In this
Catalysts Comment Article, we will present our latest published results [
1] (Fish, R. H. et al.
J. Organometal. Chem. 2017
), that refutes the recently published data contention by Hollmann et al.
Catalysts,
2019, that our biomimetic 1,4-NADH co-factors do not provide chiral alcohols, from the enzymatic reduction of achiral ketones with HLADH containing <5% natural NAD
+; i.e., all the chiral alcohol activity comes from the <5% natural NAD
+ contained in the HLADH enzyme, that generates natural 1,4-NADH, and not the 1,4-NADH biomimics [
2].
In complete contrast, our use of the HLADH (<5% natural NAD
+) enzyme, with both the biomimetic NAD
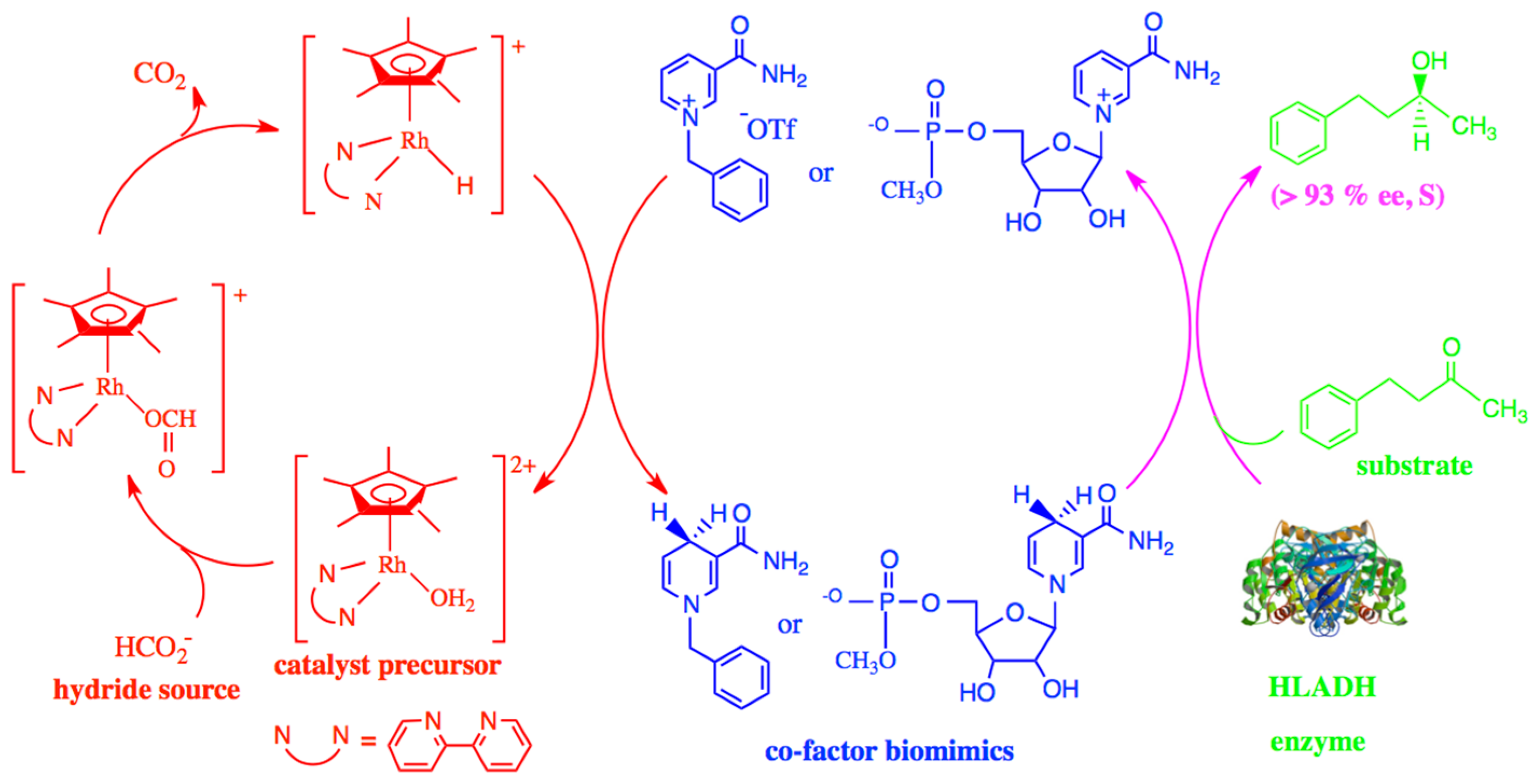
+ co-factors, N-1-benzylnicotinamide triflate,
1, and β-nicotinamide-5′-ribose methylphosphate,
2, and utilizing in situ formed [Cp*Rh(bpy)H](OTf) for biomimetic 1,4-NADH co-factor regeneration, provided chiral
S-alcohols, with high enantioselectivity, from reactions with achiral ketones. The seminal control experiment, amongst many, was the unequivocal finding that, in the absence of the biomimetic NAD
+ co-factors, and in the presence of HLADH (<5% natural NAD
+), the ketone substrate, and in situ formed [Cp*Rh(bpy)H](OTf),
only racemic alcohols were formed [
1], which we found emanated from the direct reaction of [Cp*Rh(bpy)H](OTf) with the ketones (Fish, R. H. et al.
J. Organometal. Chem. 2010) [
3]; this result implies that our HLADH enzyme contained ~0% natural NAD
+. The real contention is that Hollmann, et al., in their
Catalysts paper [
2], never cited our 2017 or 2010
J. Organometal. Chem. papers [
1,
3], or in fact, our paper on the mechanism for the chemoselective reduction of NAD
+ biomimetic co-factors with in situ formed [Cp*Rh(bpy)H](OTf), to provide biomimetic 1,4-NADH co-factors (Fish, R. H. et al.
Inorg. Chem. 2001) [
4]. This omission of key details, and failure to cite contradictory literature on the part of Hollmann and his co-workers in their 2019
Catalysts paper, is the focus of this
Catalysts Comment Article.
In our seminal paper,
Angew. Chem. Int. Ed., 2002, on the use of biomimetic co-factors in redox reactions at the interface of organometallic chemistry with biocatalysis, we did not provide our most important control experiments, as we should have, for unequivocal proof of concept [
5]. We found, serendipitously, that the simplified, biomimetic NAD
+ co-factors we were using to elucidate the chemoselective mechanism of the reaction of [Cp*Rh(bpy)H](OTf), to exclusively synthesize the biomimetic 1,4-NADH co-factors [
4], were also found to be surprisingly useful in biocatalysis enzyme reactions [
1,
5]. Thus, we decided to first test the efficacy of our biomimetic NAD
+ co-factors with the HLADH redox enzyme, again utilizing the in situ formed [Cp*Rh(bpy)H](OTf), as a co-factor regeneration reagent, in tandem with the HLADH redox biocatalysis. Since 2002 many enzymes, such as, Cytochrome P450 BM-3 and P450 CAM, as well as a HbpA enzyme that converts a phenol to a catechol, have also been shown to recognize our 1,4-NADH biomimics [
6,
7];
it should be noted that Hollmann is a co-author with this author, in [
7]
, from his PhD studies at the ETH, Zurich. We now come forward to 2014, and Hollmann’s review article, on biomimetic co-factors, which was published in
ACS Catalysis, 2014 [
8].
In his
ACS Catalysis review article, Hollmann et al. [
8] stated that our initial results on biomimetic NAD
+ co-factors, published in
Angew. Chem. Int. Ed., in 2002 [
5], were due to a contamination problem of the HLADH enzyme with natural NAD
+ (<5%), that we utilized in our studies. We then decided to publish a full account of our results, and describe our extensive control experiments, which clearly demonstrated the validity of our biomimetic NAD
+ and 1,4-NADH concept for HLADH enzyme recognition [
1,
5].
We describe, from the general procedure published in Reference 1, the biocatalytic reaction with the HLADH enzyme, NAD+ biomimics 1 and 2, and the in situ formed [Cp*Rh(bpy)H](OTf). Moreover, the most decisive control experiments (1 and 2) are also described:
The reported reaction conditions, such as the reactant ratio, reaction temperature, and the pre-catalyst amount, were obtained after numerous trial tests; however, the reaction conditions were not optimized. For reaction temperatures tested, from RT to 39 °C, while 30 °C exhibited the highest ee (85–99%) for the chiral alcohol products.
A general reaction procedure is given as follows, by using NAD
+ biomimic, N-1-benzylnicotinamide trilate,
1, and substrate, PhCH
2CH
2COCH
3, as an example: [Cp*Rh(bpy)(H
2O)](OTf)
2 (1.90 mg, 2.61 × 10
−3 mmol), sodium formate (17.80 mg, 261.69 × 10
−3 mmol), NAD
+ biomimic, N-1-benzylnicotinamide triflate,
1 (4.10 mg, 11.32 × 10
−3 mmol), and HLADH (23.80 mg, 10.23 units) were placed in a 10 mL Schlenk flask capped with a septum, and Schlenk techniques were carefully followed to deoxygenate the solid mixture. Potassium phosphate buffer (5 mL, 100 mM, pH = 7.02, de-O2) was added to give a clear solution, followed by the addition of the ketone substrate (12.5 mL, 83.41 × 10
−3 mmol, de-O
2) via syringe, respectively. Under a gentle N
2 pressure, the septum was removed, and the reaction flask was capped with a glass-stopper sealed with a small amount of grease and a metal clip. The stopcock of the reaction flask to N
2 gas was then shut, and the reaction flask was immediately shaken in a water-bath shaker at 30 °C, with the water level above the reaction mixture solution. The reaction was monitored by means of GC for 24 h (yield, 90%; 93% ee, see
Figure 1 and
Table 1) [
1].
Table 1, from Reference [
1], demonstrates that the biomimetic 1,4-NADH co-factors, N-1-benzyl-1,4-dihydronicotinamide,
3, and β-1,4-dihydronicotinamide-5′-ribose methylphosphate,
4, are as efficient in providing chiral alcohols from achiral ketones, as natural 1,4-NADH. Small amounts of a precipitate were observed in all reactions tested after 24 h. The SDS-PAGE analysis indicated the observed precipitate to be mainly the HLADH enzyme.
Since the purchased HLADH enzyme contained sodium phosphate, and a very small amount of NAD+, per the description by Sigma (combined impurities, <5% by weight), a series of control experiments were performed to verify the possible influence of the stated NAD+ impurity in HLADH, used in our biomimetic reduction reactions, for the synthesis of chiral alcohols.
Control experiment (1) [
1] using 2-pentanone and PhCH
2CH
2C(O)CH
3, as the ketone substrates, and under the same conditions as described in the General Procedure,
but in the absence of the NAD+ biomimic, β-nicotinamide-5′-ribose methylphosphate, 2, only racemic alcohols,
without a measurable ee in the products, could be detected in 24 h, both at RT (0% ee; TOF, 10 d
−1), and at 30 °C (0% ee; TOF, 14 d
−1), respectively
. The reaction rates were found to be comparable to the direct reduction of ketone substrates by precatalyst [Cp*Rh(bpy)(H
2O)](OTf)
2 with sodium formate, under similar conditions. For example, a yield of 33% (TOF, 11 d
−1) was obtained in the case of 2-pentanone for racemic, 2-pentanol, after 24 h at room temperature [
3].
Control experiment (2) [
1] under the same conditions as described in the Control Experiment (1), but using twice the amounts of the ketone substrates, at room temperature, provided only racemic 2-pentanol, without a measurable % ee, detected after 24 h. The yield was not determined. As a result of these control experiments, there appears to be no detectable NAD
+ in the HLADH used in our biomimetic reactions, and could be excluded as the responsible source for the obtained enantioselectivity in our biomimetic studies. That is, the recognition of the formed 1,4-NADH biomimics,
3 and
4, by HLADH, was clearly shown under the given conditions in our biomimetic system.
The Hollmann et al. Catalysts paper describes their results from a variety of ADH enzymes, which they contend does not provide chiral products from our biomimetic NAD+ co-factors, 1 and 2, even after the ADH enzymes are extracted to remove ~<5% of natural NAD+. However, we show unequivocally that when we remove the biomimetic co-factors, 1 and 2, from the tandem biocatalysis reactions with HLADH, ketone substrates, and in situ formed [Cp*Rh(bpy)H](OTf) for biomimetic 1,4-NADH regeneration, we only see racemic alcohols, which to our mind, dictates that our samples of HLADH have ~ 0% natural NAD+.
Thus, we stand by our unequivocal control experiment results. Finally, we also noted many errors in Hollmann et al.
Catalysts paper,
Figure 1: We used the triflate salt, N-1-benzylnicotinamide triflate, and not the chloride salt; the Cl
− anion has interfered with the formation of [Cp*Rh(bpy)H](OTf) [
1]. Furthermore, in their
Catalysts paper,
Figure 1B
, the NMN we utilized, as a totally water soluble biomimetic co-factor, was a methyl ester, β-1,4-dihydronicotinamide-5′-ribose methylphosphate,
4, and was not shown; only R = H, PO
32−. Again, lack of details, appears to negate some of their results, if they do not use the exact same co-factors and counterions, as we used, or a similar batch of HLADH, in their comparison experiments. Incidentally, biomimetic co-factor,
4, a difficult to prepare compound, could be more easily synthesized with our procedure, as shown in reference 4 of our
Inorg. Chem. 2001 paper [
4], also not referenced in the Hollmann et al.
Catalysts paper; it is essential to cap the phosphate group as a methyl ester, in order to avoid reaction with the precatalyst, [Cp*Rh(bpy)(H
2O)](OTf)
2.



{kind=link}



