Cross-Linking with Polyethylenimine Confers Better Functional Characteristics to an Immobilized β-glucosidase from Exiguobacterium antarcticum B7

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Use of Different Poly-Functional Molecules and Increased Stability of EaBglA against Temperature
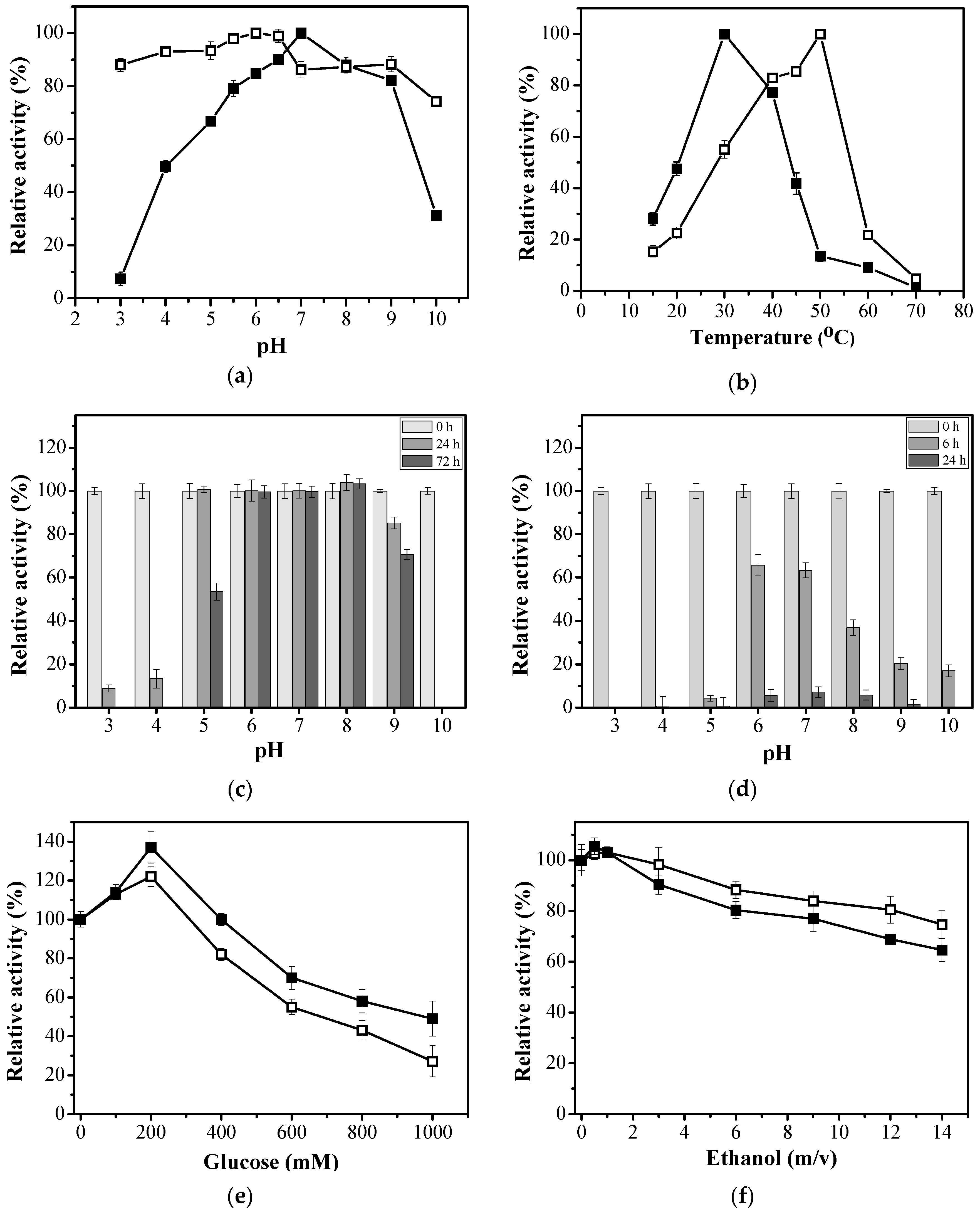
2.2. Effects of the Immobilization on the Functional Properties of EaBglA
2.3. Effect of Immobilization on the Resistance to Glucose and Ethanol
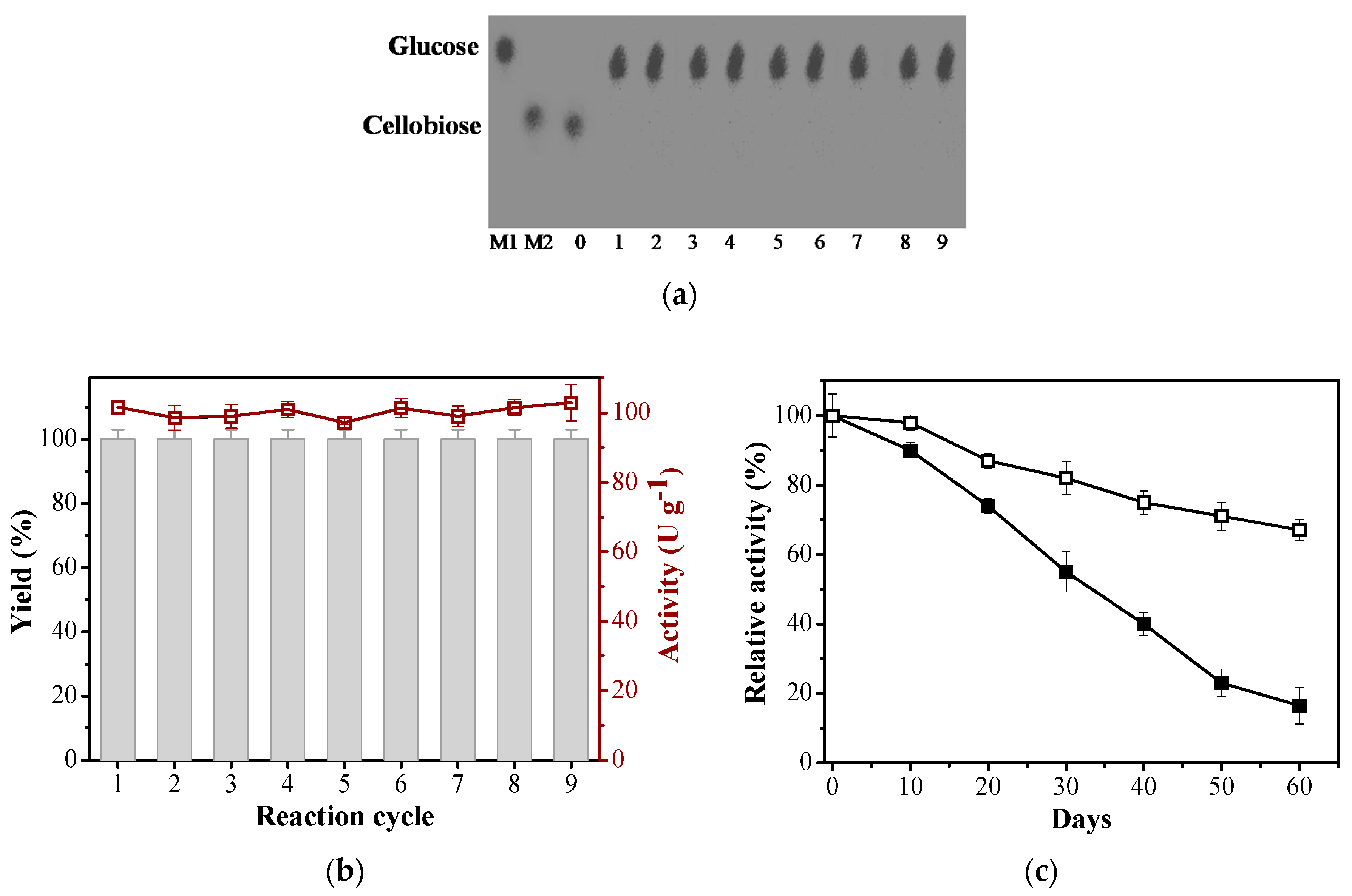
2.4. Conversion of Cellobiose to Glucose and Storage
3. Materials and Methods
3.1. Material
3.2. Cloning, Heterologous Expression and Purification Protein
3.3. Construction of Supports Applied in the EaBglA Immobilization
3.3.1. Activation of Agarose-Based Beads
3.3.2. Modification of Epoxy-Activated Agarose Beads
Support Activated with Glutaraldehyde Group
Anionic Support Activated with Glutaraldehyde Group and Metal Chelate
3.4. Immobilization of EaBglA on Activated Agarose Supports
3.5. Functional Assays
3.6. Operational Stability and Storage
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wang, S.; Dai, G.; Yang, H.; Luo, Z. Lignocellulosic biomass pyrolysis mechanism: A state-of-the-art review. Prog. Energy Combust. Sci. 2017, 62, 33–86. [Google Scholar] [CrossRef]
- Bhatia, Y.; Mishra, S.; Bisaria, V.S. Microbial β-glucosidases: Cloning, properties and applications. Crit. Rev. Biotechnol. 2002, 22, 375–407. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, A.; Lübeck, M.; Lübeck, P.S.; Ahring, B.K. Fungal β-glucosidases: A bottleneck in industrial use of lignocellulosic materials. Biomolecules 2013, 3, 612–631. [Google Scholar] [CrossRef] [PubMed]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and Biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 506–577. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda, R.H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Singhania, R.R.; Patel, A.K.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of β-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 2013, 127, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Qin, Y.; Liu, Z.; Liu, K.; Wang, F.; Qu, Y. Isolation and characterization of a β-glucosidase from Penicillium decumbens and improving hydrolysis of corncob residue by using it as cellulase supplementation. Enzym. Microb. Technol. 2010, 46, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jiang, Y.; Fan, X.J.; Liu, Y.H. Molecular cloning and characterization of a novel β-glucosidase with high hydrolyzing ability for soybean isoflavone glycosides and glucose-tolerance from soil metagenomic library. Bioresour. Technol. 2012, 123, 15–22. [Google Scholar] [CrossRef] [PubMed]
- González-Pombo, P.; Fariña, L.; Carrau, F.; Batista-Viera, F.; Brena, B.M. A novel extracellular β-glucosidase from Issatchenkia terricola: Isolation, immobilization and application for aroma enhancement of white Muscat wine. Process Biochem. 2011, 46, 385–389. [Google Scholar] [CrossRef]
- Singh, G.; Verma, A.K.; Kumar, V. Catalytic properties, functional attributes and industrial applications of β-glucosidases. 3 Biotech 2016, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Crespim, E.; Zanphorlin, L.M.; Souza, F.H.M.; Diogo, J.A.; Gazolla, A.C.; Machado, C.B.; Figueiredo, F.; Sousa, A.S.; Nóbrega, F.; Pellizari, V.H.; et al. A novel cold-adapted and glucose-tolerant GH1 β-glucosidase from Exiguobacterium antarcticum B7. Int. J. Biol. Macromol. 2016, 82, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Aswathy, U.S.; Sukumaran, R.K.; Devi, G.L.; Rajasree, K.P.; Singhania, R.R.; Pandey, A. Bio-ethanol from water hyacinth biomass: An evaluation of enzymatic saccharification strategy. Bioresour. Technol. 2010, 101, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Zanphorlin, L.M.; de Giuseppe, P.O.; Honorato, R.V.; Tonoli, C.C.C.; Fattori, J.; Crespim, E.; de Oliveira, P.S.; Ruller, R.; Murakami, M.T. Oligomerization as a strategy for cold adaptation: Structure and dynamics of the GH1 β-glucosidase from Exiguobacterium antarcticum B7. Sci. Rep. 2016, 6, 23776. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Cao, L. Immobilised enzymes: Science or art? Curr. Opin. Chem. Biol. 2005, 9, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Homaei, A.A.; Sariri, R.; Vianello, F.; Stevanato, R. Enzyme immobilization: An update. J. Chem. Biol. 2013, 6, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Porto, M.D.A.; Santos, J.P.; Hackbart, H.; Bruni, G.P.; Fonseca, L.M.; Zavareze, E.R.; Dias, A.R.G. Immobilization of α-amylase in ultrafine polyvinyl alcohol (PVA) fibers via electrospinning and their stability on different substrates. Int. J. Biol. Macromol. 2019, 126, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Bolivar, J.M.; Rocha-Martin, J.; Mateo, C.; Cava, F.; Berenguer, J.; Fernandez-Lafuente, R.; Guisan, J.M. Coating of soluble and immobilized enzymes with ionic polymers: Full stabilization of the quaternary structure of multimeric enzymes. Biomacromolecules 2009, 10, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Peirce, S.; Tacias-Pascacio, V.G.; Russo, M.E.; Marzocchella, A.; Virgen-Ortíz, J.J.; Fernandez-Lafuente, R. Stabilization of Candida antarctica Lipase B (CALB) immobilized on octyl agarose by treatment with polyethylenimine (PEI). Molecules 2016, 21, 571. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Husain, Q. Effect of metal ions present in milk on the structure and functional integrity of native and polyaniline chitosan nanocomposites bound β-galactosidase: A multi-spectroscopic approach. Food Chem. 2019, 279, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.F.; Vieira, A.M.S.; Zanin, G.M.; Tardioli, P.W.; Mateo, C.; Guisán, J.M. β-glucosidase immobilized and stabilized on agarose matrix functionalized with distinct reactive groups. J. Mol. Catal. B Enzym. 2011, 69, 47–53. [Google Scholar] [CrossRef]
- Tu, M.; Zhang, X.; Kurabi, A.; Gilkes, N.; Mabee, W.; Saddler, J. Immobilization of β-glucosidase on Eupergit C for lignocellulose hydrolysis. Biotechnol. Lett. 2006, 28, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Borges, D.G.; Junior, A.B.; Farinas, C.S.; Giordano, R.L.C.; Tardioli, P.W. Enhanced saccharification of sugarcane bagasse using soluble cellulase supplemented with immobilized β-glucosidase. Bioresour. Technol. 2014, 167, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.; Wang, J.; Lu, C.; Xu, Y.; Sun, Z. Immobilized β-glucosidase on magnetic chitosan microspheres for hydrolysis of straw cellulose. Process Biochem. 2013, 48, 683–687. [Google Scholar] [CrossRef]
- Verma, M.L.; Chaudhary, R.; Tsuzuki, T.; Barrow, C.J.; Puri, M. Immobilization of β-glucosidase on a magnetic nanoparticle improves thermostability: Application in cellobiose hydrolysis. Bioresour. Technol. 2013, 135, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, D.; Pan, J.; Wang, J.; Zhao, H.; Li, Q.; Zhou, X. Efficient resveratrol production by immobilized β-glucosidase on cross-linked chitosan microsphere modified by l-lysine. J. Mol. Catal. B Enzym. 2014, 104, 29–34. [Google Scholar] [CrossRef]
- Chang, M.; Kao, H.; Juang, R. Thermal inactivation and reactivity of β-glucosidase immobilized on chitosan-clay composite. Int. J. Biol. Macromol. 2008, 43, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Alftrén, J.; Ekelund, K.; John, T. Enzymatic In vivo biotinylation of recombinant β-glucosidase enables simultaneous purification and immobilization on streptavidin coated magnetic particles. J. Mol. Catal. B Enzym. 2013, 94, 29–35. [Google Scholar] [CrossRef]
- Zhou, Y.; Yu, S.; Liu, Q.; Yan, D.; Wang, Y.; Gao, L.; Han, J.; Shi, H. Synchronized purification and immobilization of his-tagged β-glucosidase via Fe3O4/PMG core/shell magnetic nanoparticles. Sci. Rep. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhi, W.; Wan, J.; Han, J.; Li, C.; Wang, Y. Recyclable β-glucosidase by one-pot encapsulation with Cu-MOFs for enhanced hydrolysis of cellulose to glucose. ACS Sustain. Chem. Eng. 2019, 7, 3339–3348. [Google Scholar] [CrossRef]
- Melo, R.R.; Alnoch, R.C.; Vilela, A.F.L.; Souza, E.M.; Krieger, N.; Ruller, R.; Sato, H.H.; Mateo, C. New heterofunctional supports based on glutaraldehyde-activation: A tool for enzyme immobilization at neutral pH. Molecules 2017, 22, 1088. [Google Scholar] [CrossRef] [PubMed]
- Balcão, V.M.; Mateo, C.; Fernández-Lafuente, R.; Malcata, F.X.; Guisán, J.M. Coimmobilization of l-asparaginase and glutamate dehydrogenase onto highly activated supports. Enzym. Microb. Technol. 2001, 28, 696–704. [Google Scholar] [CrossRef]
- Bolivar, J.M.; Rocha-Martín, J.; Mateo, C.; Guisan, J.M. Stabilization of a highly active but unstable alcohol dehydrogenase from yeast using immobilization and post-immobilization techniques. Process Biochem. 2012, 47, 679–686. [Google Scholar] [CrossRef]
- Fernandez-Lafuente, R. Stabilization of multimeric enzymes: Strategies to prevent subunit dissociation. Enzym. Microb. Technol. 2009, 45, 405–418. [Google Scholar] [CrossRef]
- Lopez-Gallego, F.; Betancor, L.; Hidalgo, A.; Dellamora-Ortiz, G.; Mateo, C.; Fernandez-Lafuente, R.; Guisan, J.M. Stabilization of different alcohol oxidases via immobilization and post immobilization techniques. Enzym. Microb. Technol. 2007, 40, 278–284. [Google Scholar] [CrossRef]
- Rocha-Martin, J.; Costa, A.; Berenguer, J.; Guisan, J.M.; Lopez-Gallego, F. Selective oxidation of glycerol to 1,3-dihydroxyacetone by covalently immobilized glycerol dehydrogenases with higher stability and lower product inhibition. Bioresour. Technol. 2014, 170, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Marcuello, C.; Miguel, R.; Gómez-Moreno, C.; Martínez-Júlvez, M.; Lostao, A. An efficient method for enzyme immobilization evidenced by atomic force microscopy. Protein Eng. Des. Sel. 2012, 25, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henley, J.P.; Sadana, A. Deactivation theory. Biotechnol. Bioeng. 1986, 28, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Serra, I.; Serra, C.D.; Rocchietti, S.; Ubiali, D.; Terreni, M. Stabilization of thymidine phosphorylase from Escherichia coli by immobilization and post immobilization techniques. Enzym. Microb. Technol. 2011, 49, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Hegedüs, I.; Hancsók, J.; Nagy, E. Stabilization of the cellulase enzyme complex as enzyme nanoparticle. Appl. Biochem. Biotechnol. 2012, 168, 1372–1383. [Google Scholar] [CrossRef] [PubMed]
- Samak, N.A.; Tan, Y.; Sui, K.; Xia, T.; Wang, K.; Guo, C.; Liu, C. CotA laccase immobilized on functionalized magnetic graphene oxide nano-sheets for efficient biocatalysis. Mol. Catal. 2018, 445, 269–278. [Google Scholar] [CrossRef]
- Driss, D.; Driss, Z.; Chaari, F.; Chaabouni, S.E. Immobilization of his-tagged recombinant xylanase from Penicillium occitanis on nickel-chelate Eupergit C for increasing digestibility of poultry feed. Bioengineered 2014, 5, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Mohd Azhar, S.H.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Mohd Faik, A.A.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Olsson, L.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates for ethanol production. Enzym. Microb. Technol. 1996, 229, 312–331. [Google Scholar] [CrossRef]
- Cardona, C.A.; Sánchez, O.J. Fuel ethanol production: Process design trends and integration opportunities. Bioresour. Technol. 2007, 98, 2415–2457. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.F.L.; da Silva, S.F.R.; Gonçalves, F.A.; da Paz, M.F.; Fonseca, G.G.; Leite, R.S.R. Production of β-glucosidase on solid-state fermentation by Lichtheimia ramosa in agroindustrial residues: Characterization and catalytic properties of the enzymatic extract. Electron. J. Biotechnol. 2015, 18, 314–319. [Google Scholar] [CrossRef]
- Sun, S.; Sun, S.; Cao, X.; Sun, R. The role of pretreatment in improving the enzymatic hydrolysis of lignocellulosic materials. Bioresour. Technol. 2016, 199, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, H.; Lin, C.; Xu, B. Optimization of rice wine fermentation process based on the simultaneous saccharification and fermentation kinetic model. Chin. J. Chem. Eng. 2016, 24, 1406–1412. [Google Scholar] [CrossRef]
- Figueira, J.D.A.; Dias, F.F.G.; Sato, H.H.; Fernandes, P. Screening of supports for the immobilization of β-glucosidase. Enzyme Res. 2011. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Luo, J.; Wan, Y. Immobilization of cellulase on a core-shell structured metal-organic framework composites: Better inhibitors tolerance and easier recycling. Bioresour. Technol. 2018, 268, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Cao, M.; Wen, H.; Tan, Z.; Jia, S.; Cui, J. Biodegradation of polyvinyl alcohol using cross-linked enzyme aggregates of degrading enzymes from Bacillus niacini. Int. J. Biol. Macromol. 2019, 124, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ge, D.; Xu, Z. Preparation and characterization of stable chitosan nanofibrous membrane for lipase immobilization. Eur. Polym. J. 2007, 43, 3710–3718. [Google Scholar] [CrossRef]
- Kartal, F.; Janssen, M.H.A.; Hollmann, F.; Sheldon, R.A.; Kılınc, A. Improved esterification activity of Candida rugosa lipase in organic solvent by immobilization as cross-linked enzyme aggregates (CLEAs). J. Mol. Catal. B Enzym. 2011, 71, 85–89. [Google Scholar] [CrossRef]
- Hoo, M.; Park, S.; Hwan, Y.; Won, K.; Hyun, S. Immobilization of formate dehydrogenase from Candida boidinii through cross-linked enzyme aggregates. J. Mol. Catal. B Enzym. 2013, 97, 209–214. [Google Scholar]
- Mahmod, S.S.; Yusof, F.; Jami, M.S.; Khanahmadi, S.; Shah, H. Development of an immobilized biocatalyst with lipase and protease activities as a multipurpose cross-linked enzyme aggregate. Process Biochem. 2015, 50, 2144–2157. [Google Scholar] [CrossRef]
- Jin, X.; Li, S.; Long, N.; Zhang, R. A robust and stable nano-biocatalyst by co-immobilization of chloroperoxidase and horseradish peroxidase for the decolorization of azo dyes. J. Chem. Technol. Biotechnol. 2018, 93, 489–497. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Mateo, C.; Bolivar, J.M.; Godoy, C.A.; Rocha-Martin, J.; Pessela, B.C.; Curiel, J.A.; Muñoz, R.; Guisan, J.M.; Fernández-Lorente, G. Improvement of enzyme properties with a two-step immobilization process on novel heterofunctional supports. Biomacromolecules 2010, 11, 3112–3117. [Google Scholar] [CrossRef] [PubMed]
- Whistler, R.L.; Wolfrom, M.L.; Miller, J. Methods in Carbohydrate Chemistry, 2nd ed.; Academic Press: New York, NY, USA; London, UK, 1963. [Google Scholar]
- Betancor, L.; López-Gallego, F.; Hidalgo, A.; Alonso-Morales, N.; Dellamora-Ortiz, G.; Mateo, C.; Fernández-Lorente, G.; Guisan, J.M. Different mechanisms of protein immobilization on glutaraldehyde activated supports: Effect of support activation and immobilization conditions. Enzym. Microb. Technol. 2006, 39, 877–882. [Google Scholar] [CrossRef]
- Addorisio, V.; Sannino, F.; Mateo, C.; Guisan, J.M. Oxidation of phenyl compounds using strongly stable immobilized-stabilized laccase from Trametes versicolor. Process Biochem. 2013, 48, 1174–1180. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Support | Treatment | Immobilization Efficiency a (%), EI | Recovered Activity after Treatments b (%), R |
|---|---|---|---|
| Agarose-GLU | Control (without treatment) | 100 | 50 |
| Glycine 200 mM, pH 7.0 | 100 | 34 | |
| Aspartic acid 200 mM, pH 7.0 | 100 | 47 | |
| DEAE-Dextran 20 mg mL−1, pH 7.0 | 100 | 51 | |
| Polygalacturonic acid 20 mg mL−1, pH 7.0 | 100 | 54 | |
| Polyethylenimine 20 mg mL−1, pH 7.0 | 100 | 32 | |
| Glycine 200 mM and polygalacturonic acid 20 mg mL−1, pH 7.0 | 100 | 56 | |
| Glycine 200 mM and polyethylenimine 20 mg mL−1, pH 7.0 | 100 | 19 | |
| Agarose-GLU-IDA-Co2+ | Control (without treatment) | 100 | 64 |
| Glycine 200 mM, pH 7.0 | 100 | 63 | |
| Aspartic acid 200 mM, pH 7.0 | 100 | 68 | |
| DEAE-Dextran 20 mg mL−1, pH 7.0 | 100 | 8 | |
| Polygalacturonic acid 20 mg mL−1, pH 7.0 | 100 | 64 | |
| Polyethylenimine 20 mg mL−1, pH 7.0 | 100 | 55 | |
| Glycine 200 mM and polygalacturonic acid 20 mg mL−1, pH 7.0 | 100 | 63 | |
| Glycine 200 mM and polyethylenimine 20 mg mL−1, pH 7.0 | 100 | 10 |
| Poly-Functional Macromolecule | Structure | |
|---|---|---|
| Polygalacturonic acid | Anionic |  |
| Glycine | Zwitterion |  |
| Aspartic acid | Anionic |  |
| DEAE-Dextran | Cationic |  |
| Polyethylenimine | Cationic |  |
| Support | Treatment | Half Life (T1/2, min 40 °C) | Stability Factor |
|---|---|---|---|
| Free EaBglA | 21 | ||
| Agarose-GLU | Control (without treatment) | 39 | 1.8 |
| Glycine 200 mM, pH 7.0 | 49 | 2.3 | |
| Aspartic acid 200 mM, pH 7.0 | 26 | 1.2 | |
| DEAE-Dextran 20 mg mL−1, pH 7.0 | 25 | 1.2 | |
| Polygalacturonic acid 20 mg mL−1, pH 7.0 | 30 | 1.4 | |
| Polyethylenimine 20 mg mL−1, pH 7.0 | 50 | 2.4 | |
| Glycine 200 mM and polygalacturonic acid 20 mg mL−1, pH 7.0 | 30 | 1.4 | |
| Glycine 200 mM and polyethylenimine 20 mg mL−1, pH 7.0 | 22 | 1.0 | |
| Agarose-GLU-IDA-Co2+ | Control (without treatment) | 230 | 11 |
| Glycine 200 mM, pH 7.0 | 40 | 1.9 | |
| Aspartic acid 200 mM, pH 7.0 | 23 | 1.1 | |
| DEAE-Dextran 20 mg mL−1, pH 7.0 | 28 | 1.3 | |
| Polygalacturonic acid 20 mg mL−1, pH 7.0 | 60 | 2.9 | |
| Polyethylenimine 20 mg mL−1, pH 7.0 | 960 | 46 | |
| Glycine 200 mM and polygalacturonic acid 20 mg mL−1, pH 7.0 | 55 | 2.6 | |
| Glycine 200 mM and polyethylenimine 20 mg mL−1, pH 7.0 | 28 | 1.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues de Melo, R.; Alnoch, R.C.; Sousa, A.S.d.; Harumi Sato, H.; Ruller, R.; Mateo, C. Cross-Linking with Polyethylenimine Confers Better Functional Characteristics to an Immobilized β-glucosidase from Exiguobacterium antarcticum B7. Catalysts 2019, 9, 223. https://doi.org/10.3390/catal9030223
Rodrigues de Melo R, Alnoch RC, Sousa ASd, Harumi Sato H, Ruller R, Mateo C. Cross-Linking with Polyethylenimine Confers Better Functional Characteristics to an Immobilized β-glucosidase from Exiguobacterium antarcticum B7. Catalysts. 2019; 9(3):223. https://doi.org/10.3390/catal9030223
Chicago/Turabian StyleRodrigues de Melo, Ricardo, Robson Carlos Alnoch, Amanda Silva de Sousa, Hélia Harumi Sato, Roberto Ruller, and Cesar Mateo. 2019. "Cross-Linking with Polyethylenimine Confers Better Functional Characteristics to an Immobilized β-glucosidase from Exiguobacterium antarcticum B7" Catalysts 9, no. 3: 223. https://doi.org/10.3390/catal9030223
APA StyleRodrigues de Melo, R., Alnoch, R. C., Sousa, A. S. d., Harumi Sato, H., Ruller, R., & Mateo, C. (2019). Cross-Linking with Polyethylenimine Confers Better Functional Characteristics to an Immobilized β-glucosidase from Exiguobacterium antarcticum B7. Catalysts, 9(3), 223. https://doi.org/10.3390/catal9030223