

7α and 7β Hydroxylation of Dehydroepiandrosterone by Gibberella sp. and Absidia Coerulea Biotransformation

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
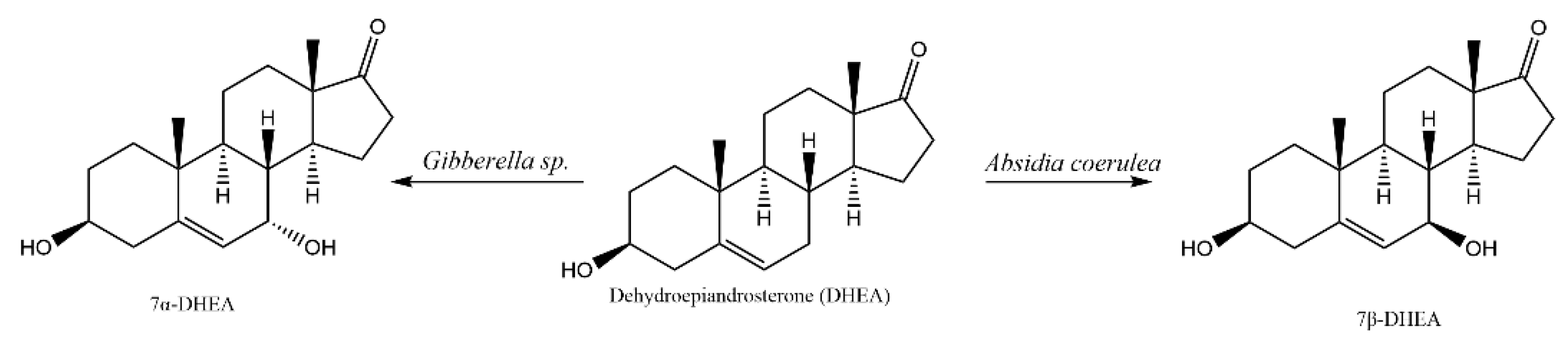
2.1. Whole-Cell Biotransformation Results of DHEA
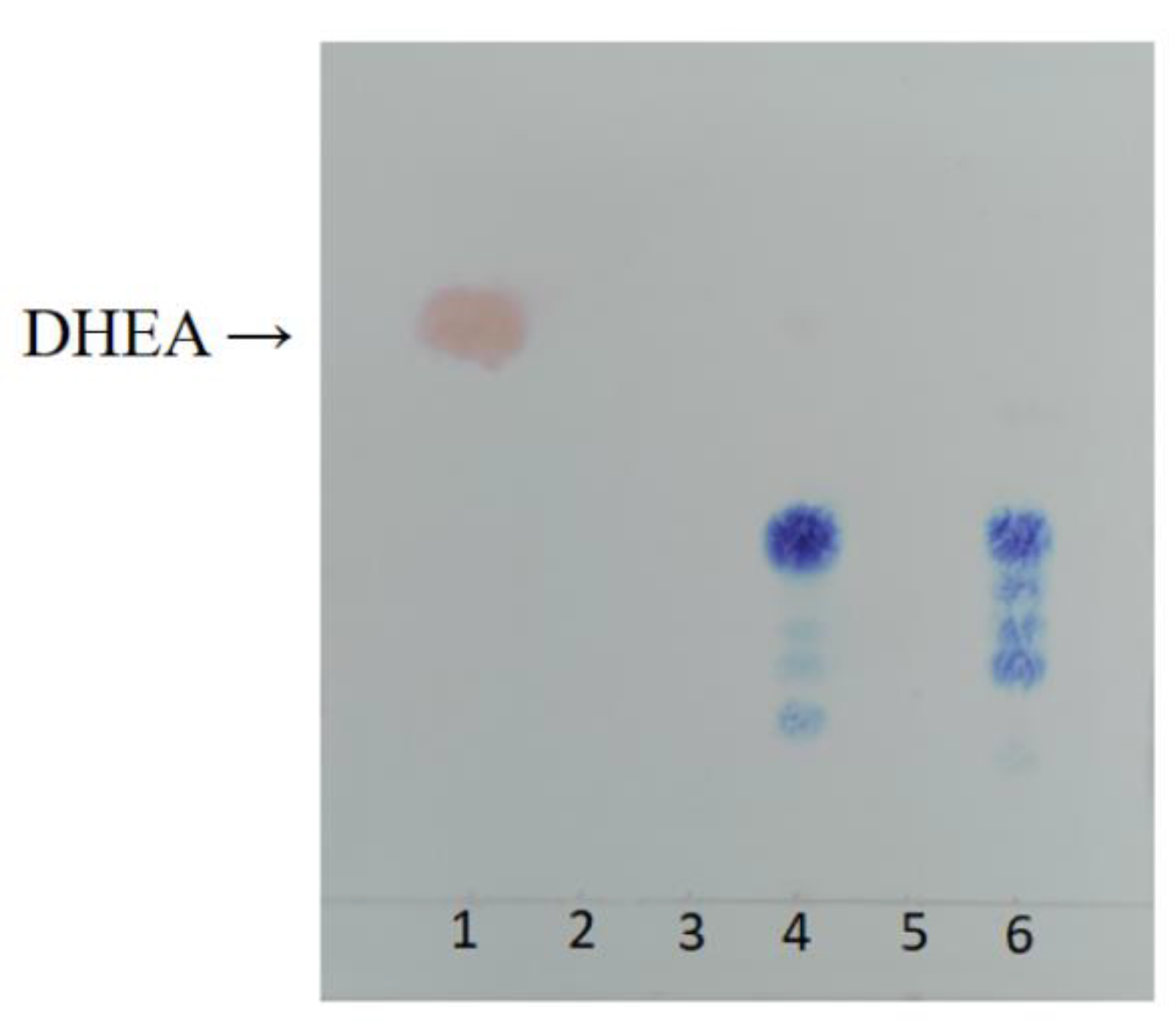
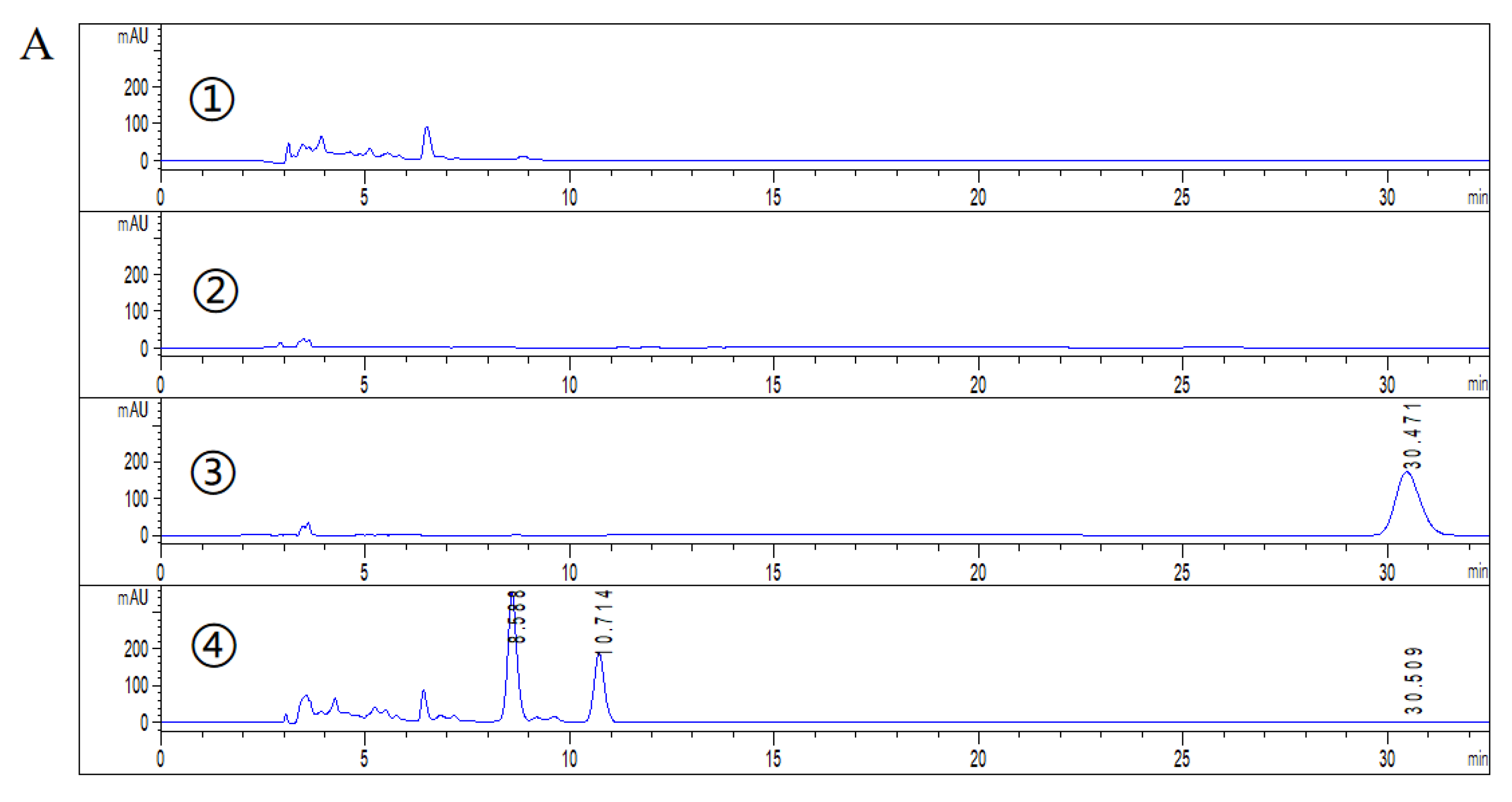
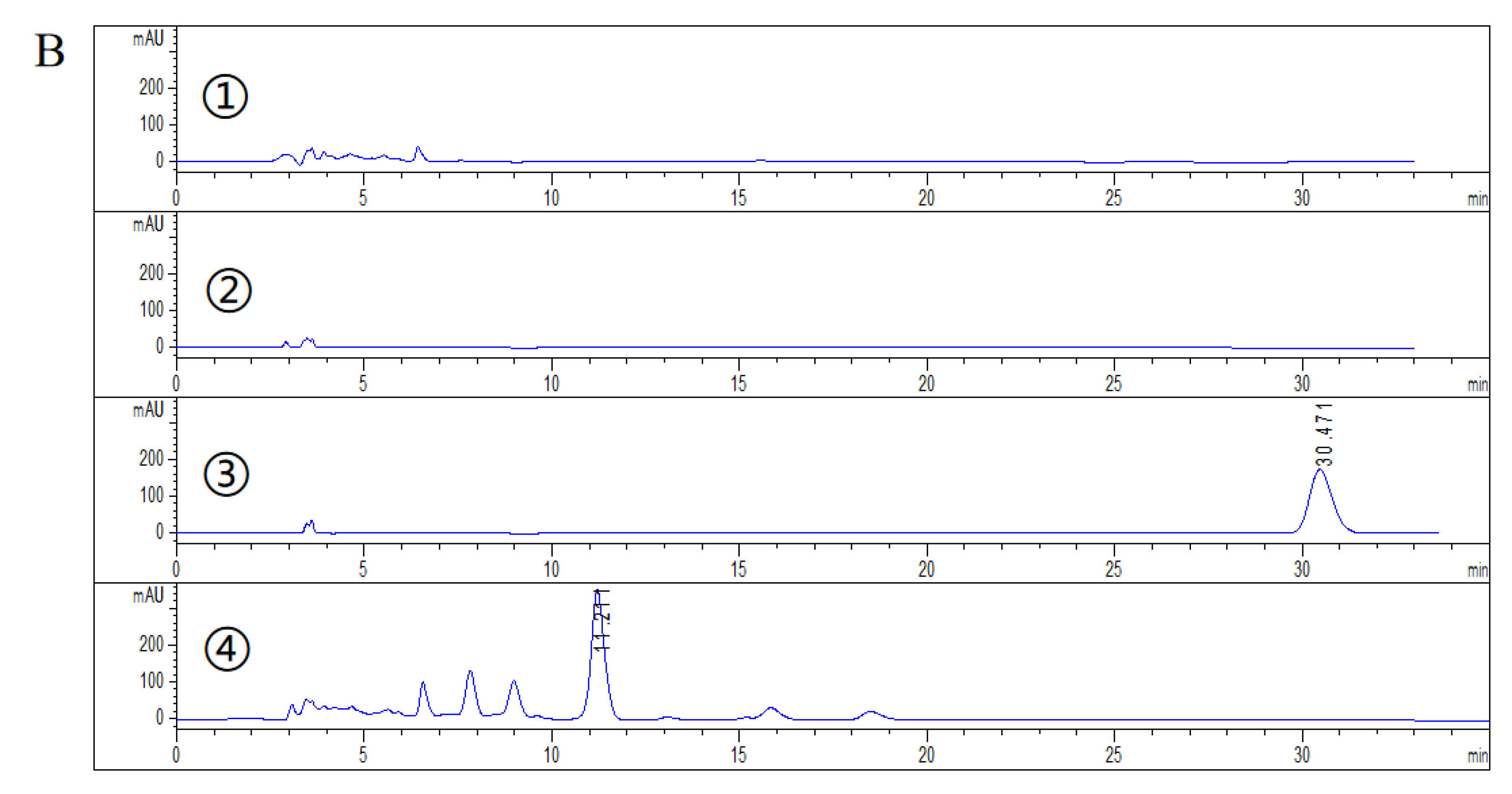
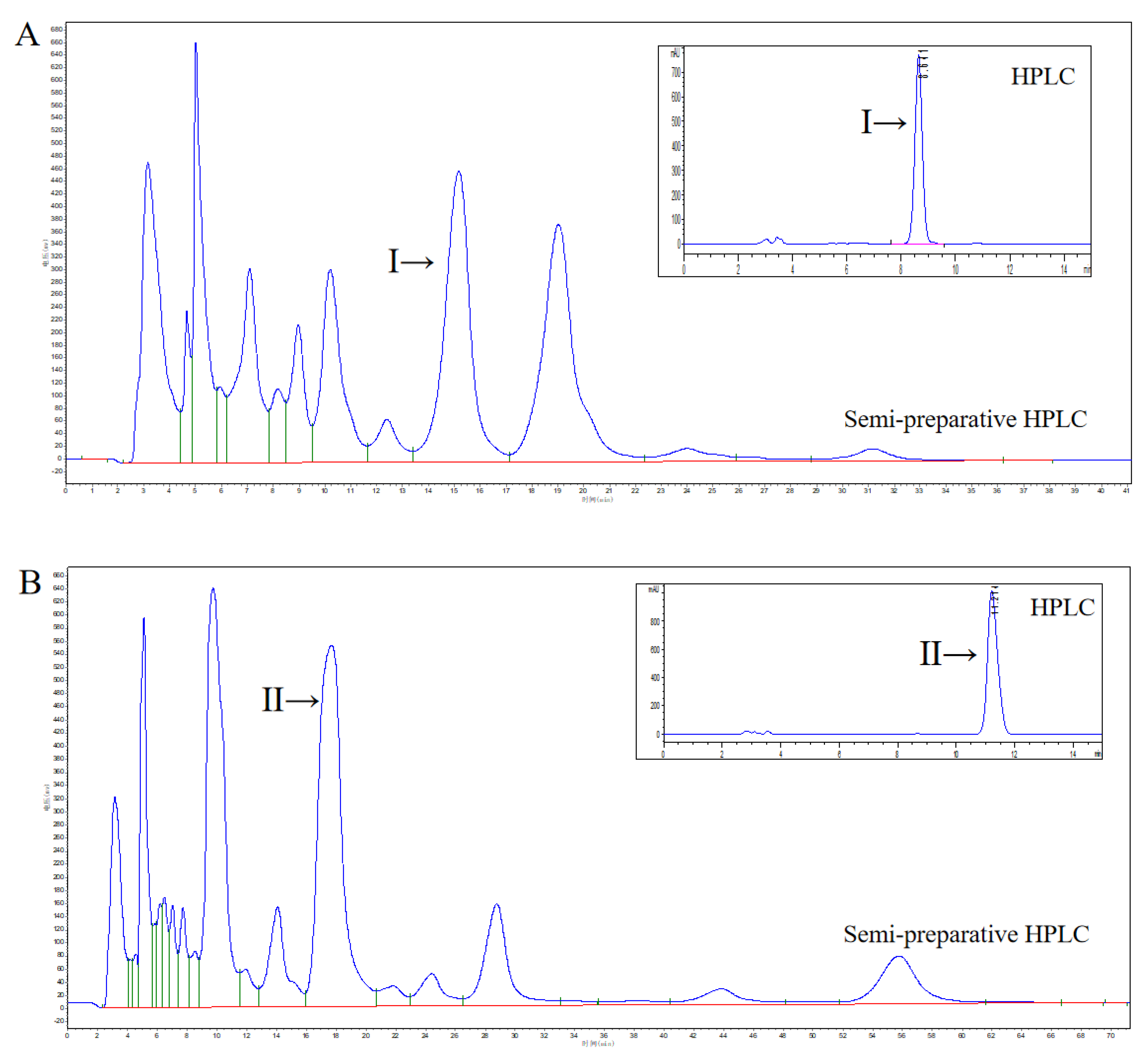
2.2. HPLC Analysis of DHEA Transformed by Gibberella sp. CICC 2498 and Absidia Coerulea CICC 41050
2.3. Isolation, Purification and Structural Identification of Metabolites
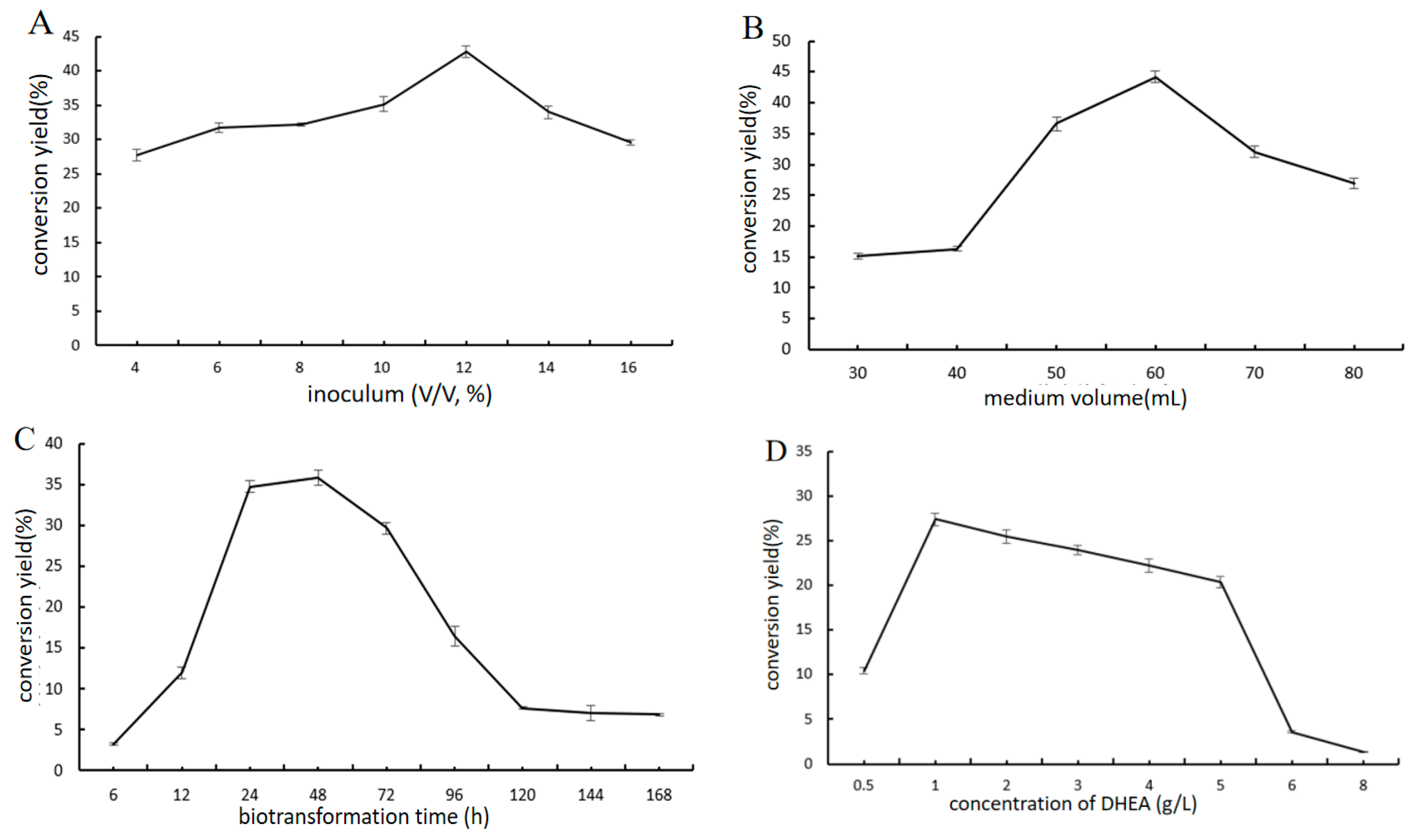
2.4. Optimization of 7β-OH-DHEA Production by Absidia Coerulea CICC 41050
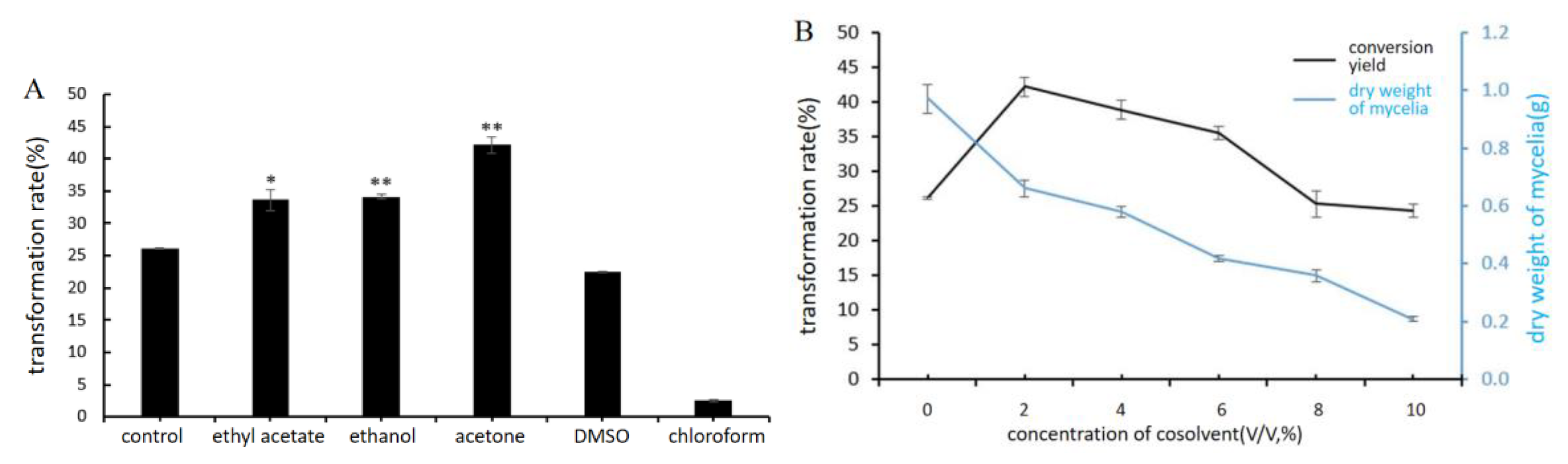
2.4.1. Influence of Different Cosolvents
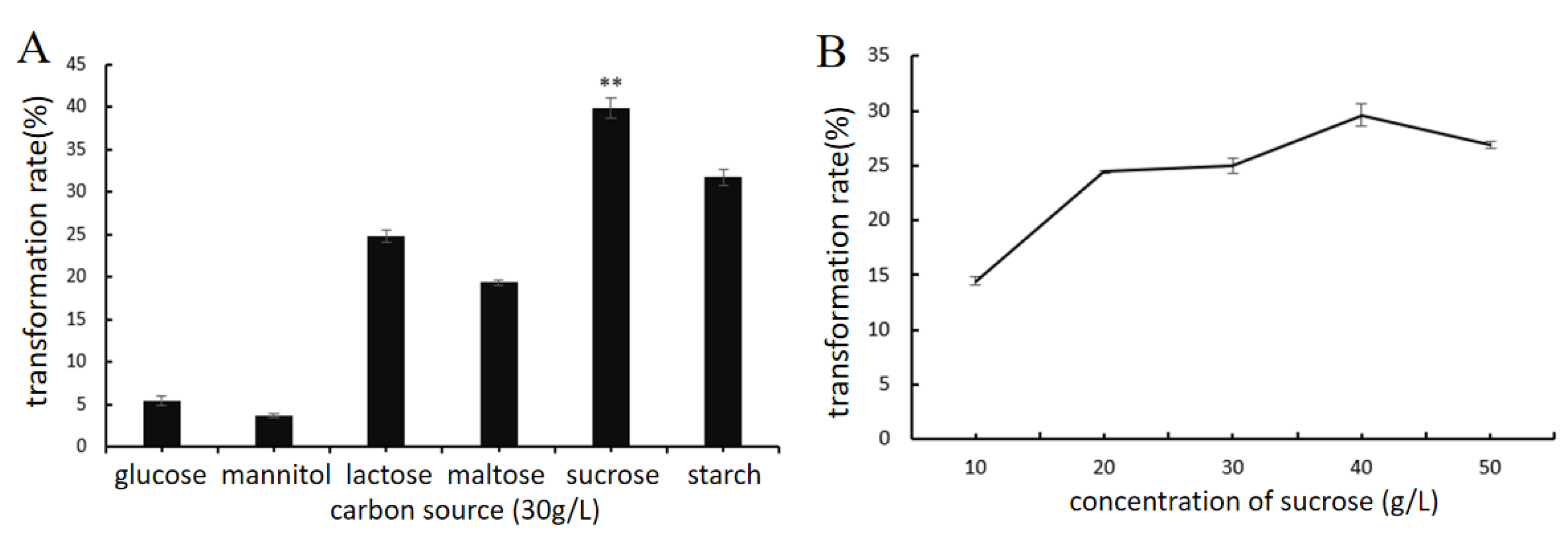
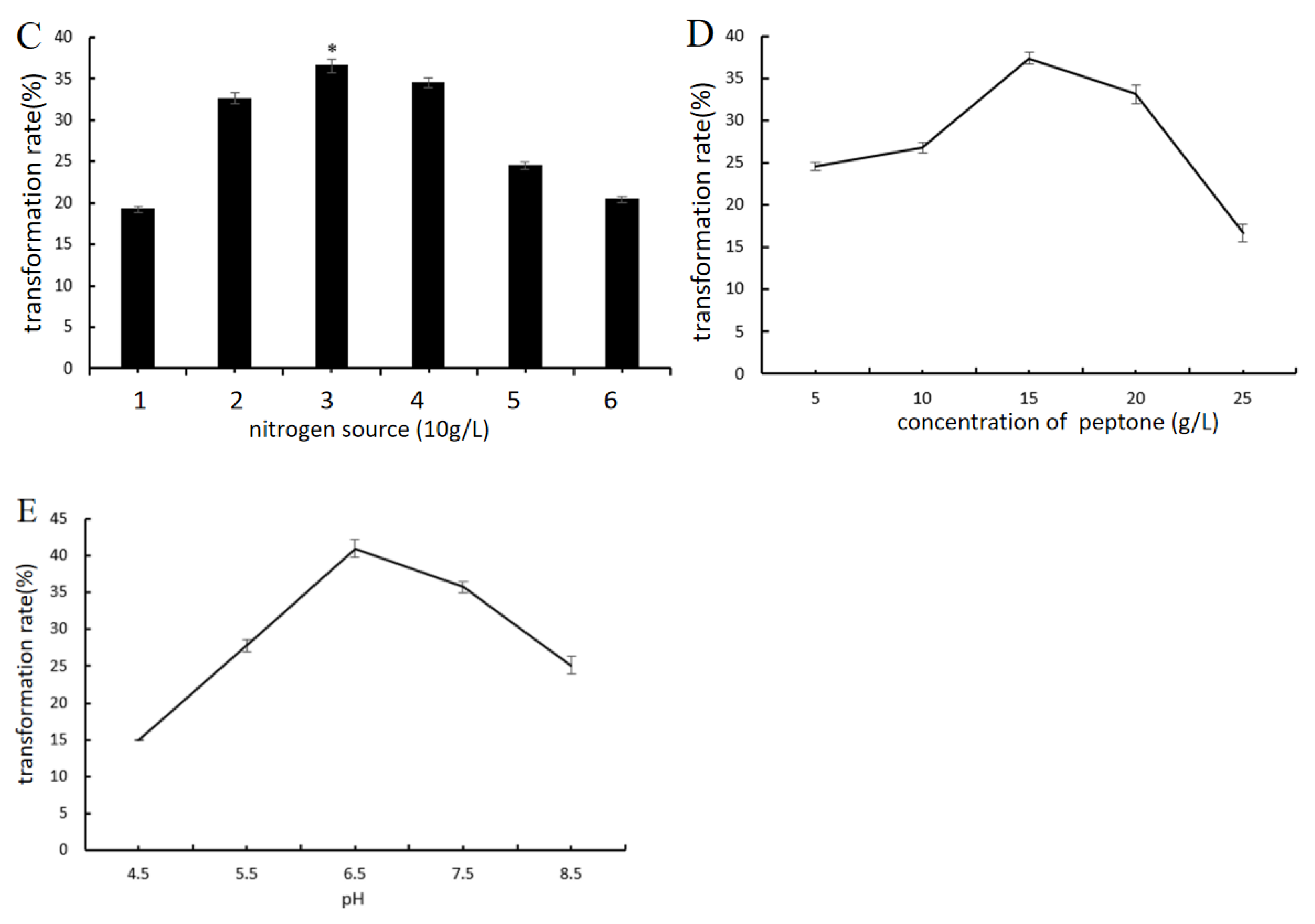
2.4.2. Effect of Key Nutrient Components and pH
2.4.3. Effect of Biotransformation Conditions
3. Materials and Methods
3.1. Chemicals
3.2. Microorganism and Cultivation
3.3. Sample Preparation
3.4. Thin Layer Chromatography (TLC)
3.5. HPLC Detection
3.6. Isolation and Identification of Major Metabolite
3.7. Establishment of Standard Curve and Calculation of Transformation Rate
3.8. Optimization of DHEA Converted to 7β-OH-DHEA by Absidia Coerulea CICC 41050
3.8.1. Effect of the Type and Concentration of Cosolvent on Transformation Rate
3.8.2. The Biotransformation Medium Was Studied by Single-Factor Experiment and Orthogonal Experiment
3.8.3. Biotransformation Conditions Were Studied by Single-Factor Experiment and Orthogonal Experiment
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- EMBL-EBI. CHEBI:28689—Dehydroepiandrosterone. Available online: http://www.ebi.ac.uk/chebi/searchId.do?chebiId=CHEBI:28689 (accessed on 1 January 2020).
- Corrigan, B. DHEA and Sport. Clin. J. Sport Med. 2002, 12, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.Y.; Cheng, A.J.; Chen, I.T.; Lan, T.Y.; Huang, I.H.; Shiau, C.W.; Hsu, C.L.; Liu, Y.W.; Chang, Z.F.; Tseng, P.H.; et al. Suppression of LPS-induced inflammatory responses by the hydroxyl groups of dexamethasone. Oncotarget 2017, 8, 49735–49748. [Google Scholar] [CrossRef] [PubMed]
- Heidary, M.; Habibi, Z. Microbial transformation of androst-4-ene-3,17-dione by three fungal species Absidia griseolla var. igachii, Circinella muscae and Trichoderma virens. J. Mol. Catal. B-Enzym. 2016, 126, 32–36. [Google Scholar] [CrossRef]
- Dragan, C.A.; Zearo, S.; Hannemann, F.; Bernhardt, R.; Bureik, M. Efficient conversion of 11-deoxycortisol to cortisol (hydrocortisone) by recombinant fission yeast Schizosaccharomyces pombe. FEMS Yeast Res. 2005, 5, 621–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumas, B.; Cauet, G.; Lacour, T.; Degryse, E.; Laruelle, L.; Ledoux, C.; Spagnoli, R.; Achstetter, T. 11 beta-hydroxylase activity in recombinant yeast mitochondria. In vivo conversion of 11-deoxycortisol to hydrocortisone. Eur. J. Biochem. 1996, 238, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Andryushina, V.A.; Voishvillo, N.E.; Druzhinina, A.V.; Stytsenko, T.S.; Yaderets, V.V.; Petrosyan, M.A.; Zeinalov, O.A. 14 alpha-Hydroxylation of steroids by mycelium of the mold fungus Curvularia lunata (VKPM F-981) to produce precursors for synthesizing new steroidal drugs. Pharm. Chem. J. 2013, 47, 103–108. [Google Scholar] [CrossRef]
- Faramarzi, M.A.; Tabatabaei Yazdi, M.; Amini, M.; Zarrini, G.; Shafiee, A. Microbial hydroxylation of progesterone with Acremonium strictum. FEMS Microbiol. Lett. 2003, 222, 183–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.-H.; Li, J.; Xu, G.; Zhang, X.H.; Wang, Y.G.; Yin, Y.L.; Liu, H.M. Biotransformation of dehydroepiandrosterone (DHEA) with Penicillium griseopurpureum Smith and Penicillium glabrum (Wehmer) Westling. Steroids 2010, 75, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Kolek, T.; Milecka, N.; Świzdor, A.; Panek, A.; Białońska, A. Hydroxylation of DHEA, androstenediol and epiandrosterone by Mortierella isabellina AM212. Evidence indicating that both constitutive and inducible hydroxylases catalyze 7 alpha- as well as 7beta-hydroxylations of 5-ene substrates. Org. Biomol. Chem. 2011, 9, 5414–5422. [Google Scholar] [CrossRef] [PubMed]
- Milecka-Tronina, N.; Kołek, T.; Świzdor, A.; Panek, A. Hydroxylation of DHEA and its analogues by Absidia coerulea AM93. Can an inducible microbial hydroxylase catalyze 7alpha- and 7beta-hydroxylation of 5-ene and 5alpha-dihydro C19-steroids? Bioorganic Med. Chem. 2014, 22, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Lyczko, P.; Panek, A.; Ceremuga, I.; Świzdor, A. The catalytic activity of mycelial fungi towards 7-oxo-DHEA—An endogenous derivative of steroidal hormone dehydroepiandrosterone. Microb. Biotechnol. 2021, 14, 2187–2198. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, K.; Kuru, A.; Yılmazer Keskin, S.; Ergin, S. Microbial transformation of dehydroepiandrosterone (DHEA) by some fungi. Biocatal. Biotransformation 2021, 39, 465–474. [Google Scholar] [CrossRef]
- Li, H.; Sun, J.; Xu, Z. Biotransformation of DHEA into 7α,15α-diOH-DHEA. In Microbial Steroids: Methods and Protocols; Barredo, J.-L., Herráiz, I., Eds.; Springer: New York, NY, USA, 2017; pp. 289–295. [Google Scholar]
- Kollerov, V.; Shutov, A.; Kazantsev, A.; Donova, M. Hydroxylation of pregnenolone and dehydroepiandrosterone by zygomycete Backusella lamprospora VKM F-944: Selective production of 7alpha-OH-DHEA. Appl. Microbiol. Biotechnol. 2022, 106, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, D.; Zapp, J.; Bernhardt, R. Steroid conversion with CYP106A2—Production of pharmaceutically interesting DHEA metabolites. Microb. Cell Factories 2014, 13, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Li, H.; Sun, J.; Zhang, X.; Shi, J.; Xu, Z. Production of 7alpha,15alpha-diOH-DHEA from dehydroepiandrosterone by Colletotrichum lini ST-1 through integrating glucose-feeding with multi-step substrate addition strategy. Bioprocess Biosyst. Eng. 2016, 39, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Sakaki, T. Practical application of cytochrome P450. Biol. Pharm. Bull. 2012, 35, 844–849. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Ability * | Microorganism | Ability * |
|---|---|---|---|
| Absidia coerulea CICC 41050 | + | Cunninghamella elegans 3.910 | − |
| Aspergillus fumigatus Aspergillus niger | − − | Gibberella fujikuroi CICC 40272 Gibberella sp. CICC 2498 | − + |
| Caldariomyces fumago CGMCC 16373 | − | Paecilomyces lilacinus | − |
| Cunninghamella blakesleeana 3.970 Cunninghamella echinulata 3.967 | − − | Penicillium Trichoderma virens CICC 2535 | − − |
| Factor | A/Sucrose (g/L) | B/Peptone (g/L) | C/Initial pH | |
|---|---|---|---|---|
| Level | ||||
| 1 | 30 | 10 | 5.5 | |
| 2 | 40 | 15 | 6.5 | |
| 3 | 50 | 20 | 7.5 | |
| Factor | A | B | C | Transformation Rate (%) | |
|---|---|---|---|---|---|
| No. | |||||
| 1 | 1 | 1 | 1 | 41.75 | |
| 2 | 1 | 2 | 2 | 40.48 | |
| 3 | 1 | 3 | 3 | 37.92 | |
| 4 | 2 | 1 | 2 | 39.80 | |
| 5 | 2 | 2 | 3 | 32.02 | |
| 6 | 2 | 3 | 1 | 28.75 | |
| 7 | 3 | 1 | 3 | 23.58 | |
| 8 | 3 | 2 | 1 | 39.29 | |
| 9 | 3 | 3 | 2 | 46.57 | |
| K1 | 40.05 | 35.04 | 36.60 | ||
| K2 | 33.52 | 37.26 | 42.28 | ||
| K3 | 36.48 | 37.75 | 31.17 | ||
| R | 6.53 | 2.70 | 11.11 | ||
| Factor | A/DHEA (g/L) | B/Time (h) | C/Medium Volume (mL) | D/Inoculum Rate (V/V, %) | |
|---|---|---|---|---|---|
| Level | |||||
| 1 | 1 | 24 | 50 | 10 | |
| 2 | 2 | 48 | 60 | 12 | |
| 3 | 3 | 72 | 70 | 14 | |
| Factor | A | B | C | D | Transformation Rate (%) | |
|---|---|---|---|---|---|---|
| No. | ||||||
| 1 | 1 | 1 | 1 | 1 | 53.62 | |
| 2 | 1 | 2 | 2 | 2 | 61.28 | |
| 3 | 1 | 3 | 3 | 3 | 62.81 | |
| 4 | 2 | 1 | 2 | 3 | 38.94 | |
| 5 | 2 | 2 | 3 | 1 | 47.61 | |
| 6 | 2 | 3 | 1 | 2 | 20.07 | |
| 7 | 3 | 1 | 3 | 2 | 30.03 | |
| 8 | 3 | 2 | 1 | 3 | 28.10 | |
| 9 | 3 | 3 | 2 | 1 | 42.73 | |
| K1 | 59.24 | 40.86 | 33.93 | 47.99 | ||
| K2 | 35.54 | 45.98 | 47.65 | 37.13 | ||
| K3 | 33.62 | 44.10 | 46.82 | 45.56 | ||
| R | 19.98 | 4.01 | 16.62 | 3.31 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, M.; Fu, R.; Cai, S.; Jiang, X.; Wang, F.; Xu, W.; Xu, W. 7α and 7β Hydroxylation of Dehydroepiandrosterone by Gibberella sp. and Absidia Coerulea Biotransformation. Catalysts 2023, 13, 272. https://doi.org/10.3390/catal13020272
Song M, Fu R, Cai S, Jiang X, Wang F, Xu W, Xu W. 7α and 7β Hydroxylation of Dehydroepiandrosterone by Gibberella sp. and Absidia Coerulea Biotransformation. Catalysts. 2023; 13(2):272. https://doi.org/10.3390/catal13020272
Chicago/Turabian StyleSong, Ming, Ruicheng Fu, Sulan Cai, Xuliang Jiang, Fuju Wang, Weizhuo Xu, and Wei Xu. 2023. "7α and 7β Hydroxylation of Dehydroepiandrosterone by Gibberella sp. and Absidia Coerulea Biotransformation" Catalysts 13, no. 2: 272. https://doi.org/10.3390/catal13020272
APA StyleSong, M., Fu, R., Cai, S., Jiang, X., Wang, F., Xu, W., & Xu, W. (2023). 7α and 7β Hydroxylation of Dehydroepiandrosterone by Gibberella sp. and Absidia Coerulea Biotransformation. Catalysts, 13(2), 272. https://doi.org/10.3390/catal13020272