Advanced Rhodococcus Biocatalysts for Environmental Biotechnologies



Abstract
:1. Biotechnological Advantages of Actinobacteria of the Genus Rhodococcus and Approaches to Obtain Stable Biocatalysts
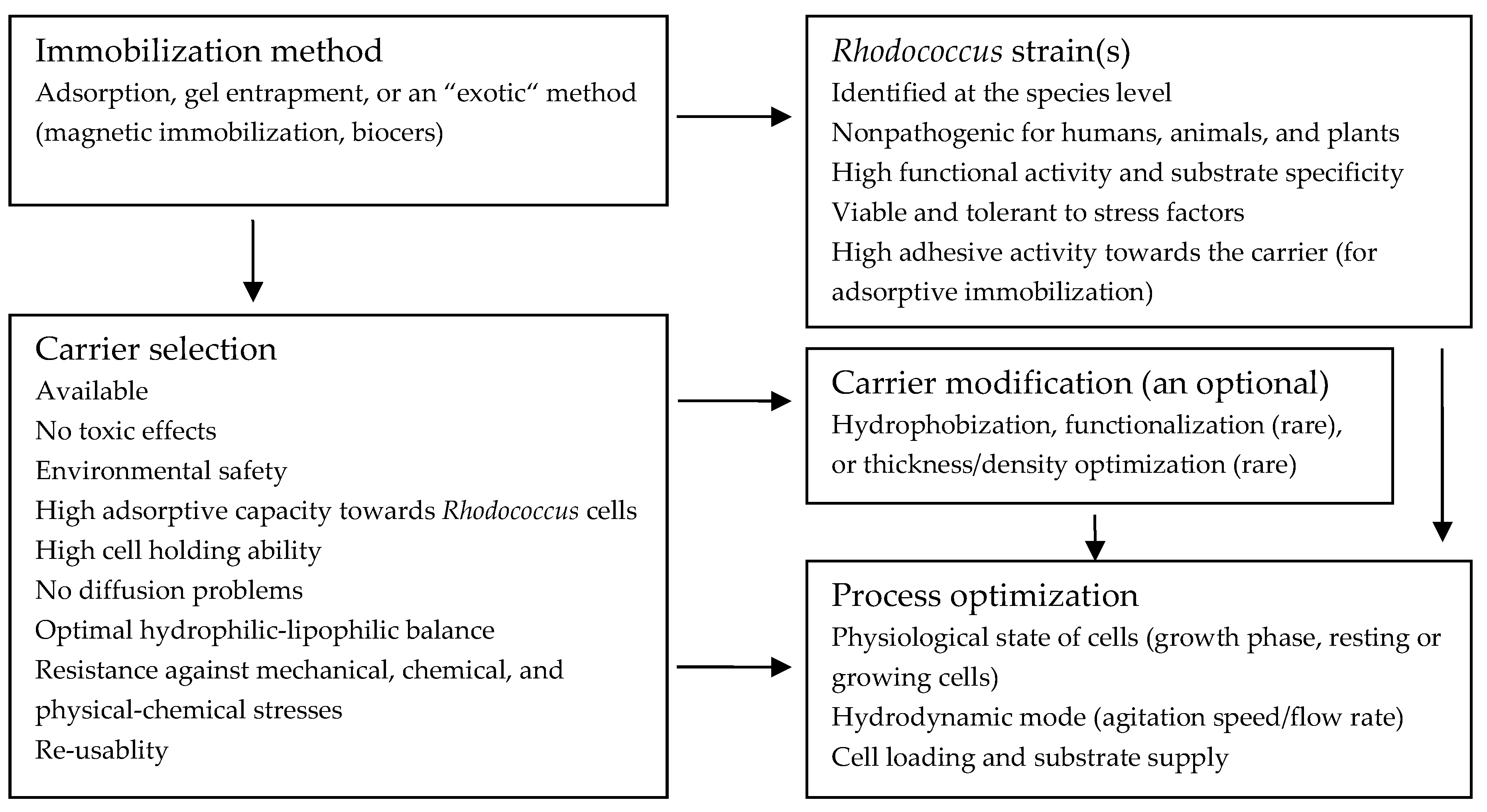
2. Guidelines for Immobilization of Rhodococcus on/into Carriers
2.1. Advantages of Immobilized Cells
2.2. Choice of the Immobilization Method
2.3. Selection of Bacterial Strains
2.4. Selection of a Carrier for Cell Immobilization
2.5. Process Optimization Strategies
3. Genetic Modifications of Rhodococcus
4. Acclimation of Rhodococcus Cells to Toxic Substrates
5. Microbial Consortia/Associations Containing Rhodococcus
6. Environmental Applications of Rhodococcus Biocatalysts
7. Conclusions
Funding
Conflicts of Interest
References
- Chen, M.; Xu, P.; Zeng, G.; Yang, C.; Huang, D.; Zhang, J. Bioremediation of soils contaminated with polycyclic aromatic hydrocarbons, petroleum, pesticides, chlorophenols and heavy metals by composting: Applications, microbes and future research needs. Biotechnol. Adv. 2015, 33, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Elkin, A.A.; Makarov, S.O.; Cunningham, C.J.; Peshkur, T.A.; Atlas, R.M.; Philp, J.C. Oil spill problems and sustainable response strategies through new technologies. Environ. Sci. Process. Impacts 2015, 17, 1201–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, A.; Peerzada, M.H.; Mohsin, M.; White, M. A review on developments in dyeing cotton fabrics with reactive dyes for reducing effluent pollution. J. Clean. Prod. 2015, 87, 50–57. [Google Scholar] [CrossRef]
- Horton, A.A.; Walton, A.; Spurgeon, D.J.; Lahive, E.; Svendsen, C. Microplastics in freshwater and terrestrial environments: Evaluating the current understanding to identify the knowledge gaps and future research priorities. Sci. Total Environ. 2017, 586, 127–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awasthi, K.A.; Wang, M.; Awasti, K.M.; Wang, Z.; Li, J. Environmental pollution and human body burden from improper recycling of e-waste in China: A short-review. Environ. Pollut. 2018, 243, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Luo, Y.; Lu, S.; Liu, M.; Song, Y.; Lei, L. Microplastics in soils: Analytical methods, pollution characteristics and ecological risks. Trends Anal. Chem. 2018, 109, 163–172. [Google Scholar] [CrossRef]
- Kanter, D.R.; Searchinger, T.D. A technology-forcing approach to reduce nitrogen pollution. Nat. Sustain. 2018, 1, 544–552. [Google Scholar] [CrossRef]
- Xu, X.; Nie, S.; Ding, H.; Hou, F.F. Environmental pollution and kidney diseases. Nat. Rev. Nephrol. 2018, 14, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, P.; Raj, A.; Bharagava, R.N. Environmental pollution and health hazards from distillery wastewater and treatment approaches to combat the environmental threats: A review. Chemosphere 2018, 194, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.; Tyumina, E.; Vikhareva, E. Biodegradation of emerging pollutants: Focus on pharmaceuticals. Microbiol. Aust. 2018, 39, 117–122. [Google Scholar] [CrossRef]
- Vilela, C.L.S.; Bassin, J.P.; Peixoto, R.S. Water contamination by endocrine disruptors: Impacts, microbiological aspects and trends for environmental protection. Environ. Pollut. 2018, 235, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, Y.; Chen, Q.; Luo, H.; Zhu, Z.; Chen, W.; Chen, J.; Mo, Y. Biochemical pathways and enhanced degradation of di-n-octyl phthalate (DOP) in sequencing batch reactor (SBR) by Arthrobacter sp. SLG-4 and Rhodococcus sp. SLG-6 isolated from activated sludge. Biodegradation 2018, 29, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.B.; Hidalgo, A.; Serra, J.L.; Llama, M.J. Degradation of phenol by Rhodococcus erythropolis UPV-1 immobilized on Biolite® in a packed-bed reactor. J. Biotechnol. 2002, 97, 1–11. [Google Scholar] [CrossRef]
- Megharaj, M.; Ramakrishnan, B.; Venkateswarlu, K.; Sethunathan, N.; Naidu, R. Bioremediation approaches for organic pollutants: A critical perspective. Environ. Int. 2011, 37, 1362–1375. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, S.; Singh, D.K. Biodegradation of persistent organic pollutants in soil, water and pristine sites by cold-adapted microorganisms: Mini review. Int. Biodeterior. Biodegrad. 2015, 100, 98–105. [Google Scholar] [CrossRef]
- de Carvalho, C.C.C.R.; da Fonseca, M.M.R. The remarkable Rhodococcus erythropolis. Appl. Microbiol. Biotechnol. 2005, 67, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.J.; Kulakov, L.A.; Allen, C.C.R. Biodegradation and Rhodococcus—Masters of catabolic versatility. Curr. Opin. Biotechnol. 2005, 16, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Martínková, L.; Uhnáková, B.; Pátek, M.; Nešvera, J.; Křen, V. Biodegradation potential of the genus Rhodococcus. Environ. Int. 2009, 35, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Kuyukina, M.S.; Ivshina, I.B. Application of Rhodococcus in bioremediation of contaminated environments. In Biology of Rhodococcus; Alvarez, H.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 231–262. ISBN 978-3-642-12936-0. [Google Scholar]
- Larkin, M.J.; Kulakov, L.A.; Allen, C.C.R. Rhodococcus. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1839–1852. ISBN 9783540775874. [Google Scholar]
- Yam, K.C.; van der Geize, R.; Eltis, L.D. Catabolism of aromatic compounds and steroids by Rhodococcus. In Biology of Rhodococcus; Alvarez, H.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 133–169. ISBN 978-3-642-12936-0. [Google Scholar]
- de Carvalho, C.C.C.R.; Costa, S.S.; Fernandes, P.; Couto, I.; Viveiros, M. Membrane transport systems and the biodegradation potential and pathogenicity of genus Rhodococcus. Front. Physiol. 2014, 5, 133. [Google Scholar] [CrossRef] [PubMed]
- Pogorevc, M.; Faber, K. Purification and characterization of an inverting stereo- and enantioselective sec-alkylsulfatase from the gram-positive bacterium Rhodococcus ruber DSM 44541. Appl. Environ. Microbiol. 2003, 69, 2810–2815. [Google Scholar] [CrossRef] [PubMed]
- Koch, D.J.; Chen, M.M.; Van Beilen, J.B.; Arnold, F.H. In vivo evolution of butane oxidation by terminal alkane hydroxylases AlkB and CYP153A6. Appl. Environ. Microbiol. 2009, 75, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Choi, K.Y.; Yoo, M.; Choi, J.N.; Lee, C.H.; Zylstra, G.J.; Kang, B.S.; Kim, E. Benzylic and aryl hydroxylations of m-xylene by o-xylene dioxygenase from Rhodococcus sp. strain DK17. Appl. Microbiol. Biotechnol. 2010, 86, 1841–1847. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, S.; Van Petegem, F.; Patrauchan, M.A.; Eltis, L.D. AnhE, a metallochaperone involved in the maturation of a cobalt-dependent nitrile hydratase. J. Biol. Chem. 2010, 285, 25126–25133. [Google Scholar] [CrossRef] [PubMed]
- Saa, L.; Jaureguibeitia, A.; Largo, E.; Llama, M.J.; Serra, J.L. Cloning, purification and characterization of two components of phenol hydroxylase from Rhodococcus erythropolis UPV-1. Appl. Microbiol. Biotechnol. 2010, 86, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Fernández de las Heras, L.; van der Geize, R.; Drzyzga, O.; Perera, J.; María Navarro Llorens, J. Molecular characterization of three 3-ketosteroid-Δ1-dehydrogenase isoenzymes of Rhodococcus ruber strain Chol-4. J. Steroid Biochem. Mol. Biol. 2012, 132, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Grigg, J.C.; Armstrong, Z.; Murphy, M.E.P.; Eltis, L.D. Distal heme pocket residues of B-type dye-decolorizing peroxidase: Arginine but not aspartate is essential for peroxidase activity. J. Biol. Chem. 2012, 287, 10623–10630. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rojas, E.; Kurt, T.; Schmidt, U.; Meyer, V.; Garbe, L.A. A bifunctional enzyme from Rhodococcus erythropolis exhibiting secondary alcohol dehydrogenase-catalase activities. Appl. Microbiol. Biotechnol. 2014, 98, 9249–9258. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Barbe, V.; Fischer, C. Draft genome sequence of propane- and butane-oxidizing actinobacterium Rhodococcus ruber IEGM 231. Genome Announc. 2014, 2, e01297-14. [Google Scholar] [CrossRef] [PubMed]
- McLeod, M.P.; Warren, R.L.; Hsiao, W.W.L.; Araki, N.; Myhre, M.; Fernandes, C.; Miyazawa, D.; Wong, W.; Lillquist, A.L.; Wang, D.; et al. The complete genome of Rhodococcus sp. RHA1 provides insights into a catabolic powerhouse. Proc. Natl. Acad. Sci. USA 2006, 103, 15582–15587. [Google Scholar] [CrossRef] [PubMed]
- Takeda, H.; Shimodaira, J.; Yukawa, K.; Hara, N.; Kasai, D.; Miyauchi, K.; Masai, E.; Fukuda, M. Dual two-component regulatory systems are involved in aromatic compound degradation in a polychlorinated-biphenyl degrader, Rhodococcus jostii RHA1. J. Bacteriol. 2010, 192, 4741–4751. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, J.A.; Zhao, Q.; Li, X.; Shu, W. Bioremediation of environmental endocrine disruptor di-n-butyl phthalate ester by Rhodococcus ruber. Chemosphere 2006, 65, 1627–1633. [Google Scholar] [CrossRef] [PubMed]
- Elkin, A.A.; Kylosova, T.I.; Grishko, V.V.; Ivshina, I.B. Enantioselective oxidation of sulfides to sulfoxides by Gordonia terrae IEGM 136 and Rhodococcus rhodochrous IEGM 66. J. Mol. Catal. B Enzym. 2013, 89, 82–85. [Google Scholar] [CrossRef]
- Gilan, I.; Sivan, A. Effect of proteases on biofilm formation of the plastic-degrading actinomycete Rhodococcus ruber C208. FEMS Microbiol. Lett. 2013, 342, 18–23. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Niu, C.; Lu, Z. Individual or synchronous biodegradation of di-n-butyl phthalate and phenol by Rhodococcus ruber strain DP-2. J. Hazard. Mater. 2014, 273, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Cheremnykh, K.M.; Luchnikova, N.A.; Grishko, V.V.; Ivshina, I.B. Bioconversion of ecotoxic dehydroabietic acid using Rhodococcus actinobacteria. J. Hazard. Mater. 2018, 346, 103–112. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, D.; Evangelista, S.; Yargeau, V. Removal of aqueous 17α-ethinylestradiol by Rhodococcus species. Environ. Eng. Sci. 2009, 26, 1393–1400. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Vikhareva, E.V.; Richkova, M.I.; Mukhutdinova, A.N.; Karpenko, J.N. Biodegradation of drotaverine hydrochloride by free and immobilized cells of Rhodococcus rhodochrous IEGM 608. World J. Microbiol. Biotechnol. 2012, 28, 2997–3006. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, H.M.; Silva, R.A. Metabolic diversity and flexibility for hydrocarbon biodegradation by Rhodococcus. In Actinobacteria: Application in Bioremediation and Production of Industrial Enzymes; Amoroso, M.J., Benimeli, C.S., Cuozzo, S.A., Eds.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2013; pp. 241–273. [Google Scholar]
- Gilan (Orr), I.; Hadar, Y.; Sivan, A. Colonization, biofilm formation and biodegradation of polyethylene by a strain of Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2004, 65, 97–104. [Google Scholar] [CrossRef]
- Sivan, A.; Szanto, M.; Pavlov, V. Biofilm development of the polyethylene-degrading bacterium Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2006, 72, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Krueger, M.C.; Harms, H.; Schlosser, D. Prospects for microbiological solutions to environmental pollution with plastics. Appl. Microbiol. Biotechnol. 2015, 99, 8857–8874. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Philp, J.C. Surface-active lipids in rhodococci. Antonie Van Leeuwenhoek 1998, 74, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Kuyukina, M.S.; Ivshina, I.B. Rhodococcus biosurfactants: Biosynthesis, properties, and potential applications. In Biology of Rhodococcus; Alvarez, H.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 291–313. ISBN 978-3-642-12936-0. [Google Scholar]
- Cortes, M.A.L.R.M.; de Carvalho, C.C.C.R. Effect of carbon sources on lipid accumulation in Rhodococcus cells. Biochem. Eng. J. 2015, 94, 100–105. [Google Scholar] [CrossRef]
- Pieper-Furst, U.; Madkour, M.H.; Mayer, F.; Steinbuchel, A. Identification of the region of a 14-kilodalton protein of Rhodococcus ruber that is responsible for the binding of this phasin to polyhydroxyalkanoic acid granules. J. Bacteriol. 1995, 177, 2513–2523. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Kobayashi, A.; Ikeda, H.; Unno, H. Rhodococcus aetherivorans IAR1, a new bacterial strain synthesizing poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from toluene. J. Biosci. Bioeng. 2009, 107, 145–150. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Ivshina, I.B.; Baeva, T.A.; Kochina, O.A.; Gein, S.V.; Chereshnev, V.A. Trehalolipid biosurfactants from nonpathogenic Rhodococcus actinobacteria with diverse immunomodulatory activities. New Biotechnol. 2015, 32, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Gasser, I.; Schmid, F.; Berg, G. Linking ecology with economy: Insights into polyhydroxyalkanoate-producing microorganisms. Eng. Life Sci. 2011, 11, 222–237. [Google Scholar] [CrossRef]
- Vandeputte, O.; Öden, S.; Mol, A.; Vereecke, D.; Goethals, K.; El Jaziri, M.; Prinsen, E. Biosynthesis of auxin by the gram-positive phytopathogen Rhodococcus fascians is controlled by compounds specific to infected plant tissues. Appl. Environ. Microbiol. 2005, 71, 1169–1177. [Google Scholar] [CrossRef]
- Kämpfer, P.; Dott, W.; Martin, K.; Glaeser, S.P. Rhodococcus defluvii sp. nov., isolated from wastewater of a bioreactor and formal proposal to reclassify [Corynebacterium hoagii] and Rhodococcus equi as Rhodococcus hoagii comb. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 755–761. [Google Scholar] [CrossRef]
- Koronelli, T.V. Principles and methods for raising the efficiency of biological degradation of hydrocarbons in the environment: Review. Appl. Biochem. Microbiol. 1996, 32, 519–525. [Google Scholar] [CrossRef]
- Qin, S.; Xing, K.; Jiang, J.H.; Xu, L.H.; Li, W.J. Biodiversity, bioactive natural products and biotechnological potential of plant-associated endophytic actinobacteria. Appl. Microbiol. Biotechnol. 2011, 89, 457–473. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B. Current situation and challenges of specialized microbial resource centres in Russia. Microbiology 2012, 81, 509–516. [Google Scholar] [CrossRef]
- Sorkhoh, N.A.; Ghannoum, M.A.; Ibrahim, A.S.; Stretton, R.J.; Radwan, S.S. Crude oil and hydrocarbon-degrading strains of Rhodococcus rhodochrous isolated from soil and marine environments in Kuwait. Environ. Pollut. 1990, 65, 1–17. [Google Scholar] [CrossRef]
- Paliwal, V.; Puranik, S.; Purohit, H.J. Integrated perspective for effective bioremediation. Appl. Biochem. Biotechnol. 2012, 166, 903–924. [Google Scholar] [CrossRef] [PubMed]
- Panicker, G.; Mojib, N.; Aislabie, J.; Bej, A.K. Detection, expression and quantitation of the biodegradative genes in Antarctic microorganisms using PCR. Antonie van Leeuwenhoek 2010, 97, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Bej, A.K.; Saul, D.; Aislabie, J. Cold-tolerant alkane-degrading Rhodococcus species from Antarctica. Polar Biol. 2000, 23, 100–105. [Google Scholar] [CrossRef]
- Whyte, L.G.; Goalen, B.; Hawari, J.; Labbe, D.; Greer, C.W.; Nahir, M. Bioremediation treatability assessment of hydrocarbon-contaminated soils from Eureka, Nunavut. Cold Reg. Sci. Technol. 2001, 32, 121–132. [Google Scholar] [CrossRef]
- de Carvalho, C.C.C.R. Adaptation of Rhodococcus erythropolis cells for growth and bioremediation under extreme conditions. Res. Microbiol. 2012, 163, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Satoi, M.; Iwasa, Y.; Honda, K.; Sameshima, Y.; Omasa, T.; Kato, J.; Ohtake, H. Utilization of hydrophobic bacterium Rhodococcus opacus B-4 as whole-cell catalyst in anhydrous organic solvents. Appl. Microbiol. Biotechnol. 2007, 74, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Retamal-Morales, G.; Mehnert, M.; Schwabe, R.; Tischler, D.; Schlömann, M.; Levicán, G.J. Genomic characterization of the arsenic-tolerant actinobacterium, Rhodococcus erythropolis S43. Solid State Phenom. 2017, 262, 660–663. [Google Scholar] [CrossRef]
- Kuyukina, M.; Krivoruchko, A.; Ivshina, I. Hydrocarbon- and metal-polluted soil bioremediation: Progress and challenges. Microbiol. Aust. 2018, 39, 133–136. [Google Scholar] [CrossRef]
- Kratzer, R.; Woodley, J.M.; Nidetzky, B. Rules for biocatalyst and reaction engineering to implement effective, NAD(P)H-dependent, whole cell bioreductions. Biotechnol. Adv. 2015, 33, 1641–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto, M.B.; Hidalgo, A.; Rodríguez-Fernández, C.; Serra, J.L.; Llama, M.J. Biodegradation of phenol in synthetic and industrial wastewater by Rhodococcus erythropolis UPV-1 immobilized in an air-stirred reactor with clarifier. Appl. Microbiol. Biotechnol. 2002, 58, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Suttinun, O.; Müller, R.; Luepromchai, E. Cometabolic degradation of trichloroethene by Rhodococcus sp. strain L4 immobilized on plant materials rich in essential oils. Appl. Environ. Microbiol. 2010, 76, 4684–4690. [Google Scholar] [CrossRef] [PubMed]
- Tapingkae, W.; Parkin, K.L.; Tanasupawat, S.; Kruenate, J.; Benjakul, S.; Visessanguan, W. Whole cell immobilisation of Natrinema gari BCC 24369 for histamine degradation. Food Chem. 2010, 120, 842–849. [Google Scholar] [CrossRef]
- Pannier, A.; Mkandawire, M.; Soltmann, U.; Pompe, W.; Böttcher, H. Biological activity and mechanical stability of sol-gel-based biofilters using the freeze-gelation technique for immobilization of Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2012, 93, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Takei, T.; Yamasaki, M.; Yoshida, M. Cesium accumulation of Rhodococcus erythropolis CS98 strain immobilized in hydrogel matrices. J. Biosci. Bioeng. 2014, 117, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Eş, I.; Vieira, J.D.G.; Amaral, A.C. Principles, techniques, and applications of biocatalyst immobilization for industrial application. Appl. Microbiol. Biotechnol. 2015, 99, 2065–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, S.; Volta, A.; Santisi, S.; Morici, C.; Mancini, G.; Quatrini, P.; Genovese, M.; Yakimov, M.M.; Torregrossa, M. Oil-degrading bacteria from a membrane bioreactor (BF-MBR) system for treatment of saline oily waste: Isolation, identification and characterization of the biotechnological potential. Int. Biodeterior. Biodegrad. 2016, 110, 235–244. [Google Scholar] [CrossRef]
- Zur, J.; Wojcieszyńska, D.; Guzik, U. Metabolic responses of bacterial cells to immobilization. Molecules 2016, 21, 958. [Google Scholar] [CrossRef] [PubMed]
- Andryushina, V.; Balabanova, T.; Beklemishev, A.; Varfolomeev, S.; Vodyakova, M.; Demakov, V.; Ditchenko, T.; Dzhavahiya, V.; Drozdova, M.; Efremenko, E.; et al. Immobilized Cells: Biocatalysts and Processes; Publishing Center RIOR: Moscow, Russia, 2018; ISBN 978-5-369-02004-3. [Google Scholar]
- Kuyukina, M.S.; Ivshina, I.B.; Serebrennikova, M.K.; Krivoruchko, A.V.; Korshunova, I.O.; Peshkur, T.A.; Cunningham, C.J. Oilfield wastewater biotreatment in a fluidized-bed bioreactor using co-immobilized Rhodococcus cultures. J. Environ. Chem. Eng. 2017, 5, 1252–1260. [Google Scholar] [CrossRef]
- Wang, X.; Gong, Z.; Li, P.; Zhang, L.; Hu, X. Degradation of pyrene and benzo(a)pyrene in contaminated soil by immobilized fungi. Environ. Eng. Sci. 2008, 25, 677–684. [Google Scholar] [CrossRef]
- Li, J.; Guo, C.; Liao, C.; Zhang, M.; Liang, X.; Lu, G.; Yang, C.; Dang, Z. A bio-hybrid material for adsorption and degradation of phenanthrene: Bacteria immobilized on sawdust coated with a silica layer. RSC Adv. 2016, 6, 107189–107199. [Google Scholar] [CrossRef]
- Xiong, B.; Zhang, Y.; Hou, Y.; Arp, H.P.H.; Reid, B.J.; Cai, C. Enhanced biodegradation of PAHs in historically contaminated soil by M. gilvum inoculated biochar. Chemosphere 2017, 182, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Coles, C.A.; Patel, T.R.; Akinnola, A.P.; Helleur, R.J. Influence of bulking agents, fertilizers and bacteria on the removal of diesel from a Newfoundland soil. Soil Sediment Contam. 2009, 18, 383–396. [Google Scholar] [CrossRef]
- Rivelli, V.; Franzetti, A.; Gandolfi, I.; Cordoni, S.; Bestetti, G. Persistence and degrading activity of free and immobilised allochthonous bacteria during bioremediation of hydrocarbon-contaminated soils. Biodegradation 2013, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kauppi, S.; Sinkkonen, A.; Romantschuk, M. Enhancing bioremediation of diesel-fuel-contaminated soil in a boreal climate: Comparison of biostimulation and bioaugmentation. Int. Biodeterior. Biodegrad. 2011, 65, 359–368. [Google Scholar] [CrossRef]
- Chand, D.; Kumar, H.; Sankhian, U.D.; Kumar, D.; Vitzthum, F.; Bhalla, T.C. Treatment of simulated wastewater containing toxic amides by immobilized Rhodococcus rhodochrous NHB-2 using a highly compact 5-stage plug flow reactor. World J. Microbiol. Biotechnol. 2004, 20, 679–686. [Google Scholar] [CrossRef]
- Junter, G.A.; Jouenne, T. Immobilized viable microbial cells: From the process to the proteome... or the cart before the horse. Biotechnol. Adv. 2004, 22, 633–658. [Google Scholar] [CrossRef] [PubMed]
- Kitova, A.E.; Kuvichkina, T.N.; Arinbasarova, A.Y.; Reshetilov, A.N. Degradation of 2,4-dinitrophenol by free and immobilized cells of Rhodococcus erythropolis HL PM-1. Appl. Biochem. Microbiol. 2004, 40, 307–311. [Google Scholar] [CrossRef]
- Claudino, M.J.C.; Soares, D.; Van Keulen, F.; Marques, M.P.C.; Cabral, J.M.S.; Fernandes, P. Immobilization of mycobacterial cells onto silicone—Assessing the feasibility of the immobilized biocatalyst in the production of androstenedione from sitosterol. Bioresour. Technol. 2008, 99, 2304–2311. [Google Scholar] [CrossRef] [PubMed]
- Junter, G.A.; Coquet, L.; Vilain, S.; Jouenne, T. Immobilized-cell physiology: Current data and the potentialities of proteomics. Enzym. Microb. Technol. 2002, 31, 201–212. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Ivshina, I.B.; Serebrennikova, M.K.; Krivorutchko, A.B.; Podorozhko, E.A.; Ivanov, R.V.; Lozinsky, V.I. Petroleum-contaminated water treatment in a fluidized-bed bioreactor with immobilized Rhodococcus cells. Int. Biodeterior. Biodegrad. 2009, 63, 427–432. [Google Scholar] [CrossRef]
- Shumkova, E.S.; Solyanikova, I.P.; Plotnikova, E.G.; Golovleva, L.A. Phenol degradation by Rhodococcus opacus strain 1G. Appl. Biochem. Microbiol. 2009, 45, 51–57. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Plekhov, O.A.; Naimark, O.B.; Podorozhko, E.A.; Lozinsky, V.I. Biosurfactant-enhanced immobilization of hydrocarbon-oxidizing Rhodococcus ruber on sawdust. Appl. Microbiol. Biotechnol. 2013, 97, 5315–5327. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.E.; Christova, N.E.; Kabaivanova, L.V.; Nacheva, L.V.; Stoineva, I.B.; Petrov, P.D. Simultaneous biodegradation of phenol and n-hexadecane by cryogel immobilized biosurfactant producing strain Rhodococcus wratislaviensis BN38. Pol. J. Microbiol. 2016, 65, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Quek, E.; Ting, Y.P.; Tan, H.M. Rhodococcus sp. F92 immobilized on polyurethane foam shows ability to degrade various petroleum products. Bioresour. Technol. 2006, 97, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Kuyukina, M.S.; Ivshina, I.B.; Serebrennikova, M.K.; Rubtsova, E.V.; Krivoruchko, A.V. Simultaneous species-specific PCR detection and viability testing of poly(vinyl alcohol) cryogel-entrapped Rhodococcus spp. after their exposure to petroleum hydrocarbons. J. Microbiol. Methods 2013, 94, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Derikvand, P.; Etemadifar, Z.; Biria, D. Taguchi optimization of dibenzothiophene biodesulfurization by Rhodococcus erythropolis R1 immobilized cells in a biphasic system. Int. Biodeterior. Biodegrad. 2014, 86, 343–348. [Google Scholar] [CrossRef]
- Bhasarkar, J.B.; Dikshit, P.K.; Moholkar, V.S. Ultrasound assisted biodesulfurization of liquid fuel using free and immobilized cells of Rhodococcus rhodochrous MTCC 3552: A mechanistic investigation. Bioresour. Technol. 2015, 187, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Liu, F.; Wu, N.; Ju, J.; Yu, B. Efficient biodegradation of chlorophenols in aqueous phase by magnetically immobilized aniline-degrading Rhodococcus rhodochrous strain. J. Nanobiotechnology 2016, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Omarova, E.O.; Lobakova, E.S.; Dolnikova, G.A.; Nekrasova, V.V.; Idiatulov, R.K.; Kashcheeva, P.B.; Perevertailo, N.G.; Dedov, A.G. Immobilization of bacteria on polymer matrices for degradation of crude oil and oil products. Mosc. Univ. Biol. Sci. Bull. 2012, 67, 24–30. [Google Scholar] [CrossRef]
- Kovalenko, G.A.; Perminova, L.V.; Chuenko, T.V.; Ivshina, I.B.; Kuyukina, M.S.; Rychkova, M.I.; Philp, J.K. Carbon-containing macrostructured ceramic carriers for adsorption immobilization of enzymes and microorganisms. 5. Immobilization of yeast non-growing cells and alkanotrophic Rhodococcus growing cells. Biotechol. Russ. 2006, 1, 102–113. [Google Scholar]
- Cubitto, M.A.; Gentili, A.R. Bioremediation of crude oil-contaminated soil by immobilized bacteria on an agroindustrial waste—Sunflower seed husks. Bioremediat. J. 2015, 19, 277–286. [Google Scholar] [CrossRef]
- An, X.; Cheng, Y.; Huang, M.; Sun, Y.; Wang, H.; Chen, X.; Wang, J.; Li, D.; Li, C. Treating organic cyanide-containing groundwater by immobilization of a nitrile-degrading bacterium with a biofilm-forming bacterium using fluidized bed reactors. Environ. Pollut. 2018, 237, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Kuyukina, M.S.; Ivshina, I.B.; Gavrin, A.Y.; Podorozhko, E.A.; Lozinsky, V.I.; Jeffree, C.E.; Philp, J.C. Immobilization of hydrocarbon-oxidizing bacteria in poly(vinyl alcohol) cryogels hydrophobized using a biosurfactant. J. Microbiol. Methods 2006, 65, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Hatzinger, P.B.; Condee, C.; McClay, K.R.; Togna, A.P.; Fournier, D.; Hawari, J.; Halasz, A.; Streger, S.H.; McClay, K.R.; Masuda, H.; et al. Aerobic treatment of N-nitrosodimethylamine in a propane-fed membrane bioreactor. Water Res. 2011, 45, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Anke, M.K.; Szymańska, K.; Tischler, D. Immobilization of Rhodococcus opacus 1CP azoreductase to obtain azo dye degrading biocatalysts operative at acidic pH. Int. Biodeterior. Biodegrad. 2017, 118, 89–94. [Google Scholar] [CrossRef]
- Safonova, E.; Kvitko, K.V.; Iankevitch, M.I.; Surgko, L.F.; Afti, I.A.; Reisser, W. Biotreatment of industrial wastewater by selected algal-bacterial consortia. Eng. Life Sci. 2004, 4, 347–353. [Google Scholar] [CrossRef]
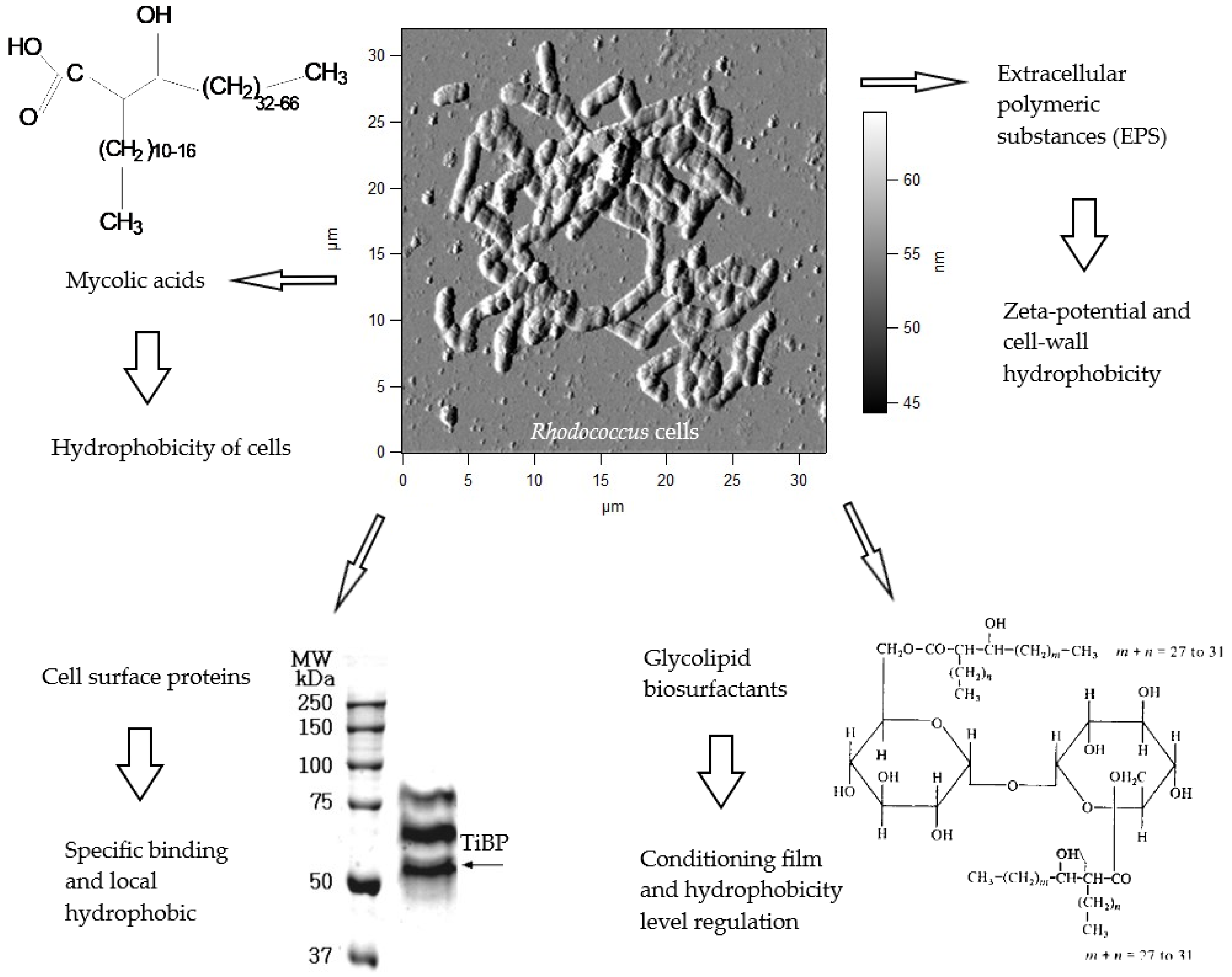
- Bendinger, B.; Rijnaarts, H.H.M.; Altendorf, K.; Zehnder, A.J.B. Physicochemical cell surface and adhesive properties of coryneform bacteria related to the presence and chain length of mycolic acids. Appl. Environ. Microbiol. 1993, 59, 3973–3977. [Google Scholar] [PubMed]
- Iwabuchi, N.; Sunairi, M.; Anzai, H.; Morisaki, H.; Nakajima, M. Relationships among colony morphotypes, cell-surface properties and bacterial adhesion to substrata in Rhodococcus. Colloids Surf. B Biointerfaces 2003, 30, 51–60. [Google Scholar] [CrossRef]
- de Mesquita, L.M.S.; Lins, F.F.; Torem, M.L. Interaction of a hydrophobic bacterium strain in a hematite-quartz flotation system. Int. J. Miner. Process. 2003, 71, 31–44. [Google Scholar] [CrossRef]
- Li, H.; Zhang, M.; Li, C.; Yang, X.; Li, A.; Zhang, L. Bioadsorption behavior of Rhodococcus opacus on the surface of calcium and magnesium minerals. JOM 2015, 67, 382–390. [Google Scholar] [CrossRef]
- Lopez, L.Y.; Merma, A.G.; Torem, M.L.; Pino, G.H. Fundamental aspects of hematite flotation using the bacterial strain Rhodococcus ruber as bioreagent. Miner. Eng. 2015, 75, 63–69. [Google Scholar] [CrossRef]
- Pen, Y.; Zhang, Z.J.; Morales-García, A.L.; Mears, M.; Tarmey, D.S.; Edyvean, R.G.; Banwart, S.A.; Geoghegan, M. Effect of extracellular polymeric substances on the mechanical properties of Rhodococcus. Biochim. Biophys. Acta Biomembr. 2015, 1848, 518–526. [Google Scholar] [CrossRef]
- Lehocký, M.; Sťahel, P.; Koutný, M.; Čech, J.; Institoris, J.; Mráček, A. Adhesion of Rhodococcus sp. S3E2 and Rhodococcus sp. S3E3 to plasma prepared Teflon-like and organosilicon surfaces. J. Mater. Process. Technol. 2009, 209, 2871–2875. [Google Scholar] [CrossRef]
- Gertler, G.; Brudo, I.; Kenig, R.; Fleminger, G.A. TiO2-binding protein isolated from Rhodococcus strain GIN-1 (NCIMB 40340)—Purification, properties and potential applications. Materialwissenschaft und Werkstofftechnik 2003, 34, 1138–1144. [Google Scholar] [CrossRef]
- Fleminger, G.; Shabtai, Y. Direct and rapid analysis of the adhesion of bacteria to solid surfaces: Interaction of fluorescently labeled Rhodococcus strain GIN-1 (NCIMB 40340) cells with titanium-rich particles. Appl. Environ. Microbiol. 1995, 61, 4357–4361. [Google Scholar] [PubMed]
- Siegmann, A.; Komarska, A.; Betzalel, Y.; Brudo, I.; Jindou, S.; Mor, G.; Fleminger, G. The titanium binding protein of Rhodococcus ruber GIN1 (NCIMB 40340) is a cell-surface homolog of the cytosolic enzyme dihydrolipoamide dehydrogenase. J. Mol. Recognit. 2009, 22, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Sharifzadeh, S.; Langsrud, S.; Heir, E.; Rickard, A.H. Coaggregation between Rhodococcus and Acinetobacter strains isolated from the food industry. Can. J. Microbiol. 2015, 61, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Podorozhko, E.A.; Lozinsky, V.I.; Ivshina, I.B.; Kuyukina, M.S.; Krivorutchko, A.B.; Philp, J.C.; Cunningham, C.J. Hydrophobised sawdust as a carrier for immobilisation of the hydrocarbon-oxidizing bacterium Rhodococcus ruber. Bioresour. Technol. 2008, 99, 2001–2008. [Google Scholar] [CrossRef]
- Kuyukina, M.; Ivshina, I.; Krivoruchko, A.; Podorozhko, E.; Lozinsky, V.; Cunningham, C.; Philp, J. Novel biocatalysts based on immobilized Rhodococcus cells for oil-contaminated water purification. J. Biotechnol. 2007, 2, S99–S100. [Google Scholar] [CrossRef]
- Nwankwoala, A.U.; Egiebor, N.O.; Nyavor, K. Enhanced biodegradation of methylhydrazine and hydrazine contaminated NASA wastewater in fixed-film bioreactor. Biodegradation 2001, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kuyukina, M.S.; Ivshina, I.B.; Rubtsova, E.V.; Ivanov, R.V.; Lozinsky, V.I. Adsorptive immobilization of rhodococcal cells in hydrophobized derivatives of wide-pore poly(acrylamide) cryogel. Appl. Biochem. Microbiol. 2011, 47, 158–164. [Google Scholar] [CrossRef]
- Lozinsky, V.I.; Galaev, I.Y.; Plieva, F.M.; Savina, I.N.; Jungvid, H.; Mattiasson, B. Polymeric cryogels as promising materials of biotechnological interest. Trends Biotechnol. 2003, 21, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Кuyukina, М.S.; Кrivoruchko, А.V. Immobilization of hydrocarbon-oxidating Rhodococcus as a factor for the strength of oil remediation. In Immobilized Cells: Biocatalysts and Processes; Publishing Center RIOR: Moscow, Russia, 2018; pp. 409–428. ISBN 978-5-369-02004-3. [Google Scholar]
- Adamczyk, Z.; Siwek, B.; Zembala, M.; Weroński, P. Kinetics of localized adsorption of colloid particles. Langmuir 1992, 8, 2605–2610. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Ivshina, I.B.; Serebrennikova, M.K.; Osipenko, M.A.; Nyashin, Y.I. Experimental and theoretical studies of the actinobacterial immobilization process in a column fluidized-bed bioreactor. Russ. J. Biomech. 2012, 16, 74–81. [Google Scholar]
- Kuyukina, M.S.; Ivshina, I.B.; Osipenko, M.A.; Nyashin, Y.I.; Tyulenyova, A.N.; Serebrennikova, M.K.; Krivoruchko, A.V. A kinetic model of bacterial cell immobilization process on the solid carrier. Russ. J. Biomech. 2007, 11, 76–84. [Google Scholar]
- Krivoruchko, A.V.; Iziumova, A.Y.; Kuyukina, M.S.; Plekhov, O.A.; Naimark, O.B.; Ivshina, I.B. Adhesion of Rhodococcus ruber IEGM 342 to polystyrene studied using contact and non-contact temperature measurement techniques. Appl. Microbiol. Biotechnol. 2018, 102, 8525–8536. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Wang, X.; Chen, S. Engineering of a xylose metabolic pathway in Rhodococcus strains. Appl. Environ. Microbiol. 2012, 78, 5483–5491. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.A.; Comba, S.; Arabolaza, A.; Gramajo, H.; Alvarez, H.M. Overexpression of a phosphatidic acid phosphatase type 2 leads to an increase in triacylglycerol production in oleaginous Rhodococcus strains. Appl. Microbiol. Biotechnol. 2015, 99, 2191–2207. [Google Scholar] [CrossRef] [PubMed]
- García-Hidalgo, J.; Hormigo, D.; Prieto, M.A.; Arroyo, M.; de la Mata, I. Extracellular production of Streptomyces exfoliatus poly(3-hydroxybutyrate) depolymerase in Rhodococcus sp. T104: Determination of optimal biocatalyst conditions. Appl. Microbiol. Biotechnol. 2012, 93, 1975–1988. [Google Scholar] [CrossRef]
- Iida, T.; Moteki, Y.; Nakamura, K.; Taguchi, K.; Otagiri, M.; Asanuma, M.; Dohmae, N.; Usami, R. Functional expression of three Rieske non-heme iron oxygenases derived from Actinomycetes in Rhodococcus species for investigation of their degradation capabilities of dibenzofuran and chlorinated dioxins. Biosci. Biotechnol. Biochem. 2009, 73, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Serebrennikova, M.K.; Kuyukina, M.S.; Krivoruchko, A.V.; Ivshina, I.B. Adaptation of coimmobilized Rhodococcus cells to oil hydrocarbons in a column bioreactor. Appl. Biochem. Microbiol. 2014, 50, 265–272. [Google Scholar] [CrossRef]
- Vinage, I.; von Rohr, P.R. Biological waste gas treatment with a modified rotating biological contactor. I. Control of biofilm growth and long-term performance. Bioprocess Biosyst. Eng. 2003, 26, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Vinage, I.; von Rohr, P.R. Biological waste gas treatment with a modified rotating biological contactor. II. Effect of operating parameters on process performance and mathematical modeling. Bioprocess Biosyst. Eng. 2003, 26, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Serebrennikova, M.K.; Golovina, E.E.; Kuyukina, M.S.; Ivshina, I.B. A consortium of immobilized rhodococci for oilfield wastewater treatment in a column bioreactor. Appl. Biochem. Microbiol. 2017, 53, 435–440. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Ivshina, I.B. An Oleophilic Preparation for Oil-Contaminated Soil Treatment. RF Patent 2180276, 10 March 2002. [Google Scholar]
- Ivshina, I.B.; Kostarev, S.M.; Kuyukina, M.S.; Zakshevskaya, L.V. Bioremediation Method for Soils Contaminated with Oil or Oil Products. RF Patent 2193464, 27 November 2002. [Google Scholar]
- Kuyukina, M.S.; Ivshina, I.B.; Ritchkova, M.I.; Philp, J.C.; Cunningham, C.J.; Christofi, N.; Kuyukina, M.S.; Ivshina, I.B.; Ritchkova, M.I.; Philp, J.C.; et al. Bioremediation of crude oil-contaminated soil using slurry-phase biological treatment and land farming techniques. Soil Sediment Contam. 2003, 12, 85–99. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V. Hydrocarbon-oxidizing bacteria and their potential in eco-biotechnology and bioremediation. In Microbial Resources; Kurtboke, I., Ed.; Elsevier Inc.: Oxford, UK, 2017; pp. 121–148. ISBN 978-0-12-804765-1. [Google Scholar]
- Banerjee, A.; Ghoshal, A.K. Biodegradation of phenol by calcium-alginate immobilized Bacillus cereus in a packed bed reactor and determination of the mass transfer correlation. J. Environ. Chem. Eng. 2016, 4, 1523–1529. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Liu, Y.-X.; Ren, R.-P.; Lv, Y.-K. An adsorption-release-biodegradation system for simultaneous biodegradation of phenol and ammonium in phenol-rich wastewater. Bioresour. Technol. 2016, 211, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Kurzbaum, E.; Raizner, Y.; Cohen, O.; Suckeveriene, R.Y.; Kulikov, A.; Hakimi, B.; Kruh, L.I.; Armon, R.; Farber, Y.; Menashe, O. Encapsulated Pseudomonas putida for phenol biodegradation: Use of a structural membrane for construction of a well-organized confined particle. Water Res. 2017, 121, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Kadri, T.; Magdouli, S.; Rouissi, T.; Brar, S.K. Ex-situ biodegradation of petroleum hydrocarbons using Alcanivorax borkumensis enzymes. Biochem. Eng. J. 2018, 132, 279–287. [Google Scholar] [CrossRef]
- Suganthi, S.H.; Murshid, S.; Sriram, S.; Ramani, K. Enhanced biodegradation of hydrocarbons in petroleum tank bottom oil sludge and characterization of biocatalysts and biosurfactants. J. Environ. Manag. 2018, 220, 87–95. [Google Scholar] [CrossRef]
- Morris, J.M.; Jin, S. Enhanced biodegradation of hydrocarbon-contaminated sediments using microbial fuel cells. J. Hazard. Mater. 2012, 213–214, 474–477. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Application | Short description of a Biocatalyst | Operating Conditions | Efficacy | Ref. |
|---|---|---|---|---|
| Treatment of wastewater containing phenol | A biocer: whole cells of R. ruber DSM 7512 embedded into ceramics (alumosilicate + SiO2) | Batch | Complete degradation of phenol (0.5 g/L) in 5 days; ≥ 5 cycles of re-using; stable for 12 months; stored at +4 °C for 6 months. | [70] |
| A bench-scale flow dynamic system | Degradation of 80–100% phenol (0.5 g/L) in 6–8 days. | |||
| Treatment of wastewater containing chlorophenols | Whole cells of R. rhodochrous DSM 6263 entrapped into κ-carrageenan mixed with Fe3O4 nanoparticles | Batch | Complete degradation of chlorophenols (0.25 mM) in 5 h; 6 cycles of re-using. | [96] |
| Treatment of wastewater and groundwater containing nitrophenols | Whole cells of R. erythropolis HL PM-1, SC 1 | Batch | Complete degradation of 2,4-dinitrophenol (0.06 g/L) in 6 h. | [85] |
| Whole cells of R. erythropolis HL PM-1 embedded into agar beads | A plug-flow bioreactor | Complete degradation of 2,4-dinitrophenol (0.005 g/L) without loss of activity for 14 days. | ||
| Treatment of wastewater containing phenol and hydrocarbons | Whole cells of R. wratislaviensis BN38 entrapped in hydroxypropylcellulose/ poly(N-isopropylacrylamide) cryogel, acclimated | Semicontinuous with a gradual increase of a contaminant concentration | Complete degradation of phenol (20 g/L in total) and n-hexadecane (20 g/L in total) in 5 days; more active than SC in 2.5 times. | [91] |
| Treatment of wastewater from phenolic resin manufacturing companies | Whole cells of R. erythropolis UPV-1 adsorbed on ceramic Biolite® beads, acclimated | A packed-bed two column bioreactor with aeration | The rate of phenol (0.1–0.8 g/L) degradation 0.4–18.0 g/(l∙day); degradation of 100% formaldehyde (0.2 g/L); stable for 50 days. | [13] |
| Treatment of wastewater from phenolic resin manufacturing companies | Whole cells of R. erythropolis UPV-1 adsorbed on diatomaceous earth, acclimated | An air-stirred bioreactor with clarifier | The rate of phenol (0.1–1.2 g/L) degradation 0.1–11.5 g/(l∙day); degradation of 100% formaldehyde (0.3–2.4 g/L). | [67] |
| Treatment of wastewater from paper and pulp industry | Whole cells of R. rhodochrous IEGM 107, SC | Batch | Complete degradation of dehydroabietic acid (0.5 g/L) in 9 days. | [38] |
| Treatment of wastewater containing pharma pollutants | Whole cells of R. rhodochrous IEGM 608 adsorbed on modified sawdust, acclimated | Batch | Complete degradation of drotaverine hydrochloride (0.2 g/L) in 25 days. | [40] |
| Treatment of wastewater containing toxic amides | Whole cells of R. rhodochrous NHB-2 entrapped in agar gel beads | A five-stage plug bioreactor | Degradation of 90–100% acrylamide, acetamide, or propionamide (100 mM) at 45 °C and pH 8.5 in 8 h. | [83] |
| Treatment of wastewater containing azo-dyes | Azoreductase AzoRo from R. opacus 1CP covalently bound to meso-cellular foams | Batch | 38 U/mg towards Methyl Red(25 μM) at pH 4; stable for 60 h. | [103] |
| Treatment of groundwater contaminated with cyanides | Whole cells of R. rhodochrous BX2 + B. mojavensis M1 adsorbed on granular activated carbon, acclimated | A fluidized bed bioreactor | Complete degradation of CN− (0.05 × 10−3–7.48 × 10−3 g/L) continuously over a 110-day steady-state operation period. | [100] |
| Treatment of groundwater contaminated with N-nitroso- dimethylamine | Whole cells of R. ruber ENV425, self-immobilized | A propane-fed membrane bioreactor | Degradation of > 99.95 % N-nitroso- dimethylamine (7.4 × 10−6–77.0 × 10−6 g/L) continuously over a 135-day steady-state operation period. | [102] |
| Treatment of wastewater containing crude oil | Whole cells of R. opacus IEGM 249 and R. ruber IEGM 615 co-immobilized on modified sawdust, acclimated | A column fluidized-bed bioreactor | Degradation of 70–90% alkanes and PAHs of crude oil (2–3%) in 21 days; 4–6 cycles of re-using. | [76,88,130] |
| Treatment of oilfield wastewater | Whole cells of R. opacus IEGM 249 and R. ruber IEGM 615 entrapped into cryo-PVA | A column bioreactor | Degradation of 80% crude oil (0.45 g/L) at strong (194 g/L) mineralization in 21 days. | [88,133] |
| Bioremediation of crude oil-contaminated soil | Whole cells of R. erythropolis IEGM 275 and R. ruber IEGM 231 entrapped into cryo-PVA | Model soil | Degradation of 45% crude oil (5%) in 14 months under dry conditions. | [93] |
| Bioremediation of crude oil-contaminated soil | Pure identified strains from the IEGM Collection, SC or adsorbed on hydrophobized sawdust | In situ; ex situ in a slurry bioreactor | Degradation of 80–90% crude oil (5–30%) in cold and temperate climate regions in a summer season. | [134,135,136,137] |
| Waste gas treatment | Whole cells of P. putida F1 and R. erythropolis PWD1 adsorbed on polypropylene disks | A rotating biological contactor | Degradation of 30–90 % toluene vapors (3–5 g/m3) under steady-state operation conditions for 400 days. | [131,132] |
| Biofuel production | Whole cells of R. jostii RHA1 pJAM2/RO00075 or pTip-QC2/RO00075, SC; R. opacus PD630 pJAM2/RO00075 or pTip-QC2/RO00075, SC | Batch | Lipid production 35–50% CDW 2 | [127] |
| Biofuel production | Whole cells of R. jostii XYLAB or R. opacus XYLAB, SC | Batch | Lipid production 53–68% CDW | [126] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krivoruchko, A.; Kuyukina, M.; Ivshina, I. Advanced Rhodococcus Biocatalysts for Environmental Biotechnologies. Catalysts 2019, 9, 236. https://doi.org/10.3390/catal9030236
Krivoruchko A, Kuyukina M, Ivshina I. Advanced Rhodococcus Biocatalysts for Environmental Biotechnologies. Catalysts. 2019; 9(3):236. https://doi.org/10.3390/catal9030236
Chicago/Turabian StyleKrivoruchko, Anastasiia, Maria Kuyukina, and Irena Ivshina. 2019. "Advanced Rhodococcus Biocatalysts for Environmental Biotechnologies" Catalysts 9, no. 3: 236. https://doi.org/10.3390/catal9030236
APA StyleKrivoruchko, A., Kuyukina, M., & Ivshina, I. (2019). Advanced Rhodococcus Biocatalysts for Environmental Biotechnologies. Catalysts, 9(3), 236. https://doi.org/10.3390/catal9030236