Effect of Cholesterol and Ibuprofen on DMPC-β-Aescin Bicelles: A Temperature-Dependent Wide-Angle X-ray Scattering Study




{kind=link}
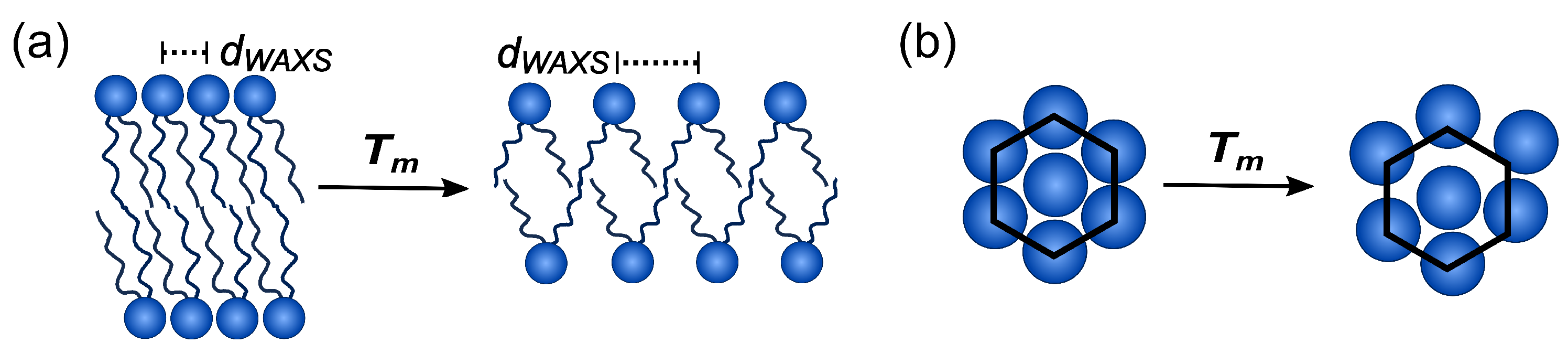{kind=link}
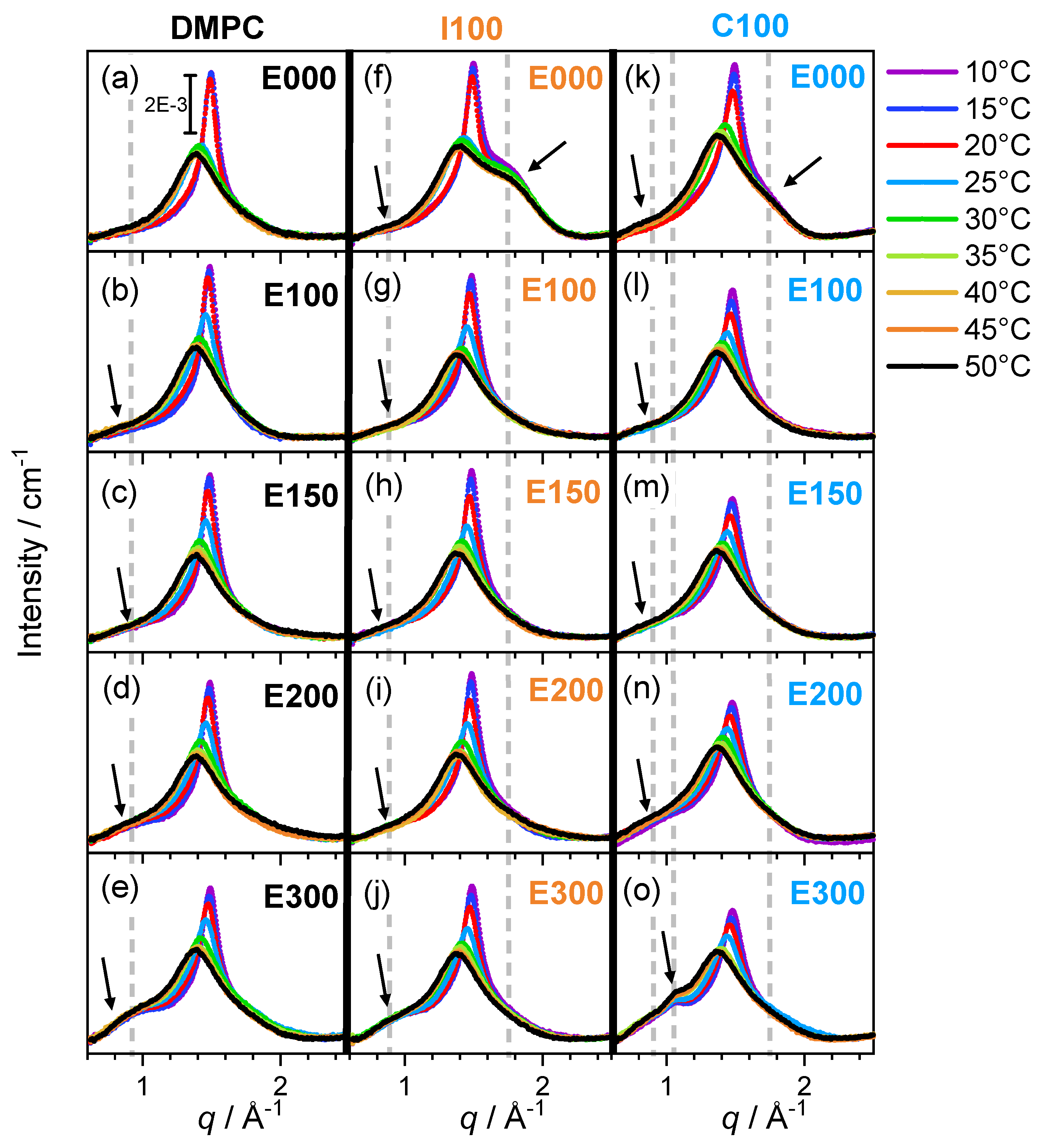{kind=link}
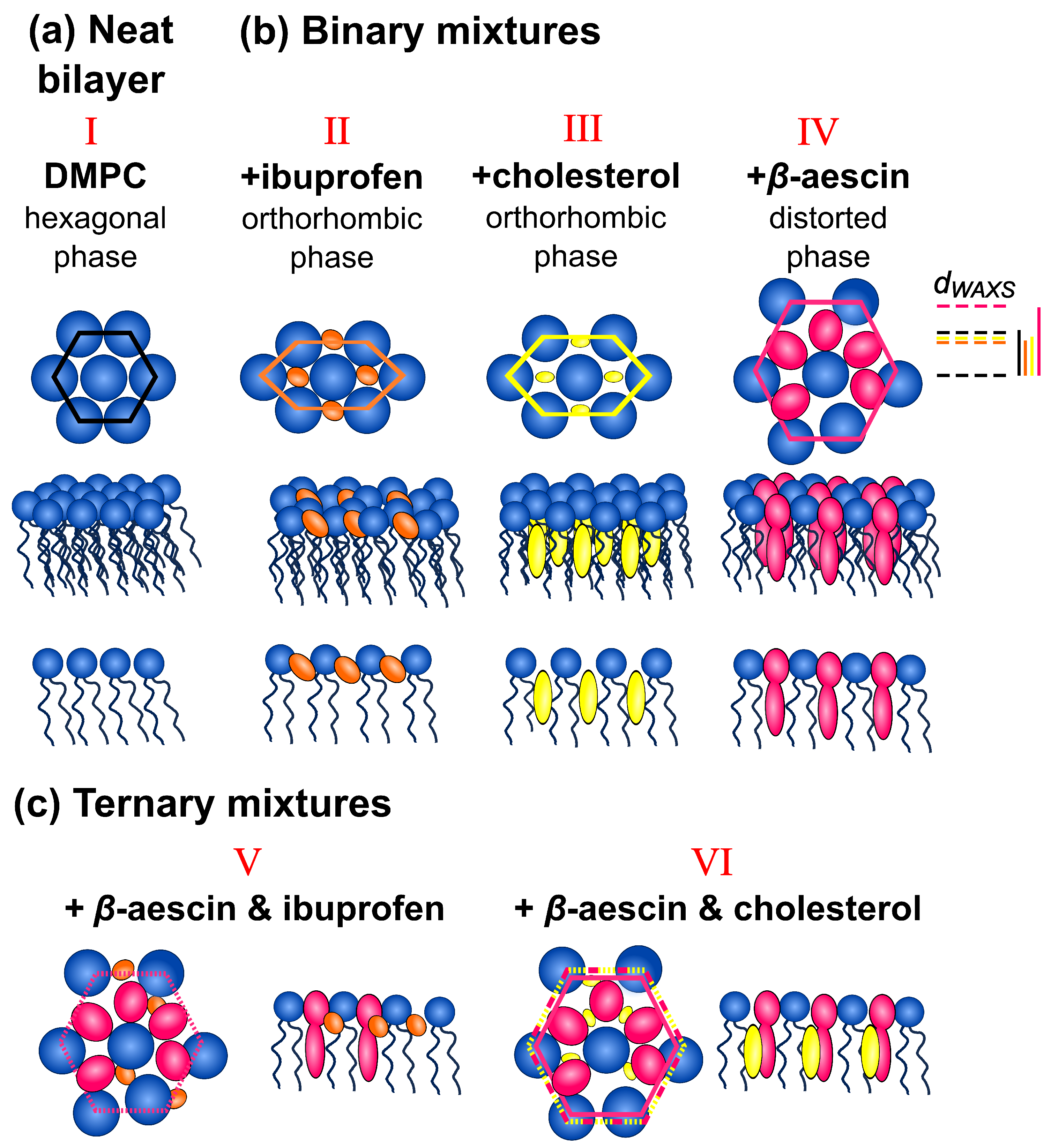{kind=link}
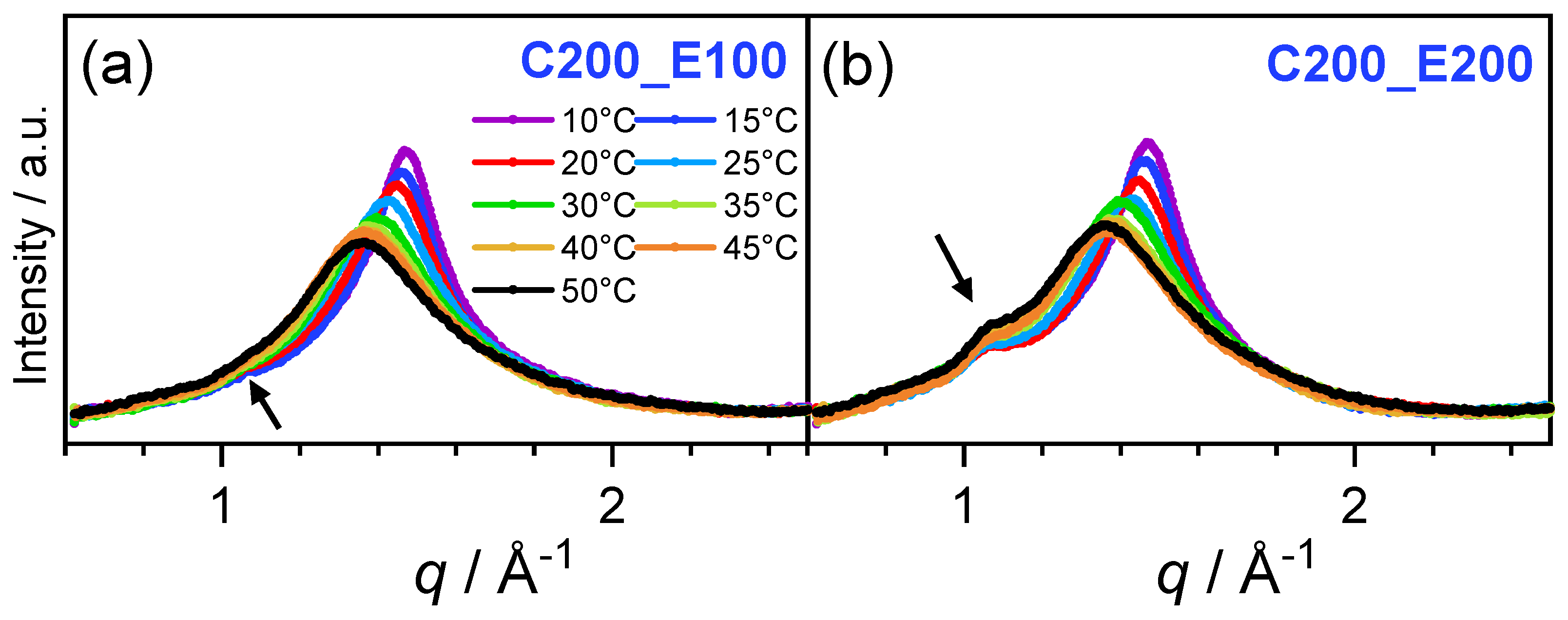{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation
2.3. Wide-Angle X-ray Scattering
3. Results and Discussion
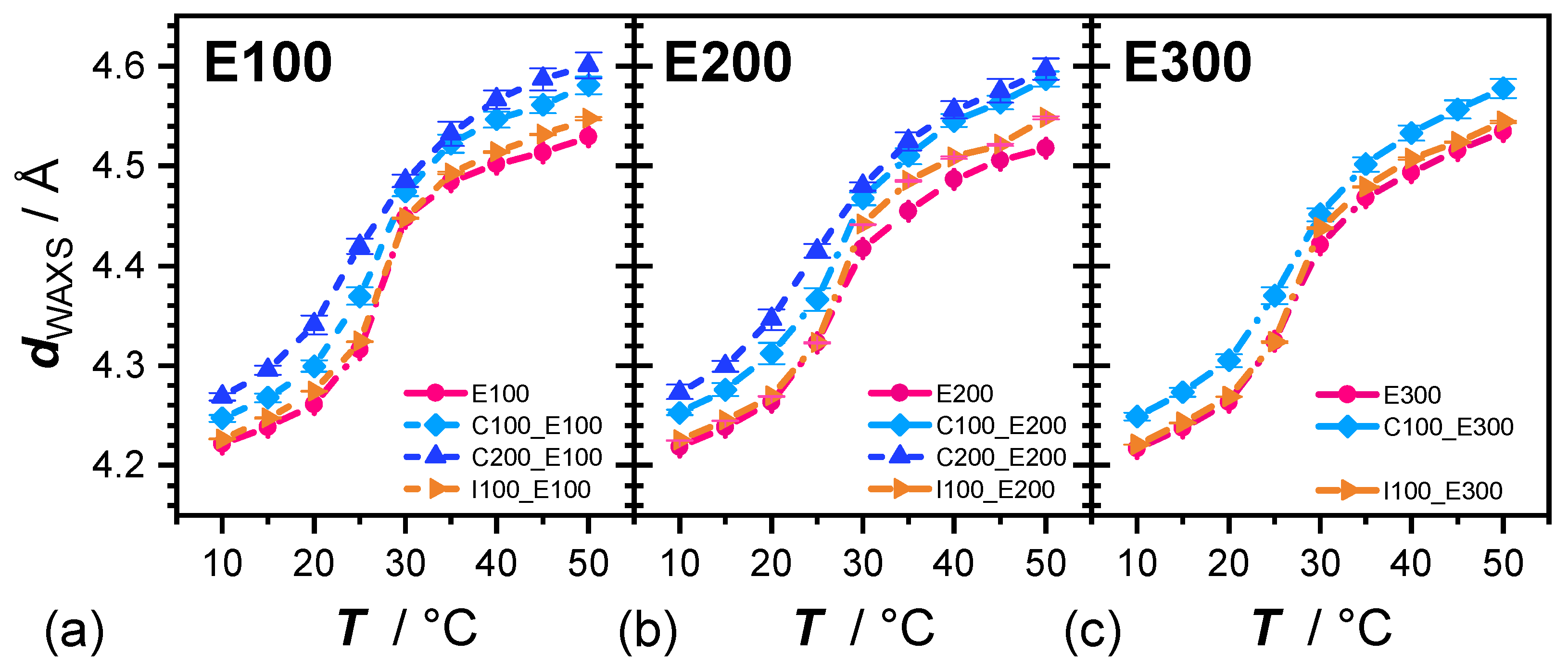
3.1. Shape of the WAXS Diffraction Patterns at Different Temperatures and Composition
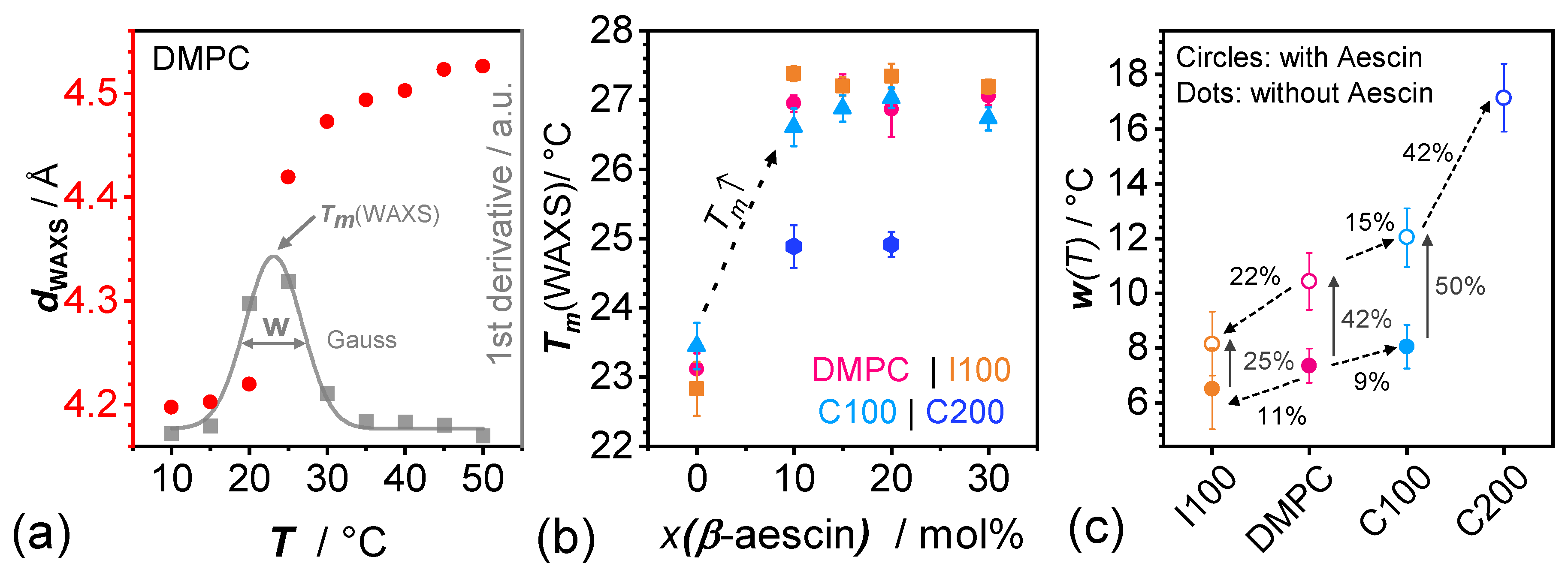
3.2. Evolution of with Temperature and Composition and Impact of the Composition on the Cooperative Phase Transition of the DMPC Bilayer
4. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vujic, Z.; Novovic, D.; Arsic, I.; Antic, D. Optimization of the extraction process of escin from dried seeds of Aesculus hippocastanum L. by Derringer’s desirability function. J. Anim. Plant Sci. 2013, 17, 2514–2521. [Google Scholar]
- Wilkinson, J.; Brown, A. Horse Chestnut—Aesculus hippocastanum: Potential Applications in Cosmetic Skin-care Products. Int. J. Cosmet. Sci. 1999, 21, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulff, G.; Tschesche, R. Über triterpene XXVI: Über die Struktur der Rosskastaniensaponnie (Aescin) und die Aglykone verwandter Glykoside. Tetrahedron 1969, 25, 415–436. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Murakami, T.; Matsuda, H.; Yamahara, J.; Murakami, N.; Kitagawa, I. Bioactive saponins and glycosides. III. Horse chestnut.(1): The structures, inhibitory effects on ethanol absorption, and hypoglycemic activity of escins Ia, Ib, IIa, IIb, and IIIa from the seeds of Aesculus hippocastanum L. Chem. Pharm. Bull. 1996, 44, 1454–1464. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, M.; Murakami, T.; Yamahara, J.; Matsuda, H. Bioactive saponins and glycosides. XII. Horse chestnut.(2): Structures of escins IIIb, IV, V, and VI and isoescins Ia, Ib, and V, acylated polyhydroxyoleanene triterpene oligoglycosides, from the seeds of horse chestnut tree (Aesculus hippocastanum L., Hippocastanaceae). Chem. Pharm. Bull. 1998, 46, 1764–1769. [Google Scholar] [PubMed] [Green Version]
- Horvath, E. Verfahren zur Gewinnung von β-Aescin-Reichen Extrakten. European Patent EP0298148B1, 18 September 1991. [Google Scholar]
- Khan, L.; Kifayatullah, Q.; Ahmad, K.; Arfan, M. Preparation of water-soluble aescin from aesculus-indica seeds. J. Chem. Soc. Pak. 1994, 16, 269–272. [Google Scholar]
- Sirtori, C.R. Aescin: Pharmacology, pharmacokinetics and therapeutic profile. Pharmacol. Res. 2001, 44, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Geisler, R.; Dargel, C.; Hellweg, T. The Biosurfactant β-Aescin: A Review on the Physico-Chemical Properties and Its Interaction with Lipid Model Membranes and Langmuir Monolayers. Molecules 2020, 25, 117. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, S.; Yang, Y.; Wang, D.; Gao, H. Natural barrigenol-like triterpenoids: A comprehensive review of their contributions to medicinal chemistry. Phytochemistry 2019, 161, 41–74. [Google Scholar] [CrossRef]
- Gallelli, L. Escin: A review of its anti-edematous, anti-inflammatory, and venotonic properties. Drug Des. Dev. Ther. 2019, 13, 3425–3437. [Google Scholar] [CrossRef] [Green Version]
- Patlolla, J.M.; Rao, C.V. Anti-inflammatory and Anti-cancer Properties of β-Escin, a Triterpene Saponin. Curr. Pharmacol. Rep. 2015, 1, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Song, Y.; Zhang, L.; Zhang, M.; Fu, F. Combined treatment with low dose prednisone and escin improves the anti-arthritic effect in experimental arthritis. Int. Immunopharmacol. 2016, 31, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Huang, Y.; Wu, C.; Du, Y.; Li, P.; Wang, M.; Wang, X.; Wang, Y.; Hao, Y.; Wang, T.; et al. Network Pharmacology Based Research on the Combination Mechanism Between Escin and Low Dose Glucocorticoids in Anti-rheumatoid Arthritis. Front. Pharmacol. 2019, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Dudek-Makuch, M.; Studzińska-Sroka, E. Horse chestnut—Efficacy and safety in chronic venous insufficiency: An overview. Rev. Bras. Farmacogn. 2015, 25, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Costantini, A. Escin in pharmaceutical oral dosage forms: Quantitative densitometric HPTLC determination. Il Farmaco 1999, 54, 728–732. [Google Scholar] [CrossRef]
- Pittler, M.H.; Ernst, E. Horse chestnut seed extract for chronic venous insufficiency. Cochrane Database Syst. Rev. 2012, 11, CD003230. [Google Scholar] [CrossRef]
- Kahn, S.R. The post-thrombotic syndrome. Hematol. Am. Soc. Hematol. Educ. Program 2010, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Kahn, S.R. The post thrombotic syndrome. Thromb. Res. 2011, 127, S89–S92. [Google Scholar] [CrossRef] [Green Version]
- Cheong, D.H.; Arfuso, F.; Sethi, G.; Wang, L.; Hui, K.M.; Kumar, A.P.; Tran, T. Molecular targets and anti-cancer potential of escin. Cancer Lett. 2018, 422, 1–8. [Google Scholar] [CrossRef]
- Fedotcheva, T.A.; Sheichenko, O.P.; Sheichenko, V.I.; Fedotcheva, N.I.; Shimanovskii, N.L. Preparation of a horse chestnut extract with a 50% content of escin and its actions on tumor cell proliferation and isolated mitochondria. Pharm. Chem. J. 2019, 53, 57–64. [Google Scholar] [CrossRef]
- Zhou, X.Y.; Fu, F.H.; Li, Z.; Dong, Q.J.; He, J.; Wang, C.H. Escin, a natural mixture of triterpene saponins, exhibits antitumor activity against hepatocellular carcinoma. Planta Medica 2009, 75, 1580–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harford-Wright, E.; Bidère, N.; Gavard, J. β-escin selectively targets the glioblastoma-initiating cell population and reduces cell viability. Oncotarget 2016, 7, 66865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dargel, C.; Geisler, R.; Hannappel, Y.; Kemker, I.; Sewald, N.; Hellweg, T. Self-assembly of the bio-surfactant aescin in solution: A small-angle X-ray scattering and Fluorescence study. Colloids Interfaces 2019, 3, 47. [Google Scholar] [CrossRef] [Green Version]
- Tcholakova, S.; Mustana, F.; Pagureva, N.; Golemanov, K.; Denkov, N.D.; Pelan, E.G.; Stoyanov, S.D. Role of surface properties for the kinetics of bubble Ostwald ripening in saponin-stabilized foams. Colloids Surf. A 2017, 534, 16–25. [Google Scholar] [CrossRef]
- Tsibranska, S.; Ivanova, A.; Tcholakova, S.; Denkov, N. Self-Assembly of Escin Molecules at the Air-Water Interface as Studied by Molecular Dynamics. Langmuir 2017, 33, 8330–8341. [Google Scholar] [CrossRef] [PubMed]
- Tsibranska, S.; Ivanova, A.; Tcholakova, S.; Denkov, N. Structure of dense adsorption layers of escin at the air-water interface studied by Molecular Dynamics simulations. Langmuir 2019, 35, 12876–12887. [Google Scholar] [CrossRef] [PubMed]
- Golemanov, K.; Tcholakova, S.; Denkov, N.; Pelan, E.; Stoyanov, S.D. Surface Shear Rheology of Saponin Adsorption Layers. Langmuir 2012, 28, 12071–12084. [Google Scholar] [CrossRef]
- Golemanov, K.; Tcholakova, S.; Denkov, N.; Pelana, E.; Stoyanov, S.D. The role of the hydrophobic phase in the unique rheological properties of saponin adsorption layers. Soft Matter 2014, 10, 7034–7044. [Google Scholar] [CrossRef]
- Sreij, R.; Dargel, C.; Moleiro, L.H.; Monroy, F.; Hellweg, T. Aescin Incorporation and Nanodomain Formation in DMPC Model Membranes. Langmuir 2017, 33, 14527. [Google Scholar] [CrossRef] [Green Version]
- Sreij, R.; Prevost, S.; Dargel, C.; Dattani, R.; Hertle, Y.; Wrede, O.; Hellweg, T. Interaction of the saponin aescin with ibuprofen in DMPC model membranes. Mol. Pharm. 2018, 15, 4446–4461. [Google Scholar] [CrossRef]
- Sreij, R.; Dargel, C.; Schweins, R.; Prevost, S.; Dattani, R.; Hellweg, T. Aescin-Cholesterol Complexes in DMPC Model Membranes: A DSC and Temperature-Dependent Scattering Study. Sci. Rep. 2019, 9, 5542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreij, R.; Dargel, C.; Hannappel, Y.; Jestin, J.; Prévost, S.; Dattani, R.; Wrede, O.; Hellweg, T. Temperature dependent self-organization of DMPC membranes promoted by intermediate amounts of the saponin aescin. Biochim. Biophys. Acta BBA Biomembr. 2019, 1861, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Sreij, R.; Dargel, C.; Geisler, P.; Hertle, Y.; Radulescu, A.; Pasini, S.; Perez, J.; Moleiro, L.H.; Hellweg, T. DMPC vesicle structure and dynamics in the presence of low amounts of the saponin aescin. Phys. Chem. Chem. Phys. 2018, 20, 9070–9083. [Google Scholar] [CrossRef] [PubMed]
- Boggara, M.B.; Faraone, A.; Krishnamoorti, R. Effect of pH and ibuprofen on the phospholipid bilayer bending modulus. J. Phys. Chem. B 2010, 114, 8061–8066. [Google Scholar] [CrossRef]
- Arriaga, L.R.; Lopez-Montero, I.; Monroy, F.; Orts-Gil, G.; Farago, B.; Hellweg, T. Stiffening Effect of Cholesterol on Disordered Lipid Phases: A Combined Neutron Spin Echo+Dynamic Light Scattering Analysis of the Bending Elasticity of Large Unilamellar Vesicles. Biophys. J. 2009, 96, 3629–3637. [Google Scholar] [CrossRef] [Green Version]
- Geisler, R.; Pedersen, M.C.; Hannappel, Y.; Schweins, R.; Prévost, S.; Dattani, R.; Arleth, L.; Hellweg, T. Aescin-induced conversion of gel-phase lipid membranes into bicelle-like lipid nanoparticles. Langmuir 2019, 35, 16244–16255. [Google Scholar] [CrossRef]
- Mahabir, S.; Small, D.; Li, M.; Wan, W.; Kucerka, N.; Littrell, K.; Katsaras, J.; Nieh, M.P. Growth kinetics of lipid-based nanodiscs to unilamellar vesicles: A time-resolved small angle neutron scattering (SANS) study. Biochim. Biophys. Acta BBA Biomembr. 2013, 1828, 1025–1035. [Google Scholar] [CrossRef]
- Genz, A.; Holzwarth, J.; Tsong, T. The influence of cholesterol on the main phase transition of unilamellar dipalmytoylphosphatidylcholine vesicles. A differential scanning calorimetry and iodine laser T-jump study. Biophys. J. 1986, 50, 1043–1051. [Google Scholar] [CrossRef] [Green Version]
- Boggara, M.B.; Krishnamoorti, R. Small-angle neutron scattering studies of phospholipid NSAID adducts. Langmuir 2010, 26, 5734–5745. [Google Scholar] [CrossRef]
- de Groot, C.; Müsken, M.; Müller-Goymann, C.C. Novel Colloidal Microstructures of β-Escin and the Liposomal Components Cholesterol and DPPC. Planta Medica 2018, 84, 1219–1227. [Google Scholar] [CrossRef]
- Bombardelli, E.; Patri, G.; Pozzi, R. Complexes of Aescin, β-Sitosterol or Cholesterol, and Phospholipids and Pharmaceutical Compositions Containing Them. U.S. Patent 5118671, 2 June 1992. [Google Scholar]
- Marsh, D. Lateral order in gel, subgel and crystalline phases of lipid membranes: Wide-angle X-ray scattering. Chem. Phys. Lipids 2012, 165, 59–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tristam-Nagle, S.; Liu, Y.; Legleiter, J.; Nagle, J.F. Structure of Gel and Phase DMPC and Determined by X-Ray and Diffraction. Biophys. J. 2002, 83, 3324–3335. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, T.; Sztucki, M.; Van Vaerenbergh, P.; Léonardon, J.; Gorini, J.; Claustre, L.; Sever, F.; Morse, J.; Boesecke, P. A multipurpose instrument for time-resolved ultra-small-angle and coherent X-ray scattering. J. Appl. Crystallogr. 2018, 51, 1511–1524. [Google Scholar] [CrossRef] [PubMed]
- Boesecke, P. Reduction of two-dimensional small- and wide-angle X-ray scattering data. J. Appl. Crystallogr. 2007, 40, 423–427. [Google Scholar] [CrossRef]
- Gestetner, B.; Assa, Y.; Henis, Y.; Tencer, M.; Rotman, Y.; Birk, Y.; Bondi, A. Interaction of lucerne saponins with sterols. Biochim. Biophys. Acta 1972, 270, 181–187. [Google Scholar] [CrossRef]
- Moreno, M.M.; Garidel, P.; Suwalsky, M.; Howe, J.; Brandenburg, K. The membrane-activity of Ibuprofen, Diclofenac, and Naproxen: A physico-chemical study with lecithin phospholipids. Biochim. Biophys. Acta BBA Biomembr. 2009, 1788, 1296–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maghsoudi, H.; Hallajzadeh, J.; Rezaeipour, M. Evaluation of the effect of polyphenol of escin compared with ibuprofen and dexamethasone in synoviocyte model for osteoarthritis: An in vitro study. Clin. Rheumatol. 2018, 37, 2471–2478. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geisler, R.; Prévost, S.; Dattani, R.; Hellweg, T. Effect of Cholesterol and Ibuprofen on DMPC-β-Aescin Bicelles: A Temperature-Dependent Wide-Angle X-ray Scattering Study. Crystals 2020, 10, 401. https://doi.org/10.3390/cryst10050401
Geisler R, Prévost S, Dattani R, Hellweg T. Effect of Cholesterol and Ibuprofen on DMPC-β-Aescin Bicelles: A Temperature-Dependent Wide-Angle X-ray Scattering Study. Crystals. 2020; 10(5):401. https://doi.org/10.3390/cryst10050401
Chicago/Turabian StyleGeisler, Ramsia, Sylvain Prévost, Rajeev Dattani, and Thomas Hellweg. 2020. "Effect of Cholesterol and Ibuprofen on DMPC-β-Aescin Bicelles: A Temperature-Dependent Wide-Angle X-ray Scattering Study" Crystals 10, no. 5: 401. https://doi.org/10.3390/cryst10050401
APA StyleGeisler, R., Prévost, S., Dattani, R., & Hellweg, T. (2020). Effect of Cholesterol and Ibuprofen on DMPC-β-Aescin Bicelles: A Temperature-Dependent Wide-Angle X-ray Scattering Study. Crystals, 10(5), 401. https://doi.org/10.3390/cryst10050401