi-Bakó, K. Assessment via the Modified Gompertz-Model Reveals New Insights Concerning the Effects of Ionic Liquids on Biohydrogen Production. Int. J. Hydrogen Energy 2018, 43, 18918–18924. [Google Scholar] [CrossRef]
- Wang, J.; Wan, W. Kinetic Models for Fermentative Hydrogen Production: A Review. Int. J. Hydrogen Energy 2009, 34, 3313–3323. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, L.; Hao, Q.; Li, X.; Liu, F. Ferrihydrite Reduction Exclusively Stimulated Hydrogen Production by Clostridium with Community Metabolic Pathway Bifurcation. ACS Sustain. Chem. Eng. 2020, 8, 7574–7580. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van’t Riet, K. Modeling of the Bacterial Growth Curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [Green Version]
- Turhal, S.; Turanbaev, M.; Argun, H. Hydrogen Production from Melon and Watermelon Mixture by Dark Fermentation. Int. J. Hydrogen Energy 2019, 44, 18811–18817. [Google Scholar] [CrossRef]
- Martinez, V.; Alfonso, A.; García, R.; Lavorante, M.; Galvagno, M.; Cassanello, M. Design of Experiments to Optimize Hydrogen Production through Dark Fermentation of Sugars. In Proceedings of the 8th Symposium on Hydrogen, Fuel Cells and Advanced Batteries, Buenos Aires, Argentina, 11–14 July 2022. [Google Scholar]
- Lay, J. Influence of Chemical Nature of Organic Wastes on Their Conversion to Hydrogen by Heat-Shock Digested Sludge. Int. J. Hydrogen Energy 2003, 28, 1361–1367. [Google Scholar] [CrossRef]
- Lay, J.-J. Biohydrogen Generation by Mesophilic Anaerobic Fermentation of Microcrystalline Cellulose. Biotechnol. Bioeng. 2001, 74, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Gadhamshetty, V.; Arudchelvam, Y.; Nirmalakhandan, N.; Johnson, D.C. Modeling Dark Fermentation for Biohydrogen Production: ADM1-Based Model vs. Gompertz Model. Int. J. Hydrogen Energy 2010, 35, 479–490. [Google Scholar] [CrossRef]
- Kim, D.-H.; Yoon, J.-J.; Kim, S.-H.; Park, J.-H. Acceleration of Lactate-Utilizing Pathway for Enhancing Biohydrogen Production by Magnetite Supplementation in Clostridium Butyricum. Bioresour. Technol. 2022, 359, 127448. [Google Scholar] [CrossRef]
- Del Angel-Acosta, Y.A.; Alvarez, L.H.; Garcia-Reyes, R.B.; Carrillo-Reyes, J.; Garcia-Gonzalez, A.; Meza-Escalante, E.R. Co-Digestion of Corn (Nejayote) and Brewery Wastewater at Different Ratios and PH Conditions for Biohydrogen Production. Int. J. Hydrogen Energy 2021, 46, 27422–27430. [Google Scholar] [CrossRef]
- Logan, B.E.; Oh, S.-E.; Kim, I.S.; Van Ginkel, S. Biological Hydrogen Production Measured in Batch Anaerobic Respirometers. Environ. Sci. Technol. 2002, 36, 2530–2535. [Google Scholar] [CrossRef]








| Batch | λ (h) | Slope (L−1) | r2 | η (%) |
|---|---|---|---|---|
| I | 13 | 4.8 × 10−5 | 0.9452 | 0.041 |
| II | 13 | 7.7 × 10−5 | 0.9553 | 0.033 |
| III | 13 | 3.8 × 10−5 | 0.8934 | 0.043 |
| IV | 16 | 5.0 × 10−5 | 0.9788 | 0.040 |
| V | 29 | 9.4 × 10−5 | 0.8728 | 0.029 |
| Mean | 17 | 6.1 × 10−5 | 0.9291 | 0.037 |
| Std. dev. | 7 | 2.3 × 10−5 | 0.0443 | 0.006 |
| Batch | fit | A (mol) | Rm (mol/h) | λ (h) | r2 |
|---|---|---|---|---|---|
| I | H | 0.85 ± 0.39 | 0.011 ± 0.003 | 15.2 ± 6.1 | 0.9748 |
| 0.84 ± 0.01 | 0.018 ± 0.001 | 26.7 ± 0.5 | 0.9782 | ||
| II | H | 0.44 ± 0.07 | 0.011 ± 0.003 | 12.9 ± 5.3 | 0.9938 |
| 0.52 ± 0.01 | 0.011 ± 0.001 | 26.7 ± 0.5 | 0.9781 | ||
| III | H | 2.39 ± 1.02 | 0.017 ± 0.002 | 30.0 ± 0.1 | 0.9771 |
| 0.98 ± 0.01 | 0.016 ± 0.001 | 19.2 ± 0.4 | 0.9886 | ||
| IV | H | 0.99 ± 0.03 | 0.010 ± 0.001 | 12.2 ± 2.5 | 0.9986 |
| 1.05 ± 0.001 | 0.011 ± 0.001 | 22.2 ± 0.3 | 0.9953 | ||
| V | H | 0.87 ± 0.19 | 0.014 ± 0.008 | 20.8 ± 16.7 | 0.9713 |
| 1.23 ± 0.01 | 0.008 ± 0.001 | 23.2 ± 1.0 | 0.9685 |
| Batch | Fit | A (mol) | Rm (mol/h) | λ (h) | r2 |
|---|---|---|---|---|---|
| I | H | 0.85 ± 0.40 | 0.010 ± 0.002 | 13 | 0.9712 |
| 0.98 ± 0.04 | 0.012 ± 0.001 | 13 | 0.9474 | ||
| II | H | 0.44 ± 0.02 | 0.011 ± 0.001 | 13 | 0.9997 |
| 0.61 ± 0.02 | 0.007 ± 0.001 | 13 | 0.9473 | ||
| III | H | 1.97 ± 1.04 | 0.015 ± 0.008 | 13 | 0.9376 |
| 1.05 ± 0.02 | 0.014 ± 0.001 | 13 | 0.9800 | ||
| IV | H | 0.98 ± 0.04 | 0.011 ± 0.001 | 16 | 0.9973 |
| 1.07 ± 0.001 | 0.009 ± 0.001 | 16 | 0.9922 | ||
| V | H | 0.85 ± 0.20 | 0.018 ± 0.005 | 29 | 0.9518 |
| 1.20 ± 0.01 | 0.009 ± 0.001 | 29 | 0.9469 |
| Conditions | Gompertz Parameters * | Ref. | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Inoculum | Substrate | VR. (L) | T (°C) | pH | Agitation | A (mmol H2/L Reactor) | Rm (mmol H2/L.h) | λ (h) | |
| Preheated anaerobic sludge from citrate producing wastewater | Sucrose | 5 | 30–45 | Regulated at 5.5 | 120 rpm | 168 ± 7 | 15 ± 1 | 9.8 ± 0.3 | [30] |
| Heat-shocked digester sludge | Microcrystalline Cellulose | 0.12 | 37 | 7.0 (initial) | 1.5 rpm | 1.63 ± 0.07 | 0.29 ± 0.02 | 4.6 ± 0.3 | [30,37,38] ** |
| Clostridium-rich sludge from a pig manure digester | Rice organic waste | 30 ± 2 | 0.17 ± 0.01 | 54 ± 2 | |||||
| Potato organic waste | 25 ± 3 | 0.55 ± 0.06 | 110 ± 10 | ||||||
| Compost-based and low-pH inocula | Glucose and sucrose solutions | 0.15–0.175 | 22–30 | 7–8.5 (initial) | Unstirred vs. 160 rpm | 49–120 | 2.7–13.7 | 18–60 | [39] |
| Sea sediment clostridia community | Mineral salt—glucose | 0.04 | 30 | 3.25–4 (final) | None | 17 | 0.030 | 15.9 | [33] |
| Ferrihydrite amended | 20 | 0.033 | 15.2 | ||||||
| Ferrihydrite addition | 21 | 0.033 | 17.9 | ||||||
| Heat-treated anaerobic sludge | Melon and watermelon fruit mixture with 0.74 to 37 g/L total Solid concentration, with (wi) and without inoculation (woi) | 0.08 | 36 | 5.5–6 | 100 | 37 gTS/L | [24] | ||
| 218 (wi) 176 (woi) | 15.7 (wi) 3.6 (woi) | 3.65 (wi) 27.3 (woi) | |||||||
| 0.74 gTS/L | |||||||||
| 16 (wi) 6.7 (woi) | 0.5 (wi) 0.19 (woi) | 4.1 (wi) 76.4 (woi) | |||||||
| Clostridium Butirycum DSM 10,702 | Mineral salt—glucose | 0.1 | 37 | N/D | 60 rpm | 32.4 | 0.49 ± 0.05 | 4 | [40] |
| Fe3O4 addition | 40.7 | 0.93 ± 0.20 | 3.8 | ||||||
| Lactate addition | 38.0 | 0.72 ± 0.04 | 4.8 | ||||||
| Lactate + Fe3O4 addition | 51.4 | 0.76 ± 0.13 | 3.6 | ||||||
| Anaerobic sludge blanket | Brewery waste and corn waste mixtures | 0.08 | 35 | Variable or 6 (initial) | 150 rpm | 106–167 | 0.6–2.1 | 1.5–122 | [41] |
| Ship wastewater Clostridia, selected by thermotolerant evolution | Mineral salt—acetate media | 5 | 30 | 7–8.5 (initial) 4–4.5 (final) | 50 rpm | 204 ± 68 *** | 2.6 ± 0.6 *** | 13–29 | This work |
| 196 ± 44 **** | 2.2 ± 0.6 **** | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, V.L.; Salierno, G.L.; García, R.E.; Lavorante, M.J.; Galvagno, M.A.; Cassanello, M.C. Biological Hydrogen Production by Dark Fermentation in a Stirred Tank Reactor and Its Correlation with the pH Time Evolution. Catalysts 2022, 12, 1366. https://doi.org/10.3390/catal12111366
Martínez VL, Salierno GL, García RE, Lavorante MJ, Galvagno MA, Cassanello MC. Biological Hydrogen Production by Dark Fermentation in a Stirred Tank Reactor and Its Correlation with the pH Time Evolution. Catalysts. 2022; 12(11):1366. https://doi.org/10.3390/catal12111366
Chicago/Turabian StyleMartínez, Verónica L., Gabriel L. Salierno, Rodrigo E. García, María José Lavorante, Miguel A. Galvagno, and Miryan C. Cassanello. 2022. "Biological Hydrogen Production by Dark Fermentation in a Stirred Tank Reactor and Its Correlation with the pH Time Evolution" Catalysts 12, no. 11: 1366. https://doi.org/10.3390/catal12111366
APA StyleMartínez, V. L., Salierno, G. L., García, R. E., Lavorante, M. J., Galvagno, M. A., & Cassanello, M. C. (2022). Biological Hydrogen Production by Dark Fermentation in a Stirred Tank Reactor and Its Correlation with the pH Time Evolution. Catalysts, 12(11), 1366. https://doi.org/10.3390/catal12111366