

The Stability of Dimeric D-amino Acid Oxidase from Porcine Kidney Strongly Depends on the Buffer Nature and Concentration




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
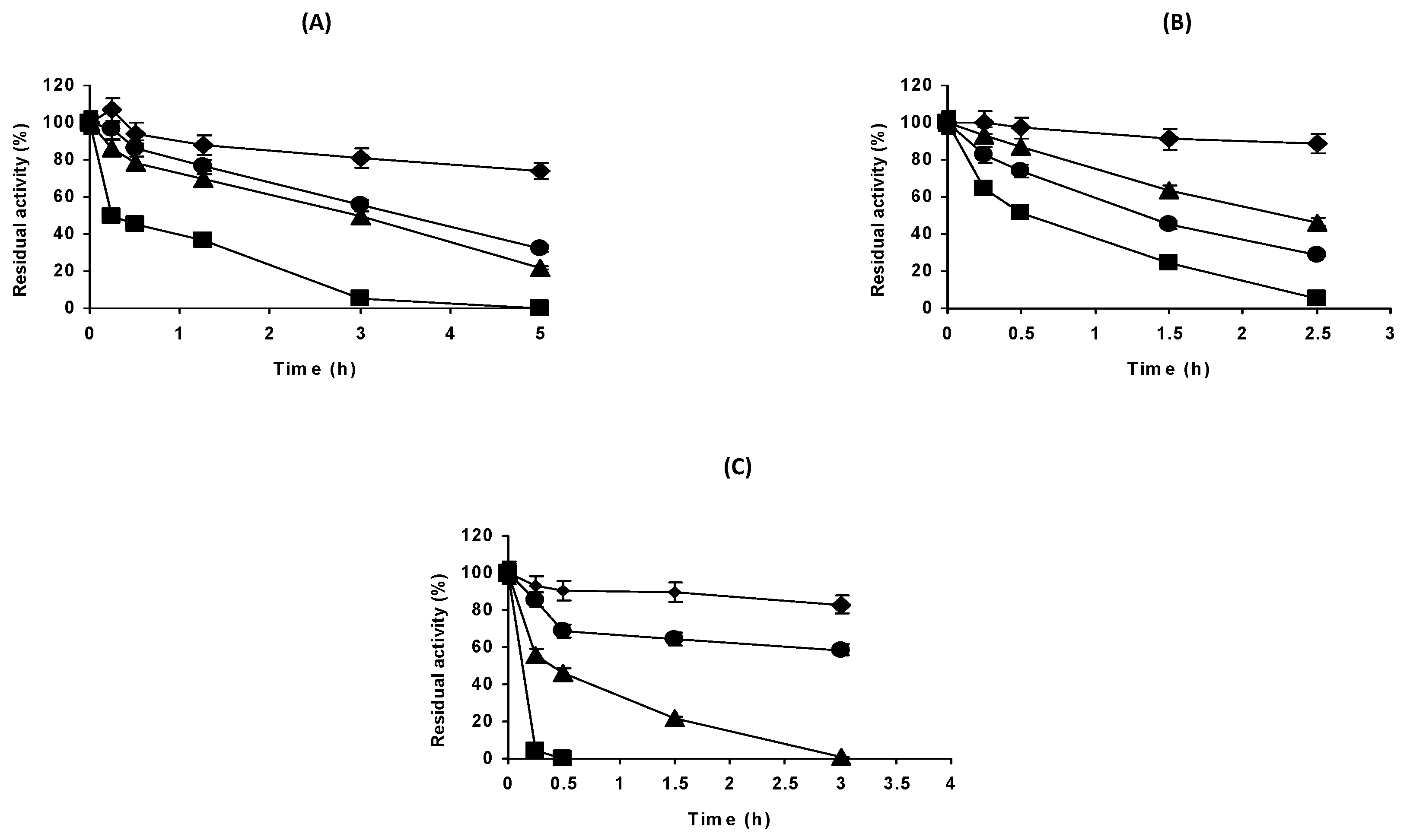
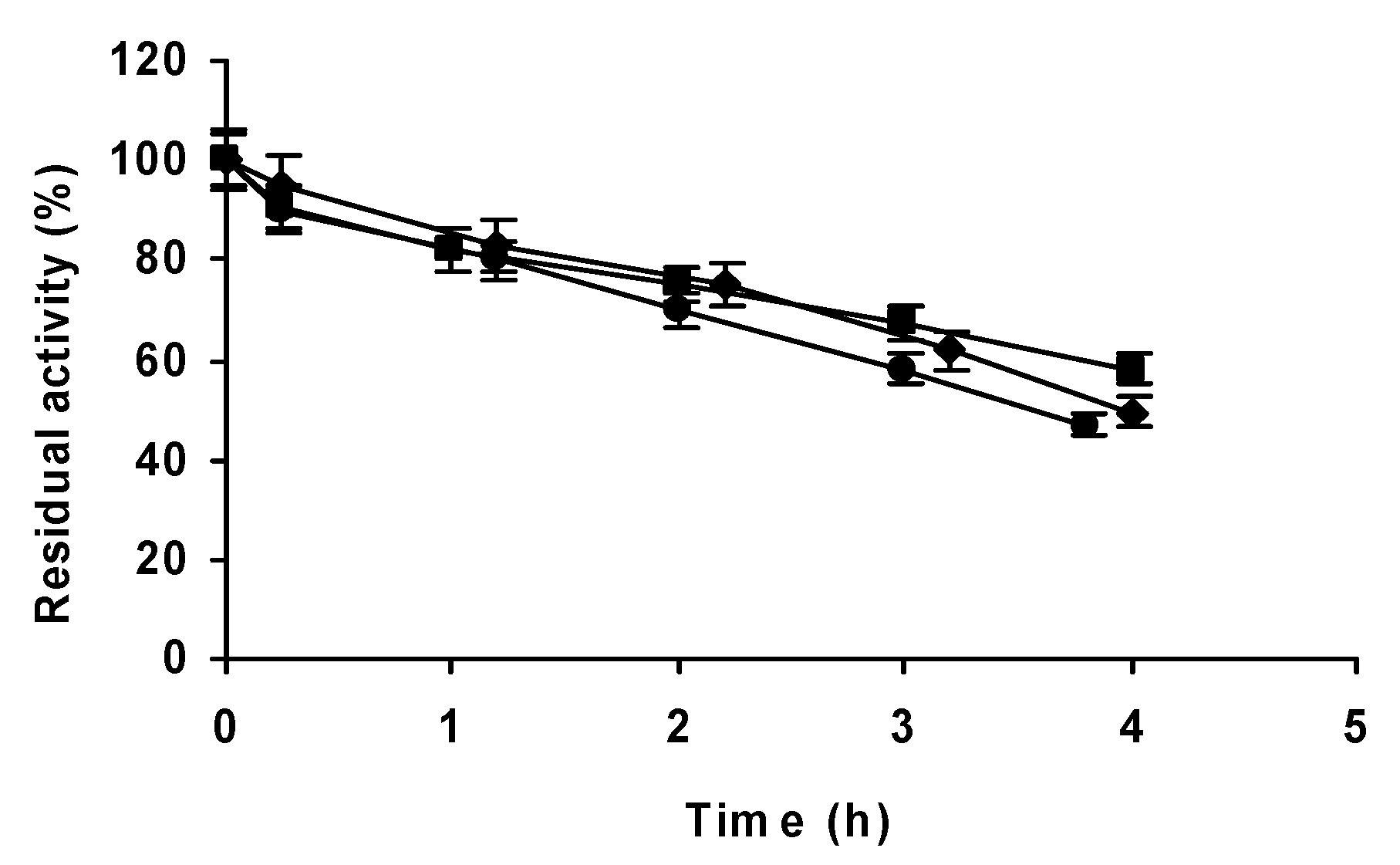
2.1. Effect of the Enzyme Concentration on the Stability of DAAO at Different pH Values
2.2. Effect of the Buffer Nature on the Stability of DAAO at pH 7
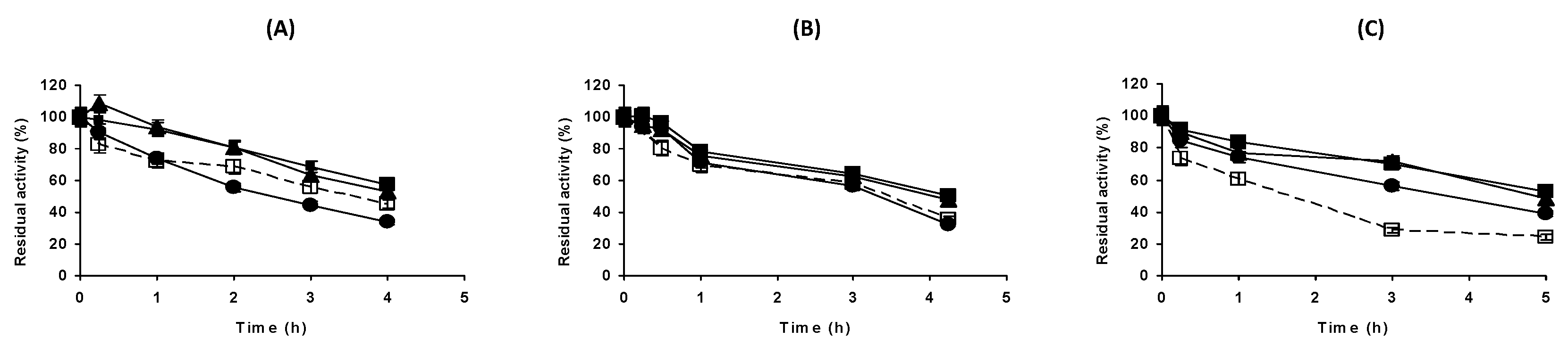
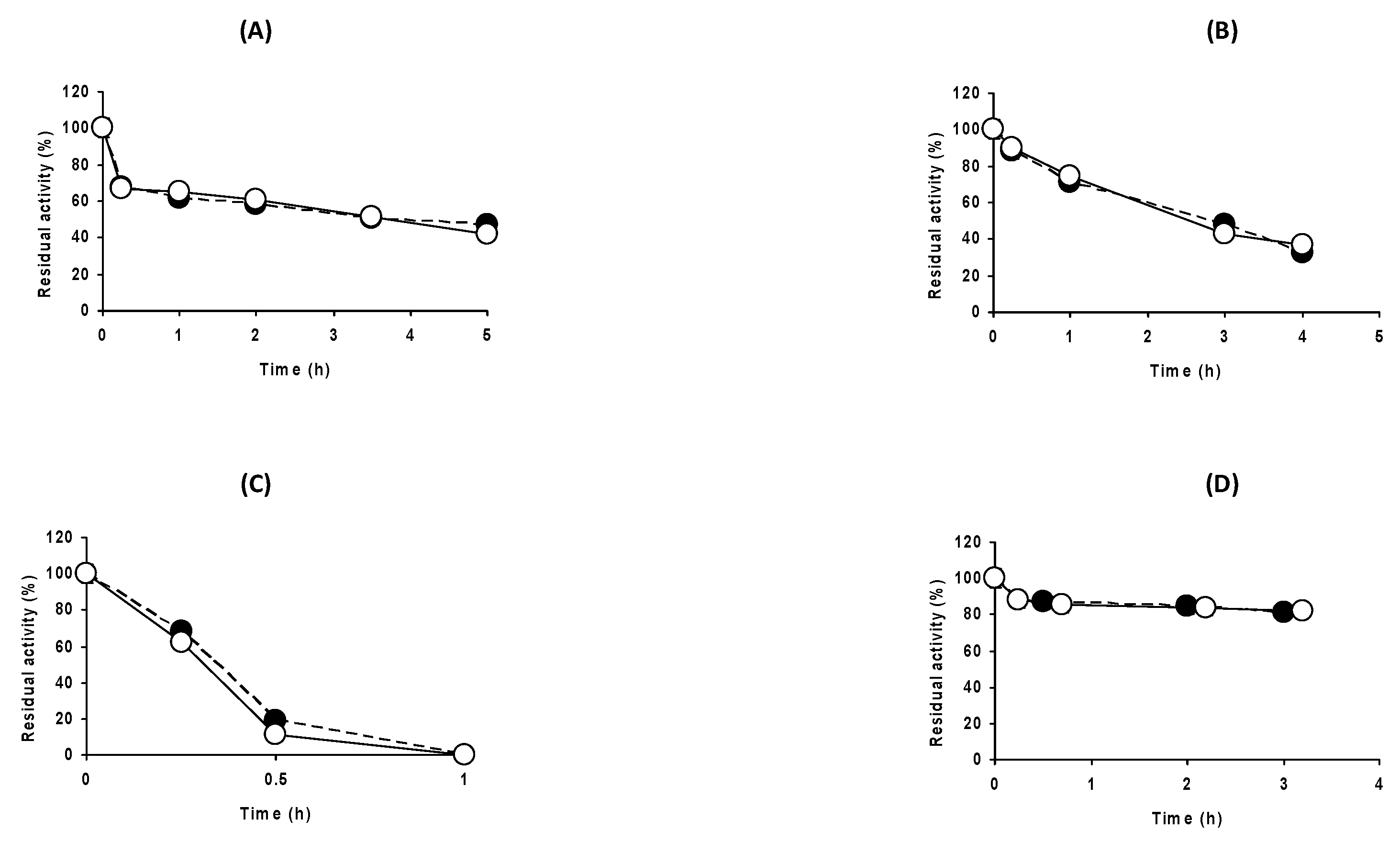
2.3. Effect of the Buffer Concentration at Different Enzyme Concentrations
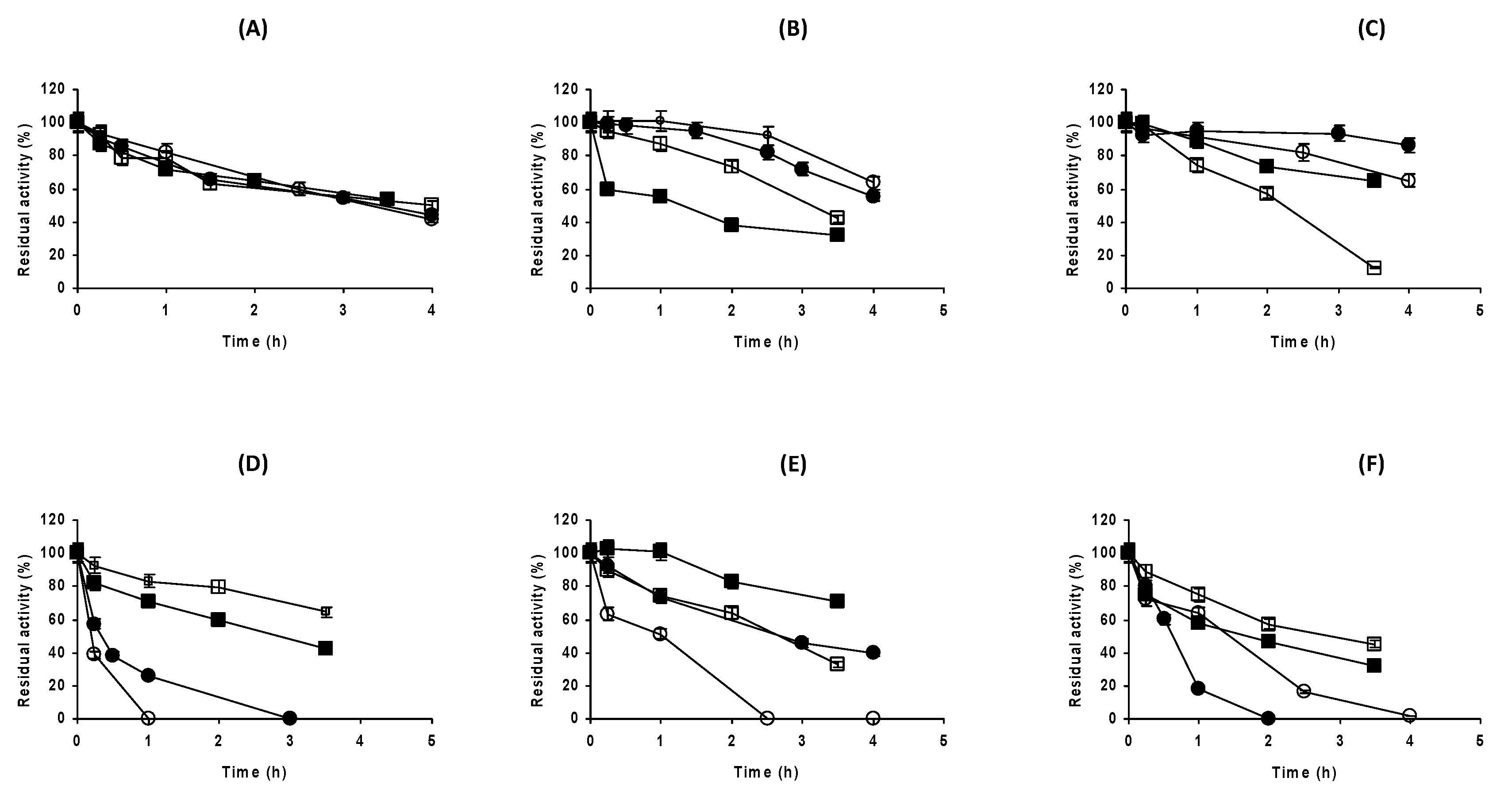
2.4. Effect of the Buffers’ Nature on the Stability of Immobilized DAAO When Enzyme Dissociation Was Not Possible
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Determination of DAAO Activity
3.2.2. Immobilization of DAAO on Glutaraldehyde-Amino-Agarose Beads
Preparation of Glutaraldehyde-Amino-Agarose Beads
Preparation of DAAO Glutaraldehyde-Amino-Agarose Beads
3.2.3. Thermal Inactivations of DAAO under Different Conditions
3.2.4. SDS-PAGE of Enzyme Preparations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tishkov, V.I.; Khoronenkova, S.V. D-Amino acid oxidase: Structure, catalytic mechanism, and practical application. Biochemistry 2005, 70, 40–54. [Google Scholar] [PubMed]
- Takamatsu, S.; Lee, I.; Lee, J.; Asano, R.; Tsugawa, W.; Ikebukuro, K.; Dick, J.E.; Sode, K. Transient potentiometry based D-serine sensor using engineered D-amino acid oxidase showing quasi-direct electron transfer property. Biosens. Bioelectron. 2022, 200, 113927. [Google Scholar] [CrossRef] [PubMed]
- Khoronenkova, S.V.; Tishkov, V.I. D-Amino acid oxidase: Physiological role and applications. Biochemistry 2008, 73, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Molla, G.; Sacchi, S.; Rosini, E.; Verga, R.; Pilone, M.S. Properties and applications of microbial D-Amino acid oxidases: Current state and perspectives. Appl. Microbiol. Biotechnol. 2008, 78, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Molla, G. New biotech applications from evolved D-Amino acid oxidases. Trends Biotechnol. 2011, 29, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.S.; Giesecke, U.; Reichert, A.; Minas, W. Industrial enzymatic production of cephalosporin-based β-Lactams. In Molecular Biotechnolgy of Fungal Beta-Lactam Antibiotics and Related Peptide Synthetases; Springer: Berlin/Heidelberg, Germany, 2004; pp. 179–215. [Google Scholar]
- Moussa, S.; Murtas, G.; Pollegioni, L.; Mauzeroll, J. Enhancing electrochemical biosensor selectivity with engineered D-Amino acid oxidase enzymes for D-Serine and D-Alanine quantification. ACS Appl. Bio. Mater. 2021, 4, 5598–5604. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, M.; Chen, L.; Zhang, F.; Yao, X.; Zhao, H.; Li, X. D-Amino acid electrochemical biosensor based on D-Amino acid oxidase: Mechanism and high performance against enantiomer interference. Biosens. Bioelectron. 2020, 151, 111971. [Google Scholar] [CrossRef] [PubMed]
- Nieh, C.H.; Kitazumi, Y.; Shirai, O.; Kano, K. Sensitive D-Amino acid biosensor based on oxidase/peroxidase system mediated by pentacyanoferrate-bound polymer. Biosens. Bioelectron. 2013, 47, 350–355. [Google Scholar] [CrossRef]
- Lata, S.; Batra, B.; Kumar, P.; Pundir, C.S. Construction of an amperometric D-Amino acid biosensor based on D-Amino acid oxidase/carboxylated mutliwalled carbon nanotube/copper nanoparticles/polyalinine modified gold electrode. Anal. Biochem. 2013, 437, 1–9. [Google Scholar] [CrossRef]
- Upadhya, R.; Nagajyothi; Bhat, S.G. Stabilization of D-Amino acid oxidase and catalase in permeabilized Rhodotorula gracilis cells and its application for the preparation of α-Ketoacids. Biotechnol. Bioeng. 2000, 68, 430–436. [Google Scholar] [CrossRef]
- Upadhya, R.; Nagajyothi, H.; Bhat, S.G. D-Amino acid oxidase and catalase of detergent permeabilized Rhodotorula gracilis cells and its potential use for the synthesis of α-keto acids. Process Biochem. 1999, 35, 7–13. [Google Scholar] [CrossRef]
- Fernández-Lafuente, R.; Rodriguez, V.; Guisán, J.M. The coimmobilization of D-Amino acid oxidase and catalase enables the quantitative transformation of D-Amino acids (D-Phenylalanine) into α-Keto acids (phenylpyruvic acid). Enzym. Microb. Technol. 1998, 23, 28–33. [Google Scholar] [CrossRef]
- Butò, S.; Pollegioni, L.; D’Angiuro, L.; Pilone, M.S. Evaluation of D-Amino acid oxidase from Rhodotorula gracilis for the production of α-keto acids: A reactor system. Biotechnol. Bioeng. 1994, 44, 1288–1294. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.S.; Fong, W.P.; Tsang, P.W.K. Entrapment of a Trigonopsis variabilis D-Amino acid oxidase variant F54Y for oxidative deamination of cephalosporin C. Eng. Life Sci. 2011, 11, 491–495. [Google Scholar] [CrossRef]
- Yu, H.L.; Li-Gong, C.; Yang, L.; Xi-Long, Y.; Guo-Yi, B.; Xue-Ming, D. Effect of hydrogen peroxide on the conversion of cephalosporin C to glutaryl-7-ACA by D-Amino acid oxidase. J. Chem. Eng. Chin. Univ. 2006, 20, 79–84. [Google Scholar]
- Shih-Yun, L.; Wang, J.D.; Lin, J.H. Expression of Trigonopsis variabilis D-Amino acid oxidase in transgenic rice for cephalosporin production. Bot. Stud. 2009, 50, 181–192. [Google Scholar]
- Pollegioni, L.; Caldinelli, L.; Molla, G.; Sacchi, S.; Pilone, M.S. Catalytic properties of D-Amino acid oxidase in cephalosporin C bioconversion: A comparison between proteins from different sources. Biotechnol. Prog. 2004, 20, 467–473. [Google Scholar] [CrossRef]
- Volpato, G.; Rodrigues, R.C.; Fernandez-Lafuente, R. Use of enzymes in the production of semi-synthetic penicillins and cephalosporins: Drawbacks and perspectives. Curr. Med. Chem. 2010, 17, 3855–3873. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, K.; Berenguer-Murcia, A.; Rodrigues, R.C.; Fernandez-Lafuente, R. Hydrogen peroxide in biocatalysis. A dangerous liaison. Curr. Org. Chem. 2012, 16, 2652–2672. [Google Scholar] [CrossRef]
- Lopez-Gallego, F.; Batencor, L.; Hidalgo, A.; Mateo, C.; Fernandez-Lafuente, R.; Guisan, J.M. One-pot conversion of cephalosporin C to 7-Aminocephalosporanic acid in the absence of hydrogen peroxide. Adv. Synth. Catal. 2005, 347, 1804–1810. [Google Scholar] [CrossRef]
- Luo, H.; Li, Q.; Yu, H.; Shen, Z. Construction and application of fusion proteins of D-Amino acid oxidase and glutaryl-7-Aminocephalosporanic acid acylase for direct bioconversion of cephalosporin C to 7-Aminocephalosporanic acid. Biotechnol. Lett. 2004, 26, 939–945. [Google Scholar] [CrossRef]
- Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Design of artificial enzymes bearing several active centers: New trends, opportunities and problems. Int. J. Mol. Sci. 2022, 23, 5304. [Google Scholar] [CrossRef]
- Mizutani, H.; Miyahara, I.; Hirotsu, K.; Nishina, Y.; Shiga, K.; Setoyama, C.; Miura, R. Three-Dimensional structure of porcine kidney D-Amino acid oxidase at 3.0 a resolution. J. Biochem. 1996, 120, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Fukui, K.; Watanabe, F.; Shibata, T.; Miyake, Y. Molecular cloning and sequence analysis of cDNAs encoding porcine kidney D-Amino acid oxidase. Biochemistry 1987, 26, 3612–3618. [Google Scholar] [CrossRef]
- Mizutani, H.; Miyahara, I.; Hirotsu, K.; Nishina, Y.; Shiga, K.; Setoyama, C.; Miura, R. Three-dimensional structure of the purple intermediate of porcine kidney D-Amino acid oxidase. Optimization of the oxidative half-reaction through alignment of the product with reduced flavin. J. Biochem. 2000, 128, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Setoyama, C.; Miura, R.; Nishina, Y.; Shiga, K.; Mizutani, H.; Miyahara, I.; Hirotsu, K. Crystallization of expressed porcine kidney D-Amino acid oxidase and preliminary X-ray crystallographic characterization. J. Biochem. 1996, 119, 1114–1117. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, K.; Motojima, F.; Ono, A.; Asano, Y. Expansion of the substrate specificity of porcine kidney D-Amino acid oxidase for S-stereoselective oxidation of 4-Cl-benzhydrylamine. ChemCatChem 2018, 10, 3500–3505. [Google Scholar] [CrossRef]
- Kawahara, N.; Yasukawa, K.; Asano, Y. New enzymatic methods for the synthesis of primary α-Aminonitriles and unnatural α-Amino acids by oxidative cyanation of primary amines with D-Amino acid oxidase from porcine kidney. Green Chem. 2017, 19, 418–424. [Google Scholar] [CrossRef]
- Yasukawa, K.; Kawahara, N.; Motojima, F.; Nakano, S.; Asano, Y. Porcine kidney D-Amino acid oxidase-derived R-Amine oxidases with new substrate specificities. Enzymes 2020, 47, 117–136. [Google Scholar]
- Kawahara, N.; Palasin, K.; Asano, Y. Novel enzymatic method for imine synthesis via the oxidation of primary amines using D-Amino acid oxidase from porcine kidney. Catalysts 2022, 12, 511. [Google Scholar] [CrossRef]
- Watanabe, F.; Fukui, K.; Momoi, K.; Miyake, Y. Expression of normal and abnormal porcine kidney D-Amino acid oxidases in Escherichia coli: Purification and characterization of the enzymes. Biochem. Biophys. Res. Commun. 1989, 165, 62–73. [Google Scholar] [CrossRef]
- Ciccarelli, E.; Massaer, M.; Guillaume, J.P.; Herzog, A.; Loriau, R.; Cravador, A.; Jacobs, P.; Bollen, A. Porcine D-Amino acid oxidase: Production of the biologically active enzyme in Escherichia coli. Biochem. Biophys. Res. Commun. 1989, 161, 865–872. [Google Scholar] [CrossRef]
- Bakke, M.; Setoyama, C.; Miura, R.; Kajiyama, N. Thermostabilization of porcine kidney D-Amino acid oxidase by a single Amino acid substitution. Biotechnol. Bioeng. 2006, 93, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Atroshenko, D.L.; Pometun, A.A.; Savin, S.S.; Tishkov, V.I. Determination of the kinetic parameters of a wild-type D-Amino acid oxidase from yeast and its mutant forms in a reaction of cephalosporin C oxidation. Mosc. Univ. Chem. Bull. 2019, 74, 169–172. [Google Scholar] [CrossRef]
- Poltorak, O.M.; Chukhray, E.S.; Torshin, I. Dissociative thermal inactivation, stability, and activity of oligomeric enzymes. Biochemistry 1998, 63, 303–311. [Google Scholar]
- Fernandez-Lafuente, R. Stabilization of multimeric enzymes: Strategies to prevent subunit dissociation. Enzym. Microb. Technol. 2009, 45, 405–418. [Google Scholar]
- Bolivar, J.M.; Rocha-Martin, J.; Mateo, C.; Cava, F.; Berenguer, J.; Fernandez-Lafuente, R.; Guisan, J.M. Coating of soluble and immobilized enzymes with ionic polymers: Full stabilization of the quaternary structure of multimeric enzymes. Biomacromolecules 2009, 10, 742–747. [Google Scholar] [CrossRef]
- Garcia-Galan, C.; Barbosa, O.; Fernandez-Lafuente, R. Stabilization of the hexameric glutamate dehydrogenase from Escherichia coli by cations and polyethyleneimine. Enzym. Microb. Technol. 2013, 52, 211–217. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Berenguer-Murcia, Á.; Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Stabilization of enzymes via immobilization: Multipoint covalent attachment and other stabilization strategies. Biotechnol. Adv. 2021, 52, 107821. [Google Scholar]
- Findrik, Z.; Tusić, M.; Vasić-Rački, D. Stabilization of D-Amino acid oxidase via covalent immobilization and mathematical model of D-Methionine oxidative deamination catalyzed by immobilized enzyme. Chem. Biochem. Eng. Q. 2016, 30, 93–102. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar]
- Garcia-Galan, C.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of different enzyme immobilization strategies to improve enzyme performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar]
- Di Cosimo, R.; Mc Auliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial use of immobilized enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar]
- Liese, A.; Hilterhaus, L. Evaluation of immobilized enzymes for industrial applications. Chem. Soc. Rev. 2013, 42, 6236–6249. [Google Scholar] [CrossRef] [PubMed]
- Bolivar, J.M.; Woodley, J.M.; Fernandez-Lafuente, R. Is Enzyme Immobilization a Mature Discipline? Some Critical Considerations to Capitalize on the Benefits of Immobilization. Chem. Soc. Rev. 2022. submitted. [Google Scholar]
- Betancor, L.; Hidalgo, A.; Fernández-Lorente, G.; Mateo, C.; Rodríguez, V.; Fuentes, M.; López-Gallego, F.; Fernández-Lafuente, R.; Guisan, J.M. Use of physicochemical tools to determine the choice of optimal enzyme: Stabilization of D-Amino acid oxidase. Biotechnol. Prog. 2003, 19, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Bauduin, P.; Nohmie, F.; Touraud, D.; Neueder, R.; Kunz, W.; Ninham, B.W. Hofmeister specific-ion effects on enzyme activity and buffer pH: Horseradish peroxidase in citrate buffer. J. Mol. Liq. 2006, 123, 14–19. [Google Scholar]
- Rej, R.; Vanderlinde, R.E. Effects of buffers on aspartate Aminotransferase activity and association of the enzyme with pyridoxal phosphate. Clin. Chem. 1975, 21, 1585–1591. [Google Scholar]
- Haije, W.G. Influence of buffer conditions on the activities of some iso-enzymes of alkaline phosphatase in the serum. Clin. Chim. Acta 1973, 48, 23–26. [Google Scholar]
- Garry, P.J. Serum cholinesterase variants: Examination of several differential inhibitors, salts, and buffers used to measure enzyme activity. Clin. Chem. 1971, 17, 183–191. [Google Scholar] [PubMed]
- Singh, S.; Sharma, S.; Agarwal, S.K. A simple purification procedure of buffalo lung cathepsin H, its properties and influence of buffer constituents on the enzyme activity. Biochem. Biophys. Rep. 2020, 22, 100739. [Google Scholar]
- Nicholas, P.C. Fructose-1,6-bisphosphate aldolases from guinea-pig cerebral cortex: Variation of enzyme activity with substrate concentration observed in Tris/HCl buffer. Biochem. Soc. Trans. 1989, 17, 227–228. [Google Scholar] [CrossRef]
- Zaak, H.; Fernandez-Lopez, L.; Velasco-Lozano, S.; Alcaraz-Fructuoso, M.T.; Sassi, M.; Lopez-Gallego, F.; Fernandez-Lafuente, R. Effect of high salt concentrations on the stability of immobilized lipases: Dramatic deleterious effects of phosphate anions. Process Biochem. 2017, 62, 128–134. [Google Scholar]
- Kornecki, J.F.; Carballares, D.; Morellon-Sterling, R.; Siar, E.H.; Kashefi, S.; Chafiaa, M.; Arana-Peña, S.; Rios, N.S.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Influence of phosphate anions on the stability of immobilized enzymes. Effect of enzyme nature, immobilization protocol and inactivation conditions. Process Biochem. 2020, 95, 288–296. [Google Scholar]
- Braham, S.A.; Siar, E.-H.; Arana-Peña, S.; Carballares, D.; Morellon-Sterling, R.; Bavandi, H.; de Andrades, D.; Kornecki, J.F.; Fernandez-Lafuente, R. Effect of concentrated salts solutions on the stability of immobilized enzymes: Influence of inactivation conditions and immobilization protocol. Molecules 2021, 26, 968. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Carballares, D.; Rocha-Martín, J.; Fernandez–Lafuente, R. Immobilization-Stabilization of the Dimeric D-Amino Acid Oxidase from Porcine Kidney. Process Biochem. submitted.
- Fernandez-Lafuente, R.; Rosell, C.M.; Rodriguez, V.; Santana, C.; Soler, G.; Bastida, A.; Guisán, J.M. Preparation of activated supports containing low pK Amino groups. A new tool for protein immobilization via the carboxyl coupling method. Enzym. Microb. Technol. 1993, 15, 546–550. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carballares, D.; Rocha-Martin, J.; Fernandez-Lafuente, R. The Stability of Dimeric D-amino Acid Oxidase from Porcine Kidney Strongly Depends on the Buffer Nature and Concentration. Catalysts 2022, 12, 1009. https://doi.org/10.3390/catal12091009
Carballares D, Rocha-Martin J, Fernandez-Lafuente R. The Stability of Dimeric D-amino Acid Oxidase from Porcine Kidney Strongly Depends on the Buffer Nature and Concentration. Catalysts. 2022; 12(9):1009. https://doi.org/10.3390/catal12091009
Chicago/Turabian StyleCarballares, Diego, Javier Rocha-Martin, and Roberto Fernandez-Lafuente. 2022. "The Stability of Dimeric D-amino Acid Oxidase from Porcine Kidney Strongly Depends on the Buffer Nature and Concentration" Catalysts 12, no. 9: 1009. https://doi.org/10.3390/catal12091009
APA StyleCarballares, D., Rocha-Martin, J., & Fernandez-Lafuente, R. (2022). The Stability of Dimeric D-amino Acid Oxidase from Porcine Kidney Strongly Depends on the Buffer Nature and Concentration. Catalysts, 12(9), 1009. https://doi.org/10.3390/catal12091009