


Characterization of a PBAT Degradation Carboxylesterase from Thermobacillus composti KWC4
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
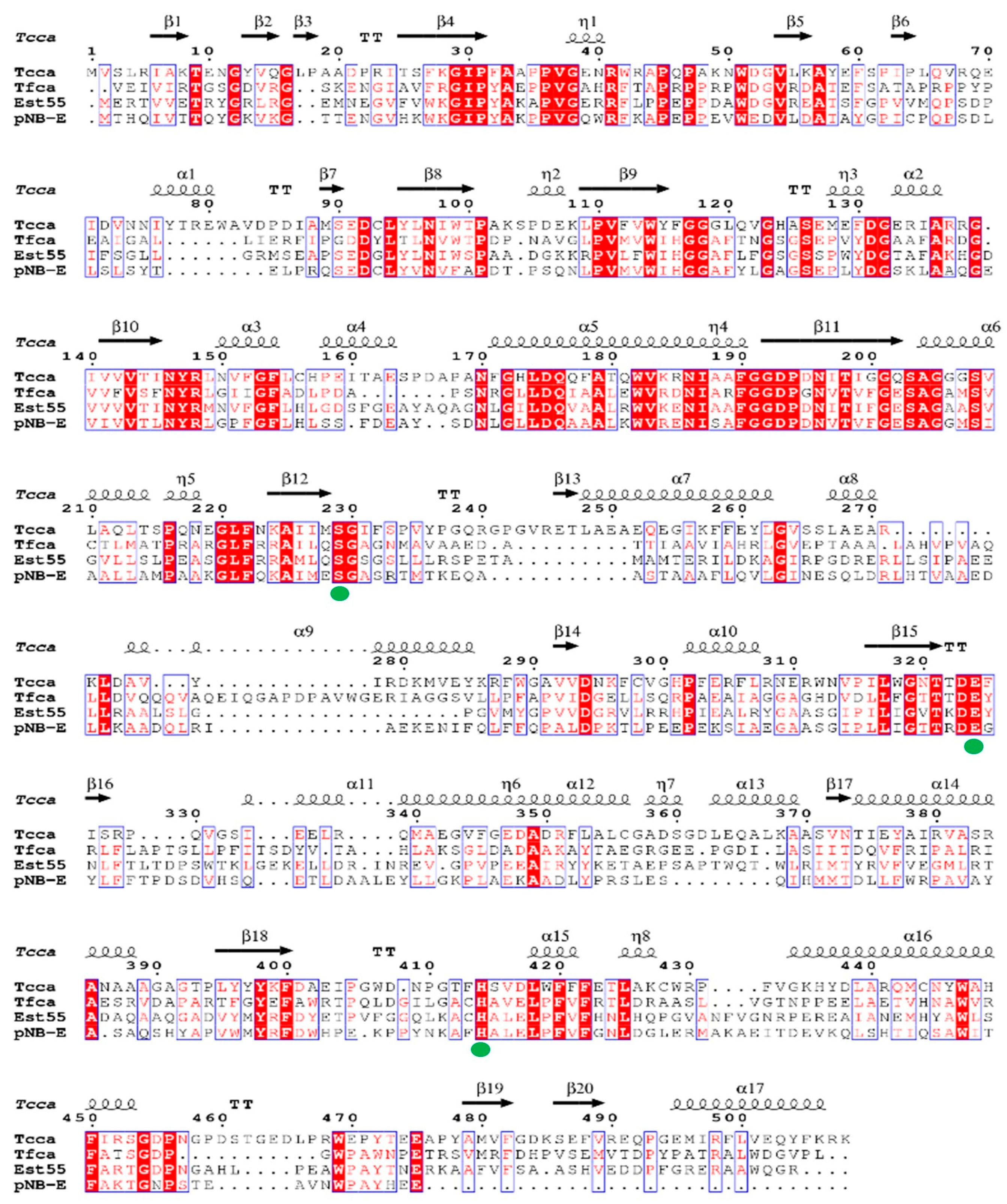
2.1. Protein Sequence Analysis
2.2. Characterization of Tcca
2.3. The Hydrolysis of PBAT Film by Tcca
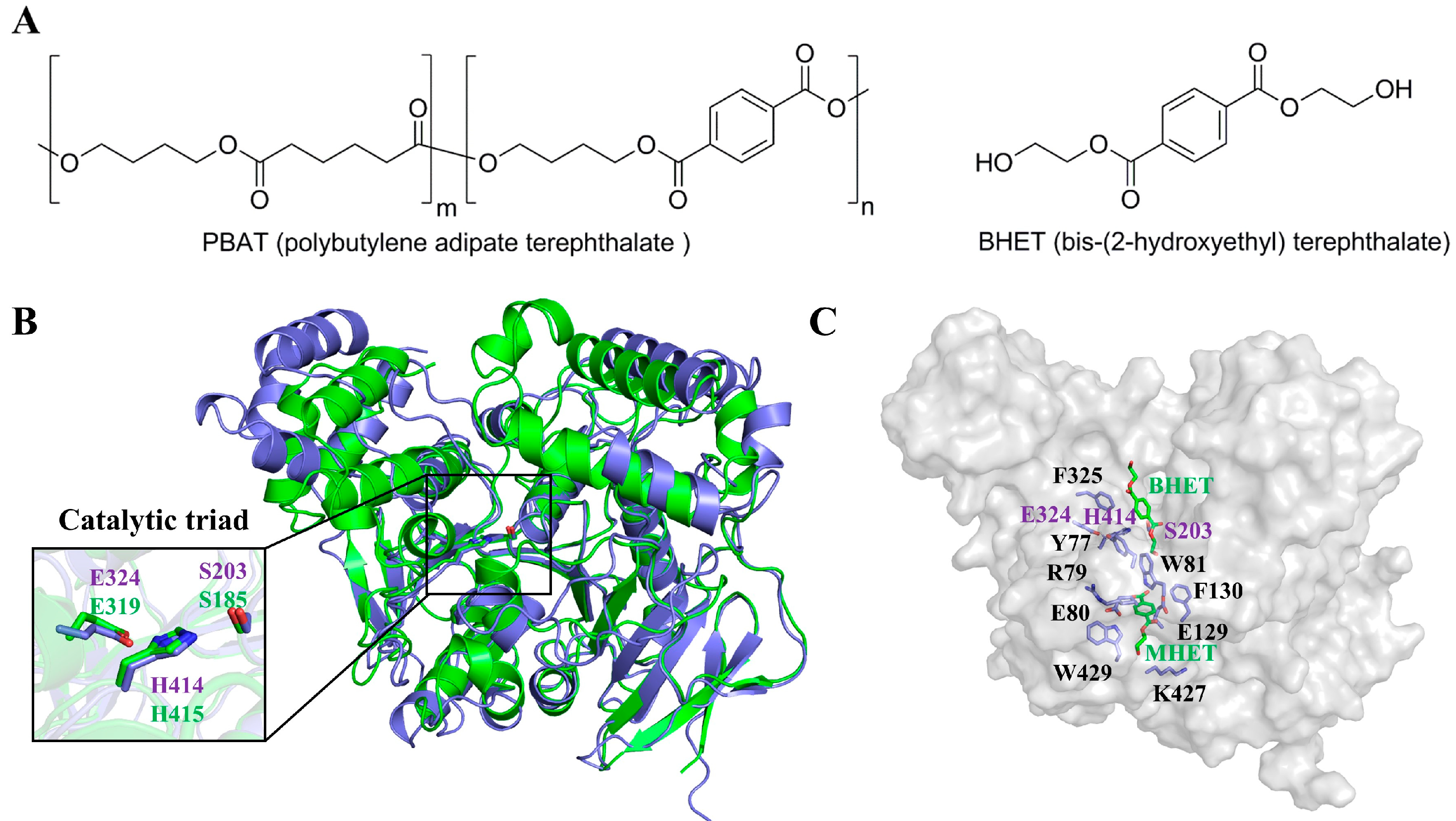
2.4. Tcca Structure Modeling and Possible Substrate Binding Cavity
2.5. Protein Engineering of Tcca
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cloning and Site−Directed Mutagenesis
4.3. Protein Purification
4.4. Characterization of Melting Temperature, Optimal pH and Temperature of Tcca
4.5. Scanning Electron Microscopy
4.6. Hydrolysis Activity of PBAT and BHET by Tcca and Its Mutants
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Witt, U.; Yamamoto, M.; Seeliger, U.; Muller, R.J.; Warzelhan, V. Biodegradable polymeric materials–not the origin but the chemical structure determines biodegradability. Angew. Chem. Int. Ed. 1999, 38, 1438–1442. [Google Scholar] [CrossRef]
- Gupta, A.; Chudasama, B.; Chang, B.P.; Mekonnen, T. Robust and sustainable PBAT–Hemp residue biocomposites: Reactive extrusion compatibilization and fabrication. Compos. Sci. Technol. 2021, 215, 109014. [Google Scholar] [CrossRef]
- Jiang, L.; Wolcott, M.P.; Zhang, J. Study of biodegradable Polylactide/Poly(butylene adipate–co–terephthalate) blends. Biomacromolecules 2006, 7, 199–207. [Google Scholar] [CrossRef]
- Pinheiro, I.F.; Ferreira, F.V.; Souza, D.H.S.; Gouveia, R.F.; Lona, L.M.F.; Morales, A.R.; Mei, L.H.I. Mechanical, rheological and degradation properties of PBAT nanocomposites reinforced by functionalized cellulose nanocrystals. Eur. Polym. J. 2017, 97, 356–365. [Google Scholar] [CrossRef]
- Pavon, C.; Aldas, M.; de la Rosa–Ramirez, H.; Lopez–Martinez, J.; Arrieta, M.P. Improvement of PBAT processability and mechanical performance by blending with pine resin derivatives for injection moulding rigid packaging with enhanced hydrophobicity. Polymers 2020, 12, 2891. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.K.; Wu, F.; Misra, M.; Mohanty, A.K. Reactive extrusion of sustainable PHBV/PBAT–based nanocomposite films with organically modified nanoclay for packaging applications: Compression moulding vs. cast film extrusion. Compos. Part B Eng. 2020, 198, 108141. [Google Scholar] [CrossRef]
- Pagno, V.; Modenes, A.N.; Dragunski, D.C.; Fiorentin–Ferrari, L.D.; Caetano, J.; Guellis, C.; Goncalves, B.C.; dos Anjos, E.V.; Pagno, F.; Martinelli, V. Heat treatment of polymeric PBAT/PCL membranes containing activated carbon from Brazil nutshell biomass obtained by electrospinning and applied in drug removal. J. Environ. Chem. Eng. 2020, 8, 104159. [Google Scholar] [CrossRef]
- Oguz, H.; Dogan, C.; Kara, D.; Ozen, Z.T.; Ovali, D.; Nofar, M. Development of PLA–PBAT and PLA–PBSA bio–blends: Effects of processing type and PLA crystallinity on morphology and mechanical properties. In Proceedings of the Europe/Africa Conference Dresden 2017—Polymer Processing Society PPS, Online, 27–29 June 2019; Volume 2055. [Google Scholar] [CrossRef]
- Nunes, F.C.; Ribeiro, K.C.; Martini, F.A.; Barrioni, B.R.; Santos, J.P.F.; Carvalho, B.M. PBAT/PLA/cellulose nanocrystals biocomposites compatibilized with polyethylene grafted maleic anhydride (PE–g–MA). J. Appl. Polym. Sci. 2021, 138, 51342. [Google Scholar] [CrossRef]
- Nobile, M.R.; Crocitti, A.; Malinconico, M.; Santagata, G.; Cerruti, P. Preparation and characterization of polybutylene succinate (PBS) and polybutylene adipate–terephthalate (PBAT) biodegradable blends. In Proceedings of the 9th International Conference on Times of Polymers and Composites: From Aerospace to Nanotechnology, Ischia, Italy, 17–21 June 2018; Volume 1981. [Google Scholar] [CrossRef]
- Pietrosanto, A.; Scarfato, P.; Di Maio, L.; Incarnato, L. Development of eco–sustainable PBAT–based blown films and performance analysis for food packaging applications. Materials 2020, 13, 5395. [Google Scholar] [CrossRef]
- Gambarini, V.; Pantos, O.; Kingsbury, J.M.; Weaver, L.; Handley, K.M.; Lear, G. Phylogenetic distribution of plastic–degrading microorganisms. mSystems 2021, 6, e01112-20. [Google Scholar] [CrossRef]
- Kanwal, A.; Zhang, M.; Sharaf, F.; Li, C.T. Screening and characterization of novel lipase producing Bacillus species from agricultural soil with high hydrolytic activity against PBAT poly (butylene adipate co terephthalate) co–polyesters. Polym. Bull. 2022, 79, 10053–10076. [Google Scholar] [CrossRef]
- Kanwal, A.; Zhang, M.; Sharaf, F.; Li, C.T. Polymer pollution and its solutions with special emphasis on Poly (butylene adipate terephthalate (PBAT)). Polym. Bull. 2022, 79, 9303–9330. [Google Scholar] [CrossRef]
- Qi, X.; Ren, Y.; Wang, X. New advances in the biodegradation of Poly(lactic) acid. Int. Biodeterior. Biodegrad. 2017, 117, 215–223. [Google Scholar] [CrossRef]
- Marten, E.; Muller, R.J.; Deckwer, W.D. Studies on the enzymatic hydrolysis of polyesters. II. Aliphatic–aromatic copolyesters. Polym. Degrad. Stab. 2005, 88, 371–381. [Google Scholar] [CrossRef]
- Wei, R.; von Haugwitz, G.; Pfaff, L.; Mican, J.; Badenhorst, C.P.S.; Liu, W.; Weber, G.; Austin, H.P.; Bednar, D.; Damborsky, J.; et al. Mechanism–based design of dfficient PET hydrolases. ACS Catal. 2022, 12, 3382–3396. [Google Scholar] [CrossRef]
- Tokiwa, Y.; Suzuki, T. Hydrolysis of polyesters by lipases. Nature 1977, 270, 76–78. [Google Scholar] [CrossRef]
- Herrero Acero, E.; Ribitsch, D.; Steinkellner, G.; Gruber, K.; Greimel, K.; Eiteljoerg, I.; Trotscha, E.; Wei, R.; Zimmermann, W.; Zinn, M.; et al. Enzymatic Surface Hydrolysis of PET: Effect of Structural Diversity on Kinetic Properties of Cutinases from Thermobifida. Macromolecules 2011, 44, 4632–4640. [Google Scholar] [CrossRef]
- Perz, V.; Baumschlager, A.; Bleymaier, K.; Zitzenbacher, S.; Hromic, A.; Steinkellner, G.; Pairitsch, A.; Lyskowski, A.; Gruber, K.; Sinkel, C.; et al. Hydrolysis of synthetic polyesters by Clostridium botulinum esterases. Biotechnol. Bioeng. 2016, 113, 1024–1034. [Google Scholar] [CrossRef]
- Wallace, P.W.; Haernvall, K.; Ribitsch, D.; Zitzenbacher, S.; Schittmayer, M.; Steinkellner, G.; Gruber, K.; Guebitz, G.M.; Birner–Gruenberger, R. PpEst is a novel PBAT degrading polyesterase identified by proteomic screening of Pseudomonas pseudoalcaligenes. Appl. Microbiol. Biotechnol. 2017, 101, 2291–2303. [Google Scholar] [CrossRef]
- Kitadokoro, K.; Thumarat, U.; Nakamura, R.; Nishimura, K.; Karatani, H.; Suzuki, H.; Kawai, F. Crystal structure of cutinase Est119 from Thermobifida alba AHK119 that can degrade modified polyethylene terephthalate at 1.76Å resolution. Polym. Degrad. Stab. 2012, 97, 771–775. [Google Scholar] [CrossRef]
- Perz, V.; Bleymaier, K.; Sinkel, C.; Kueper, U.; Bonnekessel, M.; Ribitsch, D.; Guebitz, G.M. Substrate specificities of cutinases on aliphatic–aromatic polyesters and on their model substrates. New Biotechnol. 2016, 33, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Kukolya, J. Thermobifida cellulolytica sp. nov., a novel lignocellulose–decomposing actinomycete. Int. J. Syst. Evol. Microbiol. 2002, 52, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Biundo, A.; Hromic, A.; Pavkov–Keller, T.; Gruber, K.; Quartinello, F.; Haernvall, K.; Perz, V.; Arrell, M.S.; Zinn, M.; Ribitsch, D.; et al. Characterization of a poly(butylene adipate–co–terephthalate)–hydrolyzing lipase from Pelosinus fermentans. Appl. Microbiol. Biotechnol. 2016, 100, 1753–1764. [Google Scholar] [CrossRef]
- Kleeberg, I.; Welzel, K.; VandenHeuvel, J.; Müller, R.J.; Deckwer, W.D. Characterization of a new extracellular hydrolase from Thermobifida fusca degrading aliphatic–aromatic copolyesters. Biomacromolecules 2005, 6, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Meyer Cifuentes, I.E.; Wu, P.; Zhao, Y.; Liu, W.; Neumann–Schaal, M.; Pfaff, L.; Barys, J.; Li, Z.; Gao, J.; Han, X.; et al. Molecular and biochemical differences of the tandem and cold–adapted PET hydrolases Ple628 and Ple629, isolated from a marine microbial consortium. Front. Bioeng. Biotechnol. 2022, 10, 930140. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, Y.; Wu, P.; Wang, H.; Li, Q.; Gao, J.; Qin, H.M.; Wei, H.; Bornscheuer, U.T.; Han, X.; et al. Structural insight and engineering of a plastic degrading hydrolase Ple629. Biochem. Biophys. Res. Commun. 2022, 626, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Satti, S.M.; Shah, A.A. Polyester–based biodegradable plastics: An approach towards sustainable development. Lett. Appl. Microbiol. 2020, 70, 413–430. [Google Scholar] [CrossRef]
- Jochens, H.; Aerts, D.; Bornscheuer, U.T. Thermostabilization of an esterase by alignment–guided focussed directed evolution. Protein Eng. Des. Sel. 2010, 23, 903–909. [Google Scholar] [CrossRef] [Green Version]
- Carr, P.D.; Ollis, D.L. Alpha/beta hydrolase fold: An update. Protein Pept. Lett. 2009, 16, 1137–1148. [Google Scholar] [CrossRef]
- von Haugwitz, G.; Han, X.; Pfaff, L.; Li, Q.; Wei, H.; Gao, J.; Methling, K.; Ao, Y.; Brack, Y.; Mican, J.; et al. Structural insights into (Tere)phthalate–ester hydrolysis by a carboxylesterase and its role in promoting PET depolymerization. ACS Catal. 2022, 12, 15259–15270. [Google Scholar] [CrossRef]
- Liu, P.; Ewis, H.E.; Tai, P.C.; Lu, C.D.; Weber, I.T. Crystal structure of the Geobacillus stearothermophilus carboxylesterase Est55 and its activation of prodrug CPT–11. J. Mol. Biol. 2007, 367, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Spiller, B.; Gershenson, A.; Arnold, F.H.; Stevens, R.C. A structural view of evolutionary divergence. Proc. Natl. Acad. Sci. USA 1999, 96, 12305–12310. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Saini, P.; Grewall, A.; Hooda, S. In silico approach for identification of polyethylene terephthalate hydrolase (PETase)–like enzymes. Bioremediat. J. 2022. [Google Scholar] [CrossRef]
- Denesyuk, A.; Dimitriou, P.S.; Johnson, M.S.; Nakayama, T.; Denessiouk, K. The acid–base–nucleophile catalytic triad in ABH–fold enzymes is coordinated by a set of structural elements. PLoS ONE 2020, 15, e0229376. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, S.; Yamato, S.; Kanaya, E.; Kim, J.J.; Koga, Y.; Takano, K.; Kanaya, S. Isolation of a novel cutinase homolog with polyethylene terephthalate–degrading activity from leaf–branch compost by using a metagenomic approach. Appl. Environ. Microbiol. 2012, 78, 1556–1562. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, L.; Gao, J.; Li, Z.; Jäckering, A.; Weber, G.; Mican, J.; Chen, Y.; Dong, W.; Han, X.; Feiler, C.G.; et al. Multiple substrate binding mode–guided engineering of a thermophilic PET hydrolase. ACS Catal. 2022, 12, 9790–9800. [Google Scholar] [CrossRef]
- Tarazona, N.A.; Wei, R.; Brott, S.; Pfaff, L.; Bornscheuer, U.T.; Lendlein, A.; Machatschek, R. Rapid depolymerization of poly(ethylene terephthalate) thin films by a dual–enzyme system and its impact on material properties. Chem. Catal. 2022, 2, 3573–3589. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue Substitutions | Sequence (5′–3′) |
|---|---|
| Y77W | ATTGATGTTAATAATATT tgg ACCCGCGAATGGGCCGTG |
| R79A | GTTAATAATATTTATACCgctGAATGGGCCGTGGACCCG |
| E80A | AATAATATTTATACCCGCgctTGGGCCGTGGACCCGGAT |
| W81A | AATATTTATACCCGCGAAgctGCCGTGGACCCGGATATT |
| E129A | CAGGTGGGTCATGCAAGTgctATGGAATTTGATGGCGAA |
| F130A | CATGCAAGTGAAATGGAAgctGATGGCGAACGCATTGCA |
| F325A | GGTAATACCACCGATGAAgctATTAGTCGTCCGCAGGTT |
| K427A | TTTTTTGAAACCTTAGCCgctTGTTGGCGCCCGTTTGTG |
| W429A | GAAACCTTAGCCAAATGTgctCGCCCGTTTGTGGGCAAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.; Li, Z.; Gao, J.; Zhao, Y.; Wang, H.; Qin, H.; Gu, Q.; Wei, R.; Liu, W.; Han, X. Characterization of a PBAT Degradation Carboxylesterase from Thermobacillus composti KWC4. Catalysts 2023, 13, 340. https://doi.org/10.3390/catal13020340
Wu P, Li Z, Gao J, Zhao Y, Wang H, Qin H, Gu Q, Wei R, Liu W, Han X. Characterization of a PBAT Degradation Carboxylesterase from Thermobacillus composti KWC4. Catalysts. 2023; 13(2):340. https://doi.org/10.3390/catal13020340
Chicago/Turabian StyleWu, Pan, Zhishuai Li, Jian Gao, Yipei Zhao, Hao Wang, Huimin Qin, Qun Gu, Ren Wei, Weidong Liu, and Xu Han. 2023. "Characterization of a PBAT Degradation Carboxylesterase from Thermobacillus composti KWC4" Catalysts 13, no. 2: 340. https://doi.org/10.3390/catal13020340
APA StyleWu, P., Li, Z., Gao, J., Zhao, Y., Wang, H., Qin, H., Gu, Q., Wei, R., Liu, W., & Han, X. (2023). Characterization of a PBAT Degradation Carboxylesterase from Thermobacillus composti KWC4. Catalysts, 13(2), 340. https://doi.org/10.3390/catal13020340