
β-Sitosterol Oleate Synthesis by Candida rugosa Lipase in a Solvent-Free Mini Reactor System: Free and Immobilized on Chitosan-Alginate Beads

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results and Discussion
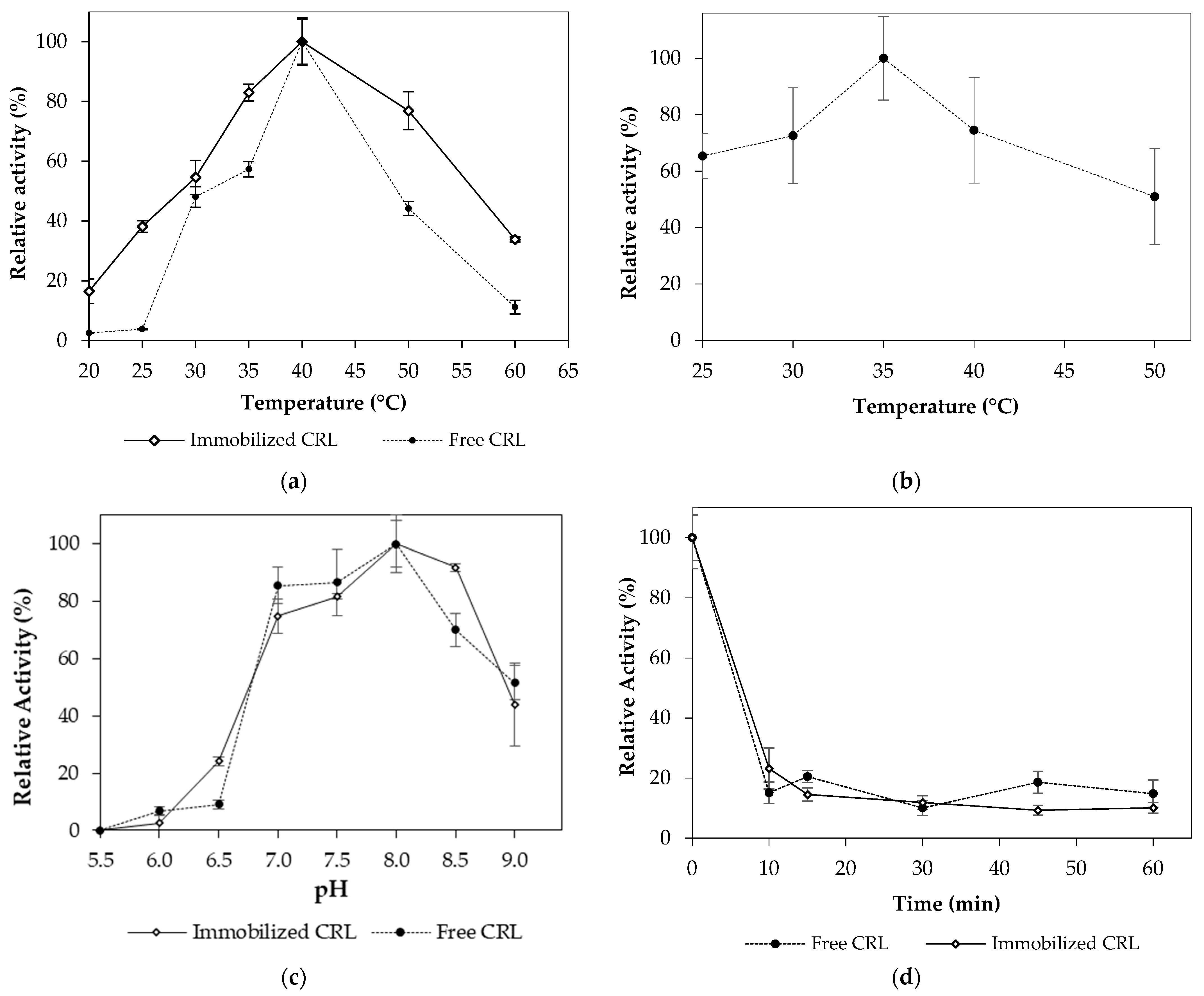
2.1. Biochemical Characterization
2.1.1. Effect of Temperature and pH on C. rugosa Lipase Activity
2.1.2. Kinetic Parameters
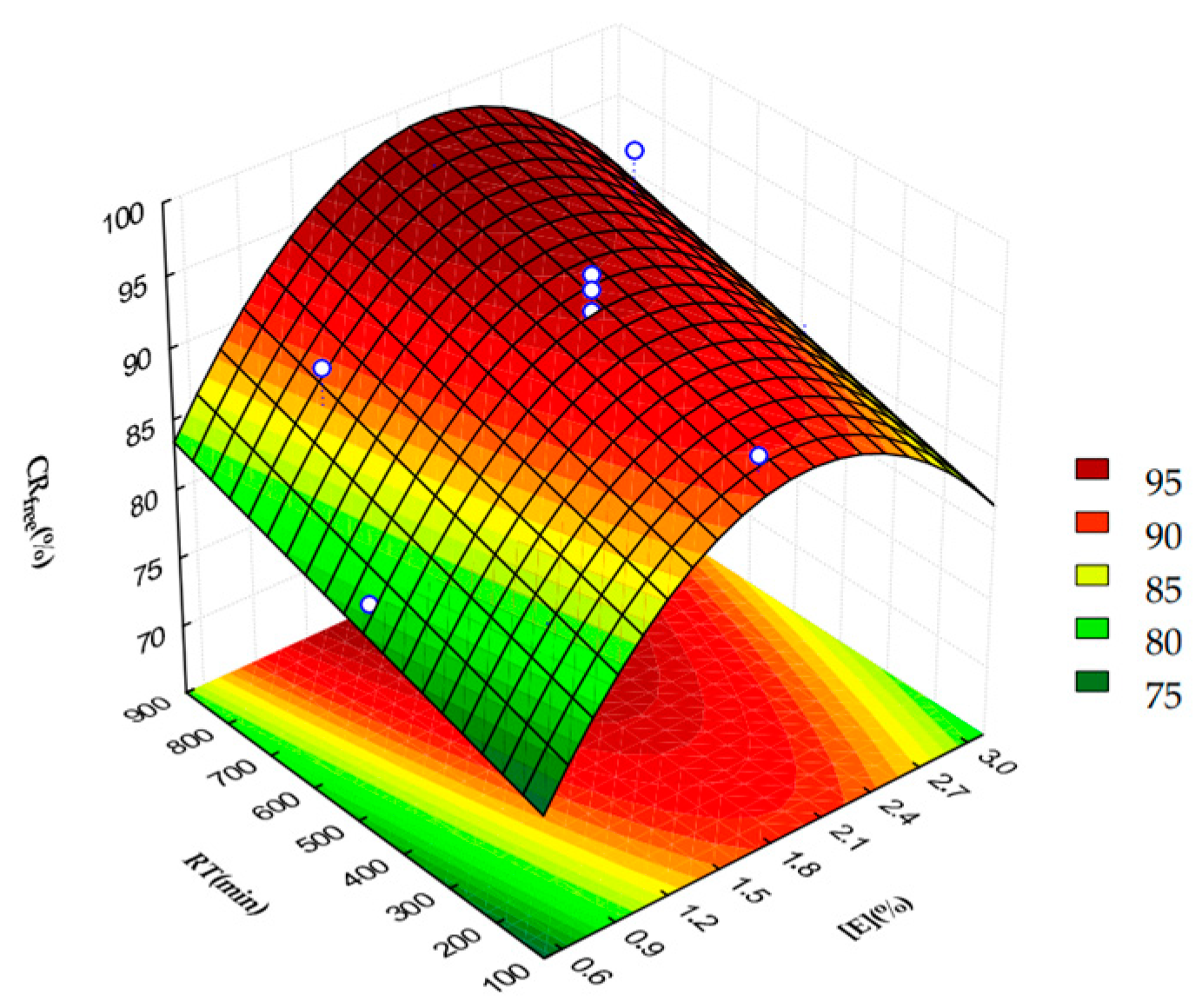
2.2. β-Sitosterol Oleate Production Using Free and Immobilized C. rugosa Lipase (CRL)
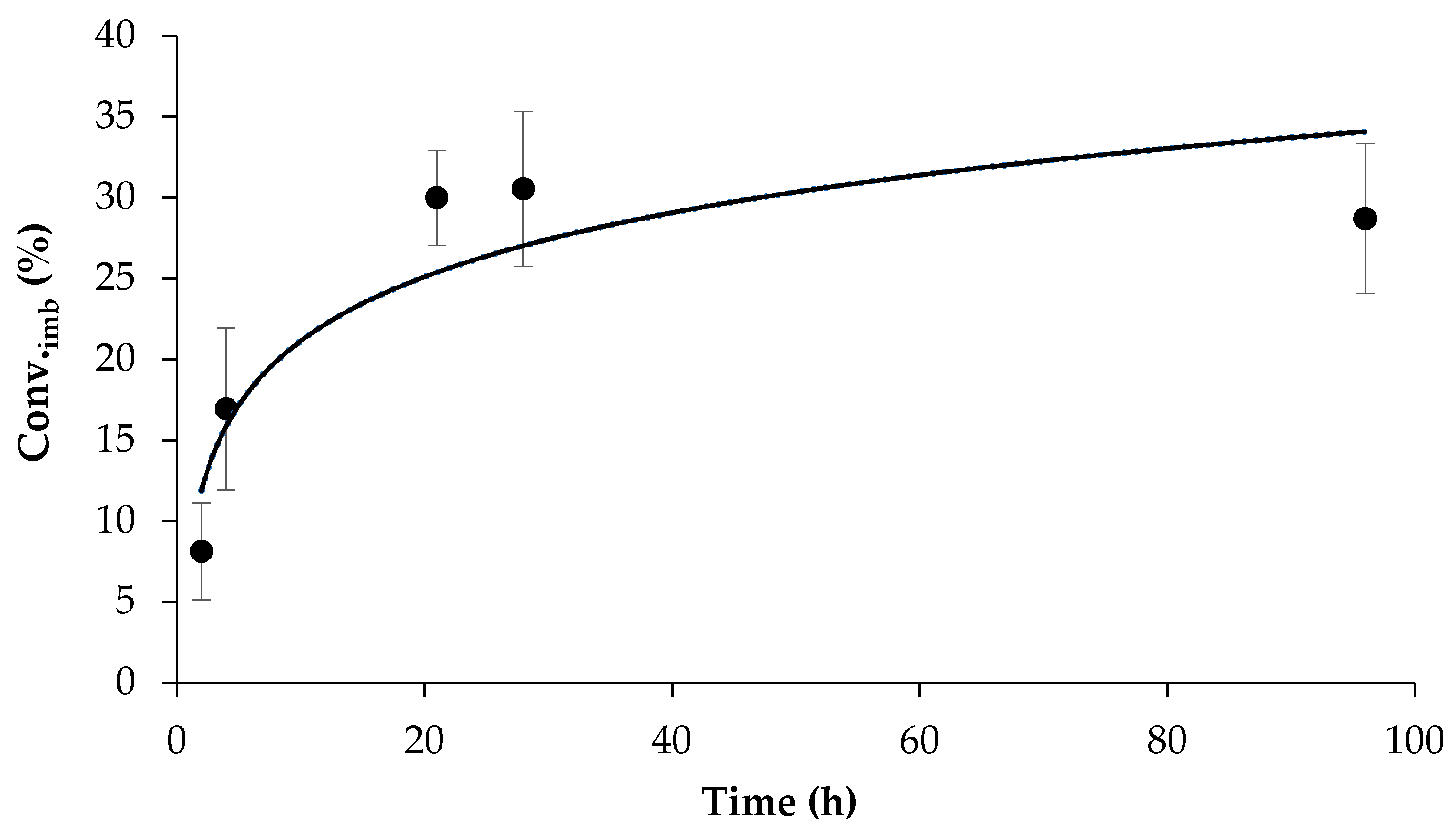
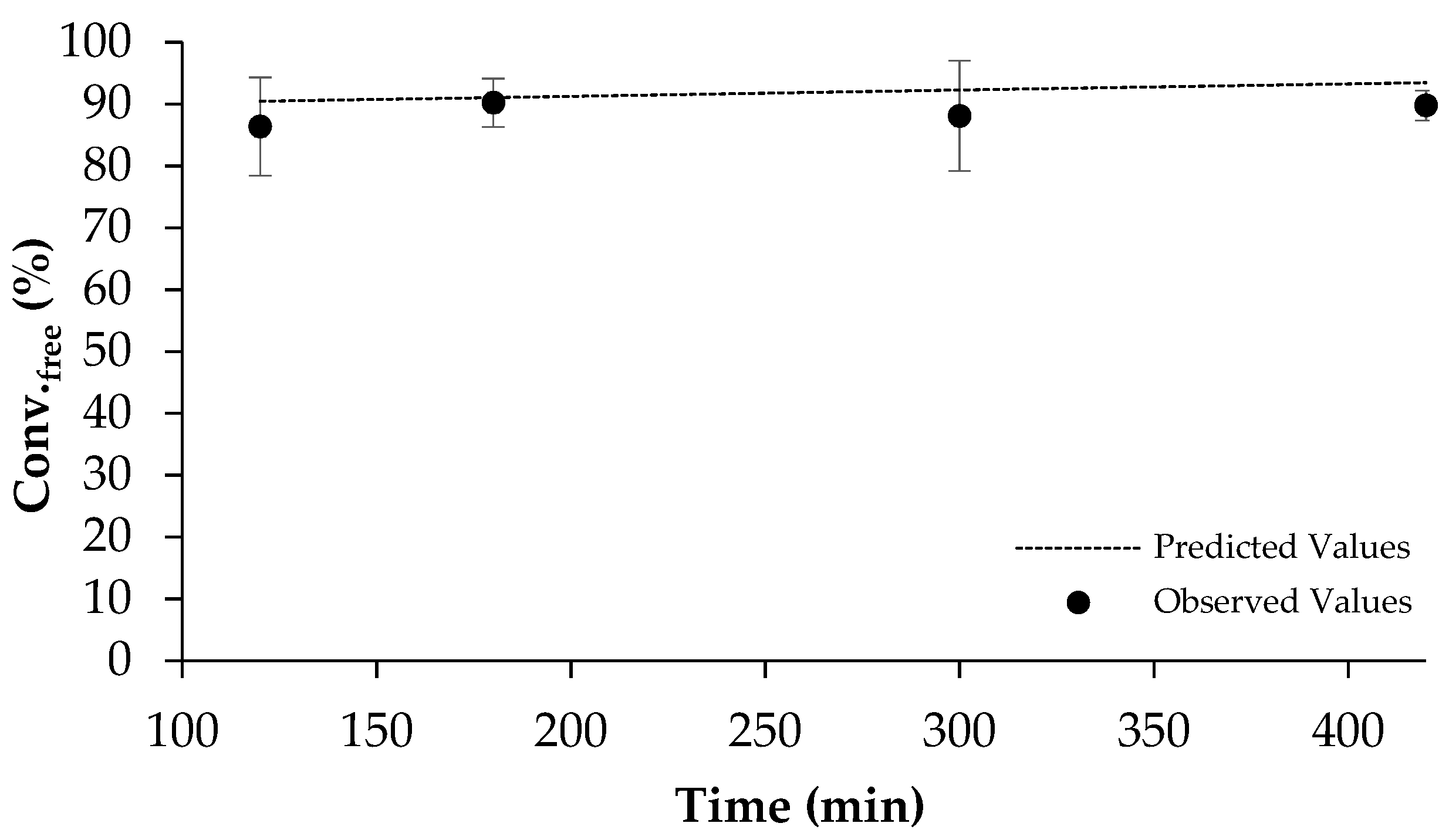
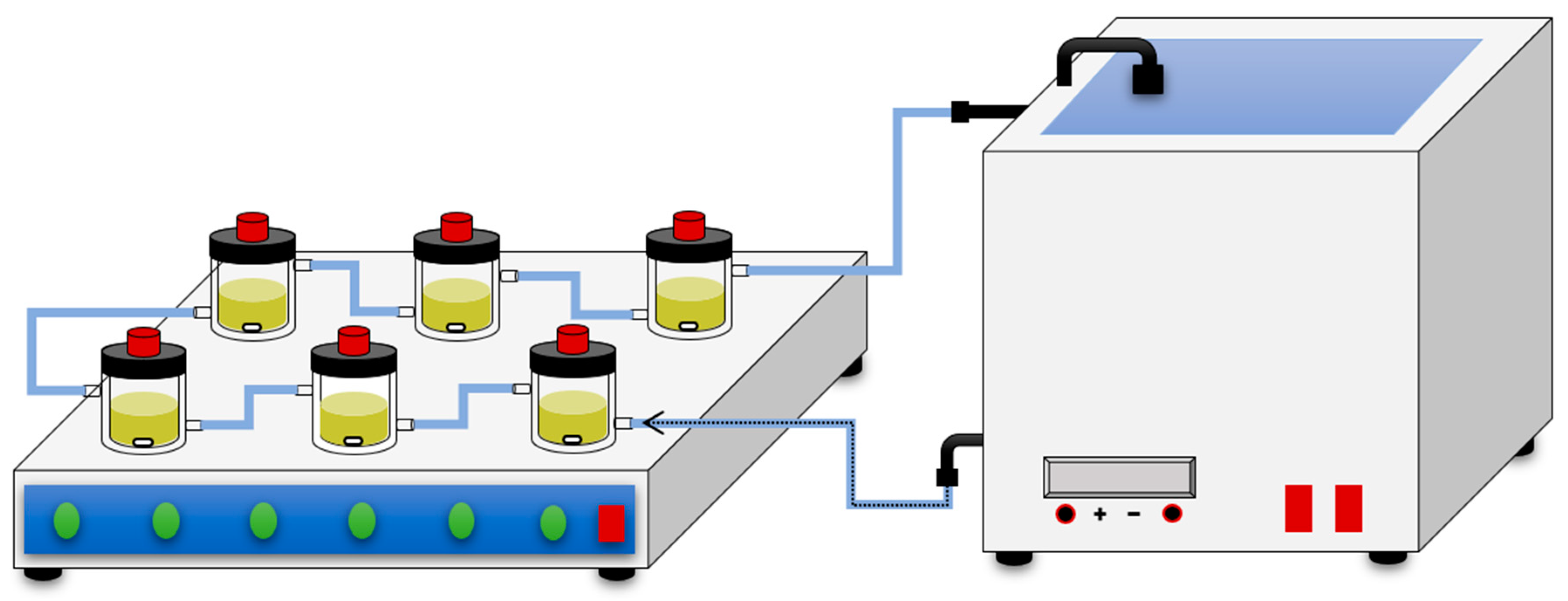
2.3. Scale-Up of β-Sitosterol Oleate Production
3. Materials and Methods
3.1. Materials
3.2. Immobilization of C. rugosa Lipase (CRL) in Chitosan-Alginate Beads
3.3. Biochemical Characterization
3.3.1. Effect of Temperature and pH on C. rugosa Lipase (CRL) Activity
3.3.2. Kinetic Parameters
3.4. β-Sitosterol Oleate Production Using Free and Immobilized C. rugosa Lipase (CRL)
3.4.1. Fractional Factorial Design (FFD)
3.4.2. Central Composite Rotational Design (CCRD)
3.5. Scale-Up of β-Sitosterol Oleate Production
3.6. Analytical Procedures
3.6.1. Hydrolytic Activity
3.6.2. Synthetic Activity
3.6.3. β-Sitosterol Concentration by Gas Chromatography (GC)
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brufau, G.; Canela, M.A.; Rafecas, M. Phytosterols: Physiologic and metabolic aspects related to cholesterol-lowering properties. Nutr. Res. 2008, 28, 217–225. [Google Scholar] [CrossRef]
- Plat, J.; Baumgartner, S.; Vanmierlo, T.; Lütjohann, D.; Calkins, K.L.; Burrin, D.G.; Guthrie, G.; Thijs, C.; Te Velde, A.A.; ACE, V.; et al. Plant-based sterols and stanols in health & disease: “Consequences of human development in a plant-based environment”? Prog. Lipid Res. 2019, 74, 87–102. [Google Scholar]
- Yang, F.; Oyeyinka, S.A.; Ma, Y. Novel Synthesis of Phytosterol Ester from Soybean Sterol and Acetic Anhydride. J. Food Sci. 2016, 81, C1629–C1635. [Google Scholar] [CrossRef]
- Mussner, M.J.; Parhofer, K.G.; Von Bergmann, K.; Schwandt, P.; Broedl, U.; Otto, C. Effects of phytosterol ester-enriched margarine on plasma lipoproteins in mild to moderate hypercholesterolemia are related to basal cholesterol and fat intake. Metabolism 2002, 51, 189–194. [Google Scholar] [CrossRef]
- Davidson, M.H.; Maki, K.C.; Umporowicz, D.M.; Ingram, K.A.; Dicklin, M.R.; Lane, R.W.; Franke, W.C.; Robins, S.J.; Schaefer, E.; McNamara, J.R.; et al. Safety and tolerability of esterified phytosterols administered in reduced-fat spread and salad dressing to healthy adult men and women. J. Am. Coll. Nutr. 2001, 20, 307–319. [Google Scholar] [CrossRef]
- Kwak, H.S.; Ahn, H.J.; Ahn, J. Development of phytosterol ester-added Cheddar cheese for lowering blood cholesterol. Asian-Australas. J. Anim. Sci. 2005, 18, 267–276. [Google Scholar] [CrossRef]
- Pouilloux, Y.; Courtois, G.; Boisseau, M.; Piccirilli, A.; Barrault, J. Solid base catalysts for the synthesis of phytosterol esters. Green Chem. 2003, 5, 89–91. [Google Scholar] [CrossRef]
- Chang, M.; Zhang, T.; Feng, W.; Wang, T.; Liu, R.; Jin, Q.; Wang, X. Preparation of highly pure stigmasteryl oleate by enzymatic esterification of stigmasterol enriched from soybean phytosterols. LWT 2020, 128, 109464. [Google Scholar] [CrossRef]
- Brígida, A.I.S.; Amaral, P.F.F.; Coelho, M.A.Z.; Gonçalves, L.R.B. Lipase from Yarrowia lipolytica: Production, characterization and application as an industrial biocatalyst. J. Mol. Catal. B Enzym. 2014, 101, 148–158. [Google Scholar] [CrossRef]
- Treichel, H.; de Oliveira, D.; Mazutti, M.A.; Di Luccio, M.; Oliveira, J.V. A review on microbial lipases production. Food Bioprocess Technol. 2010, 3, 182–196. [Google Scholar] [CrossRef]
- Pereira, A.d.S.; de Souza, A.H.; Fraga, J.L.; Villeneuve, P.; Torres, A.G.; Amaral, P.F.F. Lipases as Effective Green Biocatalysts for Phytosterol Esters’ Production: A Review. Catalysts 2022, 12, 88. [Google Scholar] [CrossRef]
- Domínguez De María, P.; Sánchez-Montero, J.M.; Sinisterra, J.V.; Alcántara, A.R. Understanding Candida rugosa lipases: An overview. Biotechnol. Adv. 2006, 24, 180–196. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ogino, A.; Miyake, Y.; Mori, H.; Hosoda, A.; Fujita, M.; Tsuno, T.; Adachi, S. Lipase-catalyzed esterification of triterpene alcohols and phytosterols with oleic acid. JAOCS J. Am. Oil Chem. Soc. 2014, 91, 1885–1890. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef]
- dos Santos, M.M.O.; de Menezes, L.H.S.; do Espirito Santo, E.L.; de Carvalho, M.S.; Gonçalves, M.S.; de Carvalho Tavares, I.M.; Mendes, A.A.; Ruiz, H.A.; Salay, L.C.; Franco, M.; et al. Synthesis of hexyl butyrate (apple and citrus aroma) by Candida rugosa lipase immobilized on Diaion HP-20 using the Box-Behnken design. Food Sci. Biotechnol. 2022, 32, 689–696. [Google Scholar] [CrossRef]
- Mendonca Ferreira, R.D.; Brackmann, R.; Pereira, E.B.; Costa da Rocha, R.D. Immobilization of lipase from Candida rugosa onto niobium oxide. Biocatal. Agric. Biotechnol. 2020, 30, 101812. [Google Scholar] [CrossRef]
- Jiang, Z.; Yu, M.; Ren, L.; Zhou, H.; Wei, P. Synthesis of phytosterol esters catalyzed by immobilized lipase in organic media. Cuihua Xuebao/Chin. J. Catal. 2013, 34, 2255–2262. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Li, Y.; Wang, X.; Peng, X.; Zhu, W. Preparation of superparamagnetic Fe3O4@alginate/chitosan nanospheres for Candida rugosa lipase immobilization and utilization of layer-by-layer assembly to enhance the stability of immobilized lipase. ACS Appl. Mater. Interfaces 2012, 4, 5169–5178. [Google Scholar] [CrossRef]
- Zheng, M.M.; Lu, Y.; Dong, L.; Guo, P.M.; Deng, Q.C.; Li, W.L.; Feng, Y.Q.; Huang, F.H. Immobilization of Candida rugosa lipase on hydrophobic/strong cation-exchange functional silica particles for biocatalytic synthesis of phytosterol esters. Bioresour. Technol. 2012, 115, 141–146. [Google Scholar] [CrossRef]
- Betigeri, S.S.; Neau, S.H. Immobilization of lipase using hydrophilic polymers in the form of hydrogel beads. Biomaterials 2002, 23, 3627–3636. [Google Scholar] [CrossRef]
- Alsarra, I.A.; Neau, S.H.; Howard, M.A. Effects of preparative parameters on the properties of chitosan hydrogel beads containing Candida rugosa lipase. Biomaterials 2004, 25, 2645–2655. [Google Scholar] [CrossRef]
- Pereira, A.d.S.; Souza, C.P.L.; Moraes, L.; Fontes-Sant’ana, G.C.; Amaral, P.F.F. Polymers as encapsulating agents and delivery vehicles of enzymes. Polymers 2021, 13, 4061. [Google Scholar] [CrossRef]
- Mohamad, N.R.; Marzuki, N.H.C.; Buang, N.A.; Huyop, F.; Wahab, R.A. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol. Biotechnol. Equip. 2015, 29, 205–220. [Google Scholar] [CrossRef]
- Pereira, A.d.S.; Fraga, J.L.; Diniz, M.M.; Fontes-Sant’ana, G.C.; Amaral, P.F.F. High catalytic activity of lipase from yarrowia lipolytica immobilized by microencapsulation. Int. J. Mol. Sci. 2018, 19, 3393. [Google Scholar] [CrossRef]
- Totaro, D.; Rothbauer, M.; Steiger, M.G.; Mayr, T.; Wang, H.Y.; Lin, Y.S.; Sauer, M.; Altvater, M.; Ertl, P.; Mattanovich, D. Downscaling screening cultures in a multifunctional bioreactor array-on-a-chip for speeding up optimization of yeast-based lactic acid bioproduction. Biotechnol. Bioeng. 2020, 117, 2046–2057. [Google Scholar] [CrossRef]
- Long, Q.; Liu, X.; Yang, Y.; Li, L.; Harvey, L.; McNeil, B.; Bai, Z. The development and application of high throughput cultivation technology in bioprocess development. J. Biotechnol. 2014, 192, 323–338. [Google Scholar] [CrossRef]
- Ben Akacha, N.; Gargouri, M. Microbial and enzymatic technologies used for the production of natural aroma compounds: Synthesis, recovery modeling, and bioprocesses. Food Bioprod. Process. 2015, 94, 675–706. [Google Scholar] [CrossRef]
- Patel, V.; Gajera, H.; Gupta, A.; Manocha, L.; Madamwar, D. Synthesis of ethyl caprylate in organic media using Candida rugosa lipase immobilized on exfoliated graphene oxide: Process parameters and reusability studies. Biochem. Eng. J. 2015, 95, 62–70. [Google Scholar] [CrossRef]
- Wu, J.C.; Selvam, V.; Teo, H.H.; Chow, Y.; Talukder, M.M.R.; Choi, W.J. Immobilization of Candida rugosa lipase by cross-linking with glutaraldehyde followed by entrapment in alginate beads. Biocatal. Biotransform. 2006, 24, 352–357. [Google Scholar] [CrossRef]
- Ozyilmaz, E.; Sayin, S. Utilization of Catalytic Properties of the Encapsulated Lipase with Calix[4]arene-Adorned Sporopollenin. Polycycl. Aromat. Compd. 2018, 38, 272–281. [Google Scholar] [CrossRef]
- Won, K.; Kim, S.; Kim, K.J.; Park, H.W.; Moon, S.J. Optimization of lipase entrapment in Ca-alginate gel beads. Process Biochem. 2005, 40, 2149–2154. [Google Scholar] [CrossRef]
- Pereira, A.d.S.; Diniz, M.M.; De Jong, G.; Gama Filho, H.S.; dos Anjos, M.J.; Finotelli, P.V.; Fontes-Sant’Ana, G.C.; Amaral, P.F.F. Chitosan-alginate beads as encapsulating agents for Yarrowia lipolytica lipase: Morphological, physico-chemical and kinetic characteristics. Int. J. Biol. Macromol. 2019, 139, 621–630. [Google Scholar] [CrossRef]
- Yuce-Dursun, B.; Cigil, A.B.; Dongez, D.; Kahraman, M.V.; Ogan, A.; Demir, S. Preparation and characterization of sol-gel hybrid coating films for covalent immobilization of lipase enzyme. J. Mol. Catal. B Enzym. 2016, 127, 18–25. [Google Scholar] [CrossRef]
- Ozyilmaz, G. The effect of spacer arm on hydrolytic and synthetic activity of Candida rugosa lipase immobilized on silica gel. J. Mol. Catal. B Enzym. 2009, 56, 231–236. [Google Scholar] [CrossRef]
- Paula, A.V.; Moreira, A.B.R.; Braga, L.P.; De Castro, H.F.; Bruno, L.M. Comparação do desempenho da lipase de candida rugosa imobilizada em suporte híbrido de polissiloxano-polivinilálcool empregando diferentes metodologias. Quim. Nov. 2008, 31, 35–40. [Google Scholar] [CrossRef]
- Chiou, S.H.; Wu, W.T. Immobilization of Candida rugosa lipase on chitosan with activation of the hydroxyl groups. Biomaterials 2004, 25, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Monajati, M.; Borandeh, S.; Hesami, A.; Mansouri, D.; Tamaddon, A.M. Immobilization of l-asparaginase on aspartic acid functionalized graphene oxide nanosheet: Enzyme kinetics and stability studies. Chem. Eng. J. 2018, 354, 1153–1163. [Google Scholar] [CrossRef]
- Brígida, A.I.S.; Pinheiro, Á.D.T.; Ferreira, A.L.O.; Gonçalves, L.R.B. Immobilization of Candida antarctica lipase B by adsorption to green coconut fiber. Appl. Biochem. Biotechnol. 2008, 146, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Elias, N.; Wahab, R.A.; Chandren, S.; Abdul Razak, F.I.; Jamalis, J. Effect of operative variables and kinetic study of butyl butyrate synthesis by Candida rugosa lipase activated by chitosan-reinforced nanocellulose derived from raw oil palm leaves. Enzym. Microb. Technol. 2019, 130, 109367. [Google Scholar] [CrossRef] [PubMed]
- Sá, A.G.A.; Meneses, A.C.d.; Araújo, P.H.H.d.; Oliveira, D.d. A review on enzymatic synthesis of aromatic esters used as flavor ingredients for food, cosmetics and pharmaceuticals industries. Trends Food Sci. Technol. 2017, 69, 95–105. [Google Scholar] [CrossRef]
- Cui, C.; Guan, N.; Xing, C.; Chen, B.; Tan, T. Immobilization of Yarrowia lipolytica lipase Ylip2 for the biocatalytic synthesis of phytosterol ester in a water activity controlled reactor. Colloids Surf. B Biointerfaces 2016, 146, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, J.; Huang, F.; Zheng, M. An efficient and robust continuous-flow bioreactor for the enzymatic preparation of phytosterol esters based on hollow lipase microarray. Food Chem. 2022, 372, 131256. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.M.; Santiago, F.L.B.; Silva, N.A.G.d.; Luiz, J.H.H.; Fernandéz-Lafuente, R.; Mendes, A.A.; Hirata, D.B. Different strategies to immobilize lipase from Geotrichum candidum: Kinetic and thermodynamic studies. Process Biochem. 2018, 67, 55–63. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Immobilized CRL | Free CRL | ||
|---|---|---|---|---|
| Hydrolysis | Synthesis | Hydrolysis | Synthesis | |
| Vmax (μmol/min/g) a | 2359.13 | - | 4121.84 | 2.24 |
| Km (μmol/L) b | 660.02 | - | 403.06 | 234.44 |
| Run | Real Values (Corresponding Coded Levels) | Conv.free (%) 7 | Conv.imb (%) 8 | |||||
|---|---|---|---|---|---|---|---|---|
| Molar Ratio (mol/mol) 1 | [Eimb] (%) 2 | [Efree] (%) 3,* | T (°C) 4 | RT (min) 5 | S (rpm) 6 | |||
| 1 | 8(−1) | 1(−1) | 0.3(−1) | 40(−1) | 30(−1) | 800(+1) | 25.14 ± 4.73 | 21.16 ± 5.60 |
| 2 | 12(+1) | 1(−1) | 0.3(−1) | 40(−1) | 30(−1) | 200(−1) | 29.75 ± 5.23 | 11.64 ± 4.40 |
| 3 | 8(−1) | 5(+1) | 1.5(+1) | 40(−1) | 30(−1) | 200(−1) | 24.40 ± 2.57 | 21.97 ± 6.99 |
| 4 | 12(+1) | 5(+1) | 1.5(+1) | 40(−1) | 30(−1) | 800(+1) | 33.06 ± 6.77 | 23.85 ± 6.93 |
| 5 | 8(−1) | 1(−1) | 0.3(−1) | 50(+1) | 30(−1) | 200(−1) | 17.35 ± 6.61 | 19.29 ± 5.75 |
| 6 | 12(+1) | 1(−1) | 0.3(−1) | 50(+1) | 30(−1) | 800(+1) | 21.17 ± 6.18 | 17.54 ± 6.30 |
| 7 | 8(−1) | 5(+1) | 1.5(+1) | 50(+1) | 30(−1) | 800(+1) | 21.38 ± 3.09 | 24.70 ± 0.91 |
| 8 | 12(+1) | 5(+1) | 1.5(+1) | 50(+1) | 30(−1) | 200(−1) | 24.87 ± 4.33 | 18.89 ± 1.24 |
| 9 | 8(−1) | 1(−1) | 0.3(−1) | 40(−1) | 360(+1) | 200(−1) | 30.91 ± 4.72 | 23.84 ± 6.70 |
| 10 | 12(+1) | 1(−1) | 0.3(−1) | 40(−1) | 360(+1) | 800(+1) | 23.01 ± 3.64 | 18.85 ± 5.20 |
| 11 | 8(−1) | 5(+1) | 1.5(+1) | 40(−1) | 360(+1) | 800(+1) | 82.13 ± 3.14 | 22.27 ± 5.70 |
| 12 | 12(+1) | 5(+1) | 1.5(+1) | 40(−1) | 360(+1) | 200(−1) | 84.01 ± 0.32 | 25.41 ± 5.51 |
| 13 | 8(−1) | 1(−1) | 0.3(−1) | 50(+1) | 360(+1) | 800(+1) | 40.54 ± 4.62 | 22.37 ± 2.60 |
| 14 | 12(+1) | 1(−1) | 0.3(−1) | 50(+1) | 360(+1) | 200(−1) | 36.97 ± 0.90 | 21.80 ± 7.70 |
| 15 | 8(−1) | 5(+1) | 1.5(+1) | 50(+1) | 360(+1) | 200(−1) | 83.42 ± 4.68 | 24.24 ± 2.29 |
| 16 | 12(+1) | 5(+1) | 1.5(+1) | 50(+1) | 360(+1) | 800(+1) | 74.25 ± 0.06 | 20.13 ± 3.80 |
| 17 (C 9) | 10(0) | 3(0) | 0.9(0) | 45(0) | 195(0) | 500(0) | 69.90 ± 1.41 | 26.35 ± 7.52 |
| 18 (C 9) | 10(0) | 3(0) | 0.9(0) | 45(0) | 195(0) | 500(0) | 59.49 ± 6.63 | 24.93 ± 6.64 |
| 19 (C 9) | 10(0) | 3(0) | 0.9(0) | 45(0) | 195(0) | 500(0) | 54.01 ± 5.34 | 24.91 ± 6.01 |
| Run | Real Values (Corresponding Coded Levels) | Conv.free(%) 3 | |
|---|---|---|---|
| [Efree] (%) 1 | RT (min) 2 | ||
| 1 | 0.9(−1) | 240.0(−1) | 80.99 ± 3.27 |
| 2 | 0.9(−1) | 720.0(+1) | 90.24 ± 6.53 |
| 3 | 2.7(+1) | 240.0(−1) | 86.87 ± 5.85 |
| 4 | 2.7(+1) | 720.0(+1) | 96.26 ± 0.40 |
| 5 | 0.54(−1.41) | 480.0(0) | 80.85 ± 2.42 |
| 6 | 3.06(+1.41) | 480.0(0) | 85.07 ± 6.96 |
| 7 | 1.8(0) | 140.6(−1.41) | 91.50 ± 2.09 |
| 8 | 1.8(0) | 819.4(+1.41) | 92.15 ± 4.90 |
| 9 (C 4) | 1.8(0) | 480.0(0) | 96.84 ± 0.14 |
| 10 (C 4) | 1.8(0) | 480.0(0) | 95.76 ± 0.33 |
| 11 (C 4) | 1.8(0) | 480.0(0) | 94.28 ± 1.22 |
| Factor | DF 1 | Sum of Square | Mean Square | F-Value 2 | p-Value | R2 |
|---|---|---|---|---|---|---|
| 0.8129 | ||||||
| (1) [E] (%) (L) | 1 | 39.9298 | 39.9298 | 24.1984 | 0.038928 | |
| [E] (%) (Q) | 1 | 191.3228 | 191.3228 | 115.9461 | 0.008515 | |
| (2) RT (min) (L) | 1 | 47.8341 | 47.8341 | 28.9886 | 0.032808 | |
| Lack of Fit | 5 | 61.0145 | 12.2029 | 7.3952 | 0.123389 | |
| Pure Error | 2 | 3.3002 | 1.6501 | |||
| Total SS | 10 | 343.4015 |
| Run | [E] (%) 1 | RT (min) 2 | Predicted Values | Observed Values Real |
|---|---|---|---|---|
| 1 | 0.60 | 200 | 78.20 | 69.13 ± 11.26 |
| 2 | 0.90 | 300 | 84.35 | 81.24 ± 4.34 |
| 3 | 1.20 | 400 | 89.24 | 92.56 ± 0.42 |
| 4 | 1.50 | 500 | 92.88 | 93.79 ± 1.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, A.d.S.; Fraga, J.L.; Souza, C.P.L.; Torres, A.G.; Amaral, P.F.F. β-Sitosterol Oleate Synthesis by Candida rugosa Lipase in a Solvent-Free Mini Reactor System: Free and Immobilized on Chitosan-Alginate Beads. Catalysts 2023, 13, 780. https://doi.org/10.3390/catal13040780
Pereira AdS, Fraga JL, Souza CPL, Torres AG, Amaral PFF. β-Sitosterol Oleate Synthesis by Candida rugosa Lipase in a Solvent-Free Mini Reactor System: Free and Immobilized on Chitosan-Alginate Beads. Catalysts. 2023; 13(4):780. https://doi.org/10.3390/catal13040780
Chicago/Turabian StylePereira, Adejanildo da S., Jully L. Fraga, Camila P. L. Souza, Alexandre G. Torres, and Priscilla F. F. Amaral. 2023. "β-Sitosterol Oleate Synthesis by Candida rugosa Lipase in a Solvent-Free Mini Reactor System: Free and Immobilized on Chitosan-Alginate Beads" Catalysts 13, no. 4: 780. https://doi.org/10.3390/catal13040780
APA StylePereira, A. d. S., Fraga, J. L., Souza, C. P. L., Torres, A. G., & Amaral, P. F. F. (2023). β-Sitosterol Oleate Synthesis by Candida rugosa Lipase in a Solvent-Free Mini Reactor System: Free and Immobilized on Chitosan-Alginate Beads. Catalysts, 13(4), 780. https://doi.org/10.3390/catal13040780