
Protein Dynamics and Time Resolved Protein Crystallography at Synchrotron Radiation Sources: Past, Present and Future


Abstract
:1. Introduction
2. Some Important Considerations for Time-Resolved Crystallography Experiments

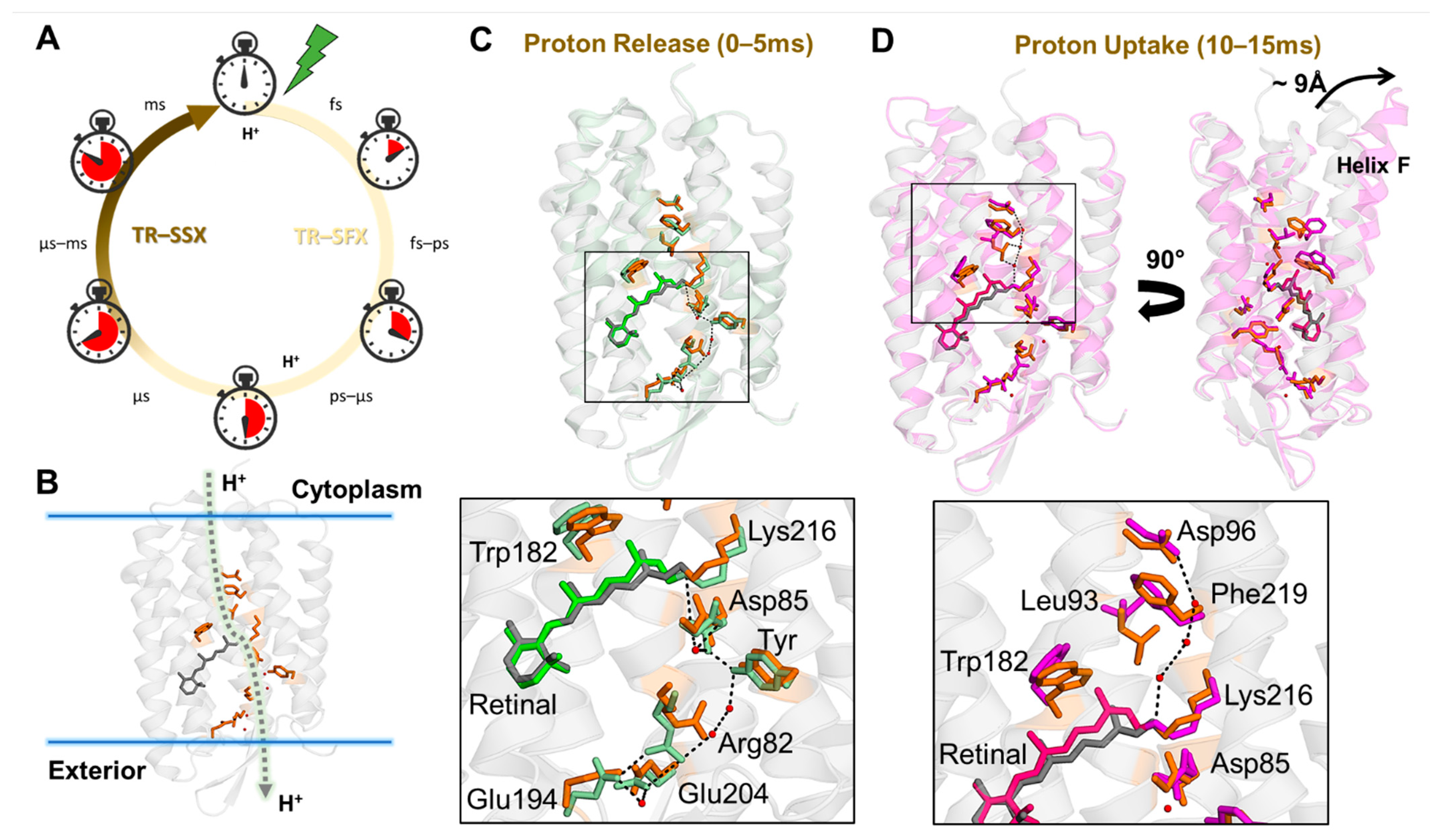{kind=link}
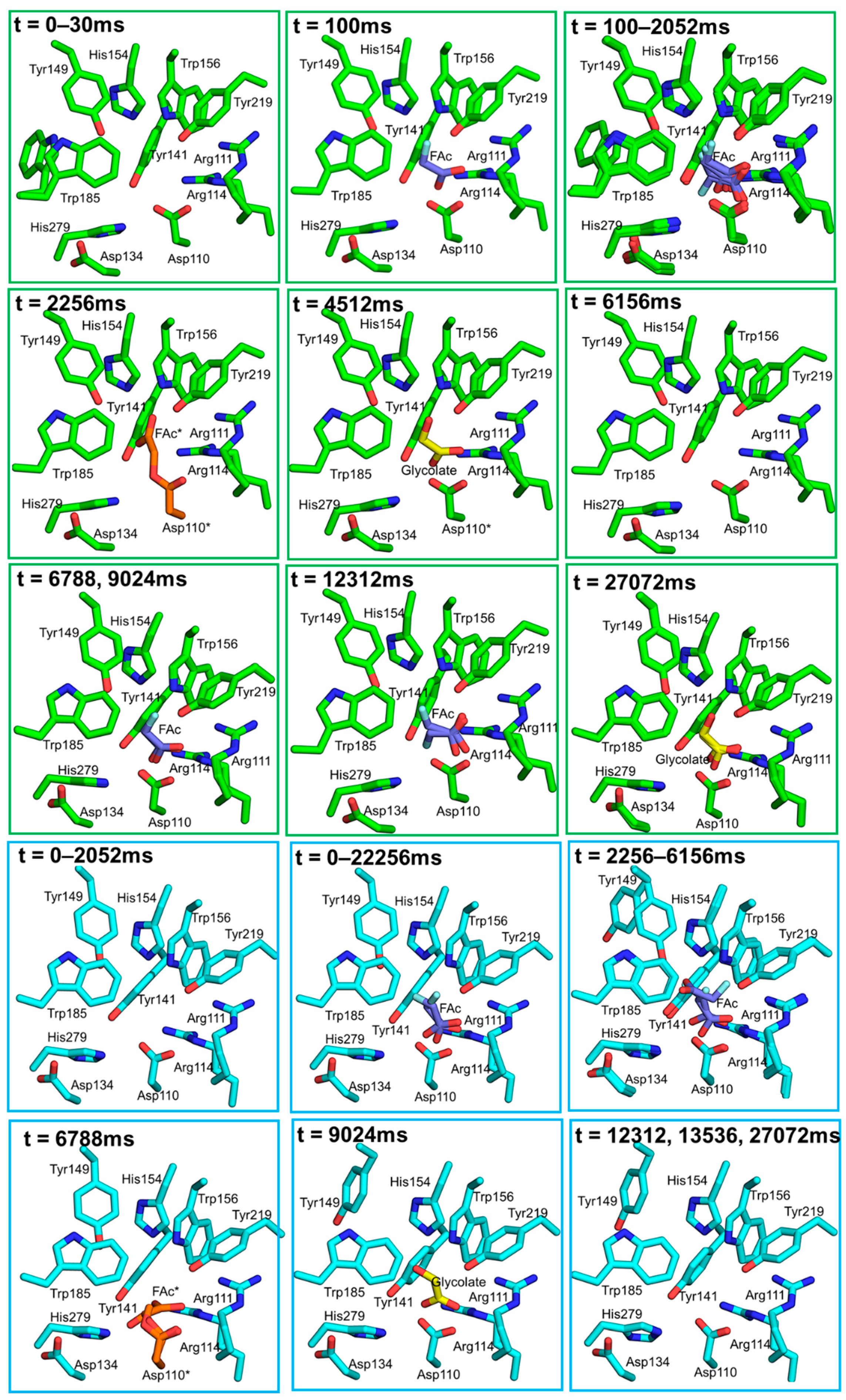{kind=link}
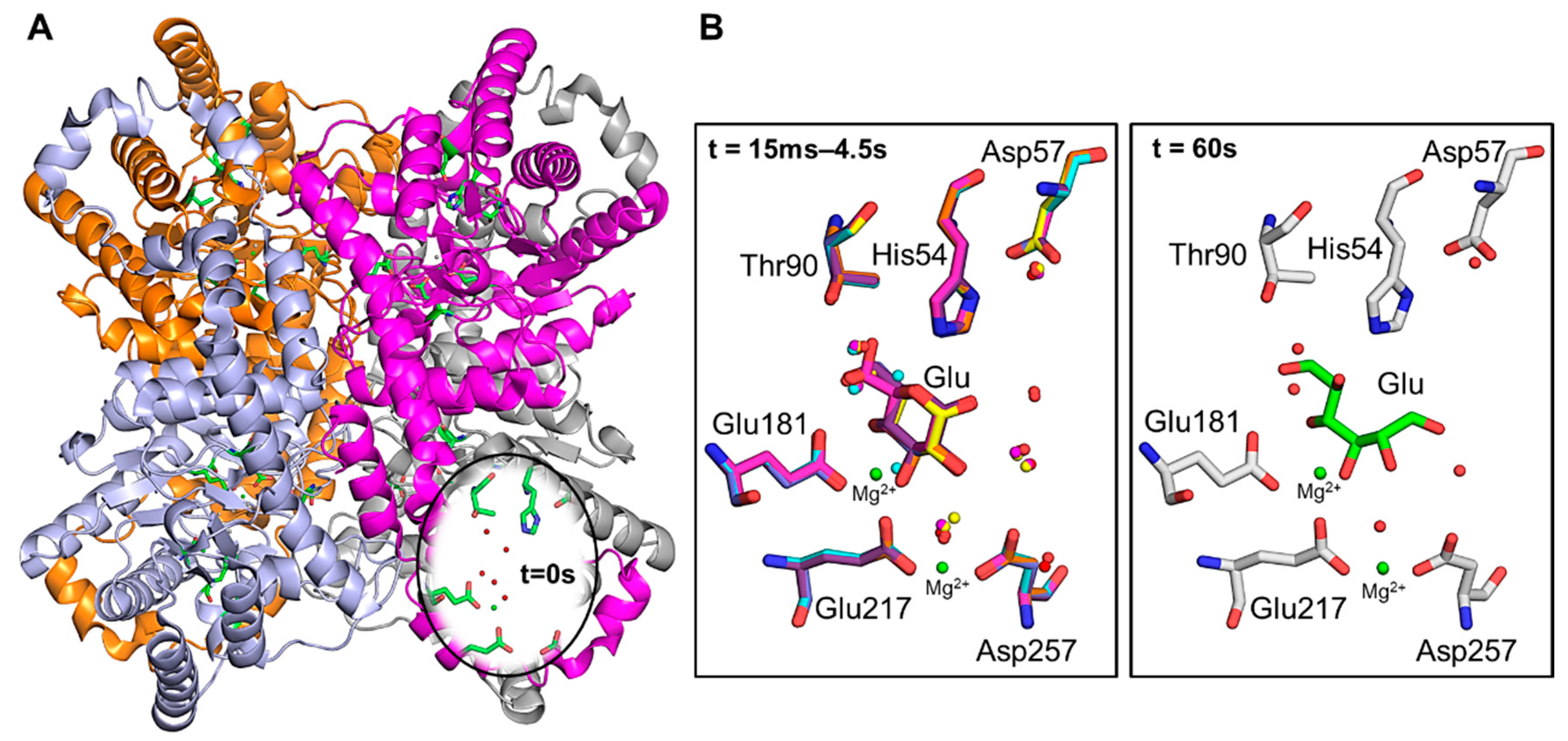{kind=link}
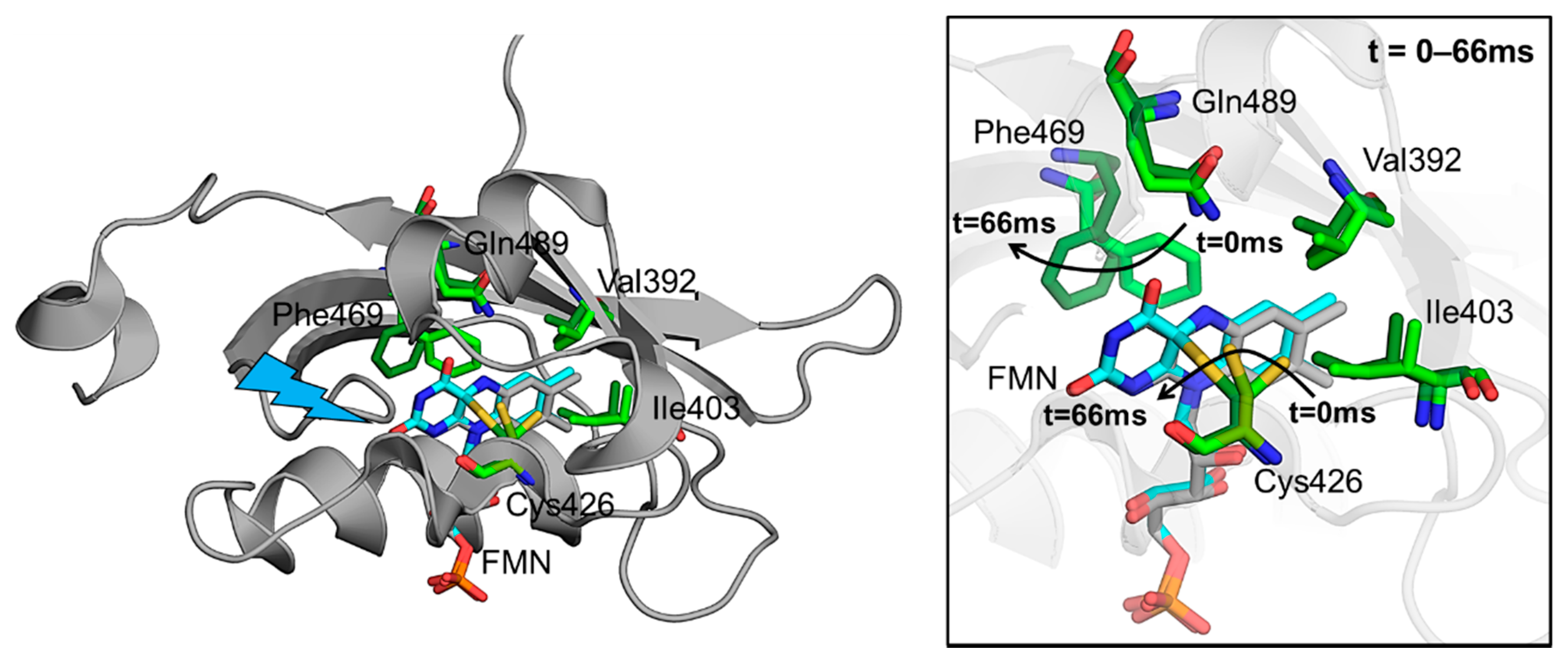{kind=link}
| Exp. # | Machine | Beamline | Delivery Method | Protein/Substrate | Temp. | Resol. (Å) | PDB Code | Ref. |
|---|---|---|---|---|---|---|---|---|
| 1 | PETRA III | P14 | Cryo-loop | Cathepsin B | 100 K | 3.00 | 4N4Z | [44] |
| 2 | PETRA III | P11 | Glass-capillary | Lysozyme | 293 K | 2.10 | 4O34 | [45] |
| 3 | CHESS | F1 | Microfluidic/chip | Glucose Isomerase (GI) | 293 K | 2.1 | Not deposited in PDB | [46] |
| 4 | ESRF | ID13 | Silicon nitride chip | Lysozyme | 293 K | 1.95 | 4WL7 | [47] |
| 5 | SLS | X10SA | High-viscosity injector | Lysozyme | 293 K | 1.90 | 4RLM | [35] |
| 6 | ESRF | ID13 | High-viscosity injector | Bacteriorhodopsin (bR) | 293 K | 2.40 | 4X31 | [36] |
| 7 | APS | 23-ID-D | Silicon nitride chip | Lysozyme | 293 K | 1.55 | 4Z98 | [48] |
| 8 | SLS | X10SA | Cyclic olefin copolymer (COC) sandwich plates | Lysozyme | 100 K | 1.8 | 4XJD | [49] |
| Alginate Transporter (AlgE) | 293 K | 2.8 | 4XNI | |||||
| Petptide Transporter (PepT) | 293 K | 2.9 | 4XNK | |||||
| 9 | ESRF | ID29 | Cryo-loop | Lysozyme | 100 K | 1.6 | 5A3Z | [50] |
| Thermolysin | 1.3 | 5A3Y | ||||||
| Bacteriorhodopsin (bR) | 2.5 | 5A44 | ||||||
| Thaumatin | 1.2 | 5A47 | ||||||
| 10 | DLS | I24 | Silicon chip | Thaumatin | 293 K | 2.2 | Not deposited in PDB | [51] |
| Proteinase K | 2.1 | |||||||
| 11 | DLS | I24 | Silicon chip | Polyhedrin | 293 K | 1.5 | 4X35 | [52] |
| Lysozyme | 2.1 | 4X3B | ||||||
| 12 | SLS | X06SA/X10SA | Cyclic olefin copolymer (COC) sandwich plates | Lysozyme | 100 K | 1.7 | 5D5C | [53] |
| Insuline | 1.5 | 5D53 | ||||||
| Alginate Transporter (AlgE) | 2.4 | 5D5D | ||||||
| Petptide Transporter (PepTst) | 2.4 | 5D58 | ||||||
| Diacylglycerol kinase (DgkA) | 2.8 | 5D56 | ||||||
| β2-adrenergic receptor (β2AR) | 2.5 | 5D5A | ||||||
| 13 | DLS | I03 | Silicon chip | Insulin | 293 K | 1.9 | 5FB6 | [54] |
| 14 | Spring-8 | BL41XU | Loop | Luciferin | 107 K | 1.6 | 5GX1 | [55] |
| 15 | APS | 23-ID-D | High-viscosity injector | Adenosine A2a Receptor (A2aAR) | 293 K | 3.1 | 5UVI | [56] |
| Phycocyanin | 3.2 | 5UVK | ||||||
| Lysozyme | 2.1 | 5UVJ | ||||||
| Proteinase K | 2.6 | 5UVL | ||||||
| 16 | APS | 14-ID-B | Chip | Phycocyanin | 293 K | 2.1 | 5MJP | [57] |
| Proteinase K | 2.2 | 5MJL | ||||||
| 17 | PETRA III | P11 | Tape-drive | Lysozyme / Chitotriose (CTO) | 293 K | 1.7 | 5JNP, 5NJQ, 5JNR, 5JNS | * [58] |
| 18 | SLS | X06SA | High-viscosity injector | Adenosine A2a Receptor (A2aAR) | 293 K | 2.1 | 5NLX | [59] |
| Lysozyme | 1.5 | 5NMJ | ||||||
| αβ-tubulin-darpin Complex (TD1) / Colchicine | 2.1 | 5NQT | ||||||
| Molybdenum Storage Protein (MOSTO) | 7.7 | 5OW5 | ||||||
| 19 | SLS | X10SA | High-viscosity injector | Thermolysin | 293 K | 2.3 | Not deposited in PDB | [60] |
| Glucose Isomerase (GI) | 2.0 | |||||||
| Lysozyme | 1.9 | |||||||
| Bacteriorhodopsin (bR) | 2.3 | |||||||
| 20 | ANSTO | MX2 | Micro-mesh | Bombyx mori cytoplasmic polyhedrosis virus 1 (BmCPV1) | 300 K | 1.51.9 | 5EXY, 5EXZ | [61] |
| 21 | ESRF | ID23-2 | Micro-mesh chips | Thaumatin | 100 K | 2.1 | 5FGT | [62] |
| Insulin | 1.7 | 9INS | ||||||
| 22 | PETRA III | P11 | High-viscosity injector | Proteinase K | 293 K | 1.9 | 6FJS | [63] |
| 23 | NSLS-II | FMX | Regular loops meshes | Proteinase K | 100 K | 2.0 | Not deposited in PDB | [64] |
| Trypsin | 1.5 | |||||||
| 24 | NSLS-II | FMX | Micro-well-mounts | Thaumatin | 100 K | 2.5 | 6C5Y | [65] |
| 25 | PETRA III | P14 | Microfluidic chips | Thaumatin | 293 K | 1.9 | 1LR2 | [66] |
| Glucose Isomerase (GI) | 1.7 | 4ZB2 | ||||||
| Thioredoxin | 3.0 | 4FYU | ||||||
| 26 | APS | 14-ID-B | Quartz chips | Ultraviolet-B receptor (UVR8) | 293 K | 2.0 | 6DD7 | [67] |
| Photolyase PhrB | 2.3 | 6DD6 | ||||||
| 27 | PETRA III | P14 | Hit And Return (HARE) chip | Fluoroacetate dehalogenase (FAcD) / Photocaged fluoroacetate | 293 K | 1.8 | 6GXH, 6FSX, 6GXD, 6GXT, 6GXF, 6GXL | * [68] |
| 28 | SLS | X06SA | Cyclic olefin copolymer (COC) sandwich plates | Petptide Transporter (PepTst) | 100 K | 2.6 | Not deposited in PDB | [69] |
| 29 | APS | 14-ID-B | High-viscosity injector | Adenosine A2a Receptor (A2aAR) | 293 K | 4.2 | 6MH8 | [70] |
| Proteinase K | 1.8 | 6MH6 | ||||||
| 30 | SLS | X06SA | High-viscosity injector | Bacteriorhodopsin (bR) | 293 K | 1.8–2.3 | 6RQP, 6RQO, 6RNJ,6RNH | * [29] |
| 31 | PETRA III | P14 | Liquid Application Method for time-resolved Analysis (LAMA) chip | Lysozyme / Chitotriose (CTO) | 293 K | 1.7–1.8 | 6RNB, 6RNC, 6QNB | * [71] |
| Xylose Isomerase (XI) / Glucose | 1.7–2.0 | 6QNH, 6RND, 6RNF, 6QNC, 6QNI, 6QNJ, 6QND | ||||||
| 32 | PETRA III | P11/P14 | Hit And Return (HARE) chip | Fluoroacetate dehalogenase (FAcD) / Photocaged fluoroacetate | 293 K | 1.7–1.9 | 6QHY, 8QHV, 6QHU, 6QHT, 6QHS, 6QHQ, 6QHP, 6QHW, 6QHZ, 6QI0, 6QI1, 6Q12, 6QI3 | * [72] |
| 33 | ESRF | ID13 | Microfluidic device | Lysozyme | 293 K | 2.1 | 6H79 | [73] |
| 34 | ESRF | ID30A-3 | Goniometer/loops | Phototropin-2 | 293 K | 2.2–2.9 | 6S45, 6S46 | *# [74] |
| 35 | ESRF | ID30A-3 | Microfluidic device | Aspartate α-decarboxylase (ADC) | 293 K | 2.0 | 6RXH | [75] |
| Lysozyme | 2.0 | 6RXI | ||||||
| 36 | SSRF | 18U1 | Microfluidic plate | Lysozyme | 293 K | 2.2 | 7C09 | [76] |
| Proteinase K | 2.1 | 7C0P | ||||||
| 37 | ESRF | ID13 | Silicon chip | Lysozyme | 293 K | 6Q8T, 6Q88 | [77] | |
| 38 | ALBA | XALOC | High-viscosity injector | Rhodopsin KR2 | 293 K | 2.5 | 6YC0 | * [78] |
| 39 | PETRA III | P14 | Loops | Glycine transporter 1 (GlyT1) | 100 K | 3.4–3.9 | 6ZBV, 6ZPL | [79] |
| 40 | Spring-8 | BL41XU | Polyimide mesh loop | Lysozyme | 293 K | 1.7–1.8 | 7CDK, 7CDM, 7CDN, 7CDP, 7CDQ, 7CDR, 7CDS, 7CDT, 7CDU | [80] |
| 41 | PETRA III | P14 | Hit And Return (HARE) chip | Fluoroacetate dehalogenase (FAcD) | 293 K | 1.7 | 7A42 | [81] |
| Myoblobin | 7A44 |
3. Time-Resolved Serial Femtosecond Crystallography (TR-SFX)
4. Time-Resolved Serial Synchrotron Crystallography (TR-SSX)
5. Examples of Time-Resolved Experiments at Synchrotrons
5.1. TR-SSX with Viscous Jets
5.2. TR-SSX with Fixed Targets
5.2.1. The HARE System
5.2.2. The LAMA System
5.3. TR-SSX with Hybrid Sample Delivery Devices
5.4. Goniometer-Based Time-Resolved Serial Oscillation Crystallography (TR-SOX)
6. In-Flow Microfluidics Devices for TR-SSX
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Blake, C.; Koenig, D.; Mair, G.; North, A.; Phillips, D.; Sarma, V. The three-dimensional structure of hen eggwhite lysozyme. Nature 1965, 206, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Van Den Bedem, H.; Fraser, J.S. Integrative, dynamic structural biology at atomic resolution—It’s about time. Nat. Methods 2015, 12, 307–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajdu, J.; Johnson, L.N. Progress with Laue Diffraction Studies on Protein and Virus Crystals. Biochemistry 1990, 29, 1669–1678. [Google Scholar] [CrossRef]
- Bourgeois, D.; Ursby, T.; Wulff, M.; Pradervand, C.; Legrand, A.; Schildkamp, W.; Labouré, S.; Srajer, V.; Teng, T.Y.; Roth, M.; et al. Feasibility and realization of single-pulse Laue diffraction on macromolecular crystals at ESRF. J. Synchrotron. Radiat. 1996, 3, 65–74. [Google Scholar] [CrossRef]
- Genick, U.K.; Borgstahl, G.E.O.; Ng, K.; Ren, Z.; Pradervand, C.; Burke, P.M.; Šrajer, V.; Teng, T.Y.; Schildkamp, W.; McRee, D.E.; et al. Structure of a protein photocycle intermediate by millisecond time- resolved crystallography. Science 1997, 275, 1471–1475. [Google Scholar] [CrossRef] [PubMed]
- Ihee, H.; Rajagopal, S.; Srajer, V.; Pahl, R.; Anderson, S.; Schmidt, M.; Schotte, F.; Anfinrud, P.A.; Wulff, M.; Moffat, K. Visualizing reaction pathways in photoactive yellow protein from nanoseconds to seconds. Proc. Natl. Acad. Sci. USA 2005, 102, 7145–7150. [Google Scholar] [CrossRef] [Green Version]
- Knapp, J.E.; Pahl, R.; Šrajer, V.; Royer, W.E. Allosteric action in real time: Time-resolved crystallographic studies of a cooperative dimeric hemoglobin. Proc. Natl. Acad. Sci. USA 2006, 103, 7649–7654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffat, K.; Szebenyi, B.D. X-ray Laue Diffraction from Protein Crystals. Science 1984, 223, 1423–1425. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Šrajer, V.; Knapp, J.E.; Royer, W.E. Cooperative macromolecular device revealed by meta-analysis of static and time-resolved structures. Proc. Natl. Acad. Sci. USA 2012, 109, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Nienhaus, K.; Pahl, R.; Krasselt, A.; Anderson, S.; Parak, F.; Nienhaus, G.U.; Šrajer, V. Ligand migration pathway and protein dynamics in myoglobin: A time-resolved crystallographic study on L29W MbCO. Proc. Natl. Acad. Sci. USA 2005, 102, 11704–11709. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Graber, T.; Henning, R.; Srajer, V. Five-dimensional crystallography. Acta Crystallogr. Sect. A Found. Crystallogr. 2010, 66, 198–206. [Google Scholar] [CrossRef]
- Bourgeois, D.; Royant, A. Advances in kinetic protein crystallography. Curr. Opin. Struct. Biol. 2005, 15, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Neutze, R.; Moffat, K. Time-resolved structural studies at synchrotrons and X-ray free electron lasers: Opportunities and challenges. Curr. Opin. Struct. Biol. 2012, 22, 651–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šrajer, V.; Schmidt, M. Watching proteins function with time-resolved x-ray crystallography. J. Phys. D Appl. Phys. 2017, 50, 373001. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, B.L. Trapping reaction intermediates in macromolecular crystals for structural analyses. Methods 2001, 24, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Schotte, F.; Lim, M.; Jackson, T.A.; Smirnov, A.V.; Soman, J.; Olson, J.S.; Phillips, G.N.; Wulff, M.; Anfinrud, P.A. Watching a protein as it functions with 150-ps time-resolved x-ray crystallography. Science 2003, 300, 1944–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schotte, F.; Cho, H.S.; Kaila, V.R.I.; Kamikubo, H.; Dashdorj, N.; Henry, E.R.; Graber, T.J.; Henning, R.; Wulff, M.; Hummer, G.; et al. Watching a signaling protein function in real time via 100-ps time-resolved Laue crystallography. Proc. Natl. Acad. Sci. USA 2012, 109, 19256–19261. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.O.; Lee, J.H.; Kim, J.; Schmidt, M.; Moffat, K.; Šrajer, V.; Ihee, H. Volume-conserving trans-cis isomerization pathways in photoactive yellow protein visualized by picosecond X-ray crystallography. Nat. Chem. 2013, 5, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Tenboer, J.; Basu, S.; Zatsepin, N.; Pande, K.; Milathianaki, D.; Frank, M.; Hunter, M.; Boutet, S.; Williams, G.J.; Koglin, J.E.; et al. Time-resolved serial crystallography captures high-resolution intermediates of photoactive yellow protein. Science 2014, 346, 1242–1246. [Google Scholar] [CrossRef] [Green Version]
- Pande, K.; Hutchison, C.D.M.; Groenhof, G.; Aquila, A.; Robinson, J.S.; Tenboer, J.; Basu, S.; Boutet, S.; DePonte, D.P.; Liang, M.; et al. Femtosecond structural dynamics drives the trans/cis isomerization in photoactive yellow protein. Science 2016, 352, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Kupitz, C.; Basu, S.; Grotjohann, I.; Fromme, R.; Zatsepin, N.A.; Rendek, K.N.; Hunter, M.S.; Shoeman, R.L.; White, T.A.; Wang, D.; et al. Serial time-resolved crystallography of photosystem II using a femtosecond X-ray laser. Nature 2014, 513, 261–265. [Google Scholar] [CrossRef]
- Kern, J.; Tran, R.; Alonso-Mori, R.; Koroidov, S.; Echols, N.; Hattne, J.; Ibrahim, M.; Gul, S.; Laksmono, H.; Sierra, R.G.; et al. Taking snapshots of photosynthetic water oxidation using femtosecond X-ray diffraction and spectroscopy. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kern, J.; Chatterjee, R.; Young, I.D.; Fuller, F.D.; Lassalle, L.; Ibrahim, M.; Gul, S.; Fransson, T.; Brewster, A.S.; Alonso-Mori, R.; et al. Structures of the intermediates of Kok’s photosynthetic water oxidation clock. Nature 2018, 563, 421–425. [Google Scholar] [CrossRef]
- Suga, M.; Akita, F.; Sugahara, M.; Kubo, M.; Nakajima, Y.; Nakane, T.; Yamashita, K.; Umena, Y.; Nakabayashi, M.; Yamane, T.; et al. Light-induced structural changes and the site of O=O bond formation in PSII caught by XFEL. Nature 2017, 543, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Suga, M.; Akita, F.; Yamashita, K.; Nakajima, Y.; Ueno, G.; Li, H.; Yamane, T.; Hirata, K.; Umena, Y.; Yonekura, S.; et al. An oxyl/oxo mechanism for oxygen-oxygen coupling in PSII revealed by an x-ray free-electron laser. Science 2019, 366, 334–338. [Google Scholar] [CrossRef] [Green Version]
- Young, I.D.; Ibrahim, M.; Chatterjee, R.; Gul, S.; Fuller, F.; Koroidov, S.; Brewster, A.S.; Tran, R.; Alonso-Mori, R.; Kroll, T.; et al. No TiStructure of photosystem II and substrate binding at room temperaturetle. Nature 2016, 540, 453–457. [Google Scholar] [CrossRef] [Green Version]
- Nango, E.; Royant, A.; Kubo, M.; Nakane, T.; Wickstrand, C.; Kimura, T.; Tanaka, T.; Tono, K.; Song, C.; Tanaka, R.; et al. A three-dimensional movie of structural changes in bacteriorhodopsin. Science 2016, 354, 1552–1557. [Google Scholar] [CrossRef]
- Nogly, P.; Weinert, T.; James, D.; Carbajo, S.; Ozerov, D.; Furrer, A.; Gashi, D.; Borin, V.; Skopintsev, P.; Jaeger, K.; et al. Retinal isomerization in bacteriorhodopsin captured by a femtosecond x-ray laser. Science 2018, 361, eaat0094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, T.; Skopintsev, P.; James, D.; Dworkowski, F.; Panepucci, E.; Kekilli, D.; Furrer, A.; Brünle, S.; Mous, S.; Ozerov, D.; et al. Proton uptake mechanism in bacteriorhodopsin captured by serial synchrotron crystallography. Science 2019, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Moffat, K. Laue diffraction. Methods Enzym. 1997, 277, 433–447. [Google Scholar]
- Kupitz, C.; Olmos, J.L.; Holl, M.; Tremblay, L.; Pande, L.; Pandey, S.; Oberthür, D.; Hunter, M.; Liang, M.; Aquila, A.; et al. Structural enzymology using X-ray free electron lasers. Struct. Dyn. 2017, 4, 044003. [Google Scholar] [CrossRef]
- Olmos, J.L.; Pandey, S.; Martin-Garcia, J.M.; Calvey, G.; Katz, A.; Knoska, J.; Kupitz, C.; Hunter, M.S.; Liang, M.; Oberthuer, D.; et al. Enzyme intermediates captured “on the fly” by mix-and-inject serial crystallography. BMC Biol. 2018, 16, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagno, J.R.; Liu, Y.; Bhandari, Y.R.; Conrad, C.E.; Panja, S.; Swain, M.; Fan, L.; Nelson, G.; Li, C.; Wendel, D.R.; et al. Structures of riboswitch RNA reaction states by mix-and-inject XFEL serial crystallography. Nature 2017, 541, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Echelmeier, A.; Sonker, M.; Ros, A. Microfluidic sample delivery for serial crystallography using XFELs. Anal. Bioanal. Chem. 2019, 411, 6535–6547. [Google Scholar] [CrossRef]
- Botha, S.; Nass, K.; Barends, T.R.M.; Kabsch, W.; Latz, B.; Dworkowski, F.; Foucar, L.; Panepucci, E.; Wang, M.; Shoeman, R.L.; et al. Room-temperature serial crystallography at synchrotron X-ray sources using slowly flowing free-standing high-viscosity microstreams. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogly, P.; James, D.; Wang, D.; White, T.A.; Zatsepin, N.; Shilova, A.; Nelson, G.; Liu, H.; Johansson, L.; Heymann, M.; et al. Lipidic cubic phase serial millisecond crystallography using synchrotron radiation. IUCrJ 2015, 2, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Fromme, P.; Graves, W.S.; Martin-Garcia, J.M. Serial Femtosecond Crystallography: A Decade at the Forefront in Structural Biology. eLS 2020, 1–17. [Google Scholar] [CrossRef]
- Martin-Garcia, J.M.; Conrad, C.E.; Coe, J.; Roy-Chowdhury, S.; Fromme, P. Serial femtosecond crystallography: A revolution in structural biology. Arch. Biochem. Biophys. 2016, 602, 32–47. [Google Scholar] [CrossRef] [Green Version]
- Kirian, R.A.; White, T.A.; Holton, J.M.; Chapman, H.N.; Fromme, P.; Barty, A.; Lomb, L.; Aquila, A.; Maia, F.R.N.C.; Martin, A.V.; et al. Structure-factor analysis of femtosecond microdiffraction patterns from protein nanocrystals. Acta Crystallogr. Sect. A Found. Crystallogr. 2011, 67, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Kirian, R.A.; Wang, X.; Weierstall, U.; Schmidt, K.E.; Spence, J.C.H.; Hunter, M.; Fromme, P.; White, T.; Chapman, H.N.; Holton, J. Femtosecond protein nanocrystallography—data analysis methods. Opt. Express 2010, 18, 5713. [Google Scholar] [CrossRef] [PubMed]
- Barty, A.; Kirian, R.A.; Maia, F.R.N.C.; Hantke, M.; Yoon, C.H.; White, T.A.; Chapman, H. Cheetah: Software for high-throughput reduction and analysis of serial femtosecond X-ray diffraction data. J. Appl. Crystallogr. 2014, 47, 1118–1131. [Google Scholar] [CrossRef] [Green Version]
- White, T.A.; Mariani, V.; Brehm, W.; Yefanov, O.; Barty, A.; Beyerlein, K.R.; Chervinskii, F.; Galli, L.; Gati, C.; Nakane, T.; et al. Recent developments in CrystFEL. J. Appl. Crystallogr. 2016, 49, 680–689. [Google Scholar] [CrossRef] [Green Version]
- Mariani, V.; Morgan, A.; Yoon, C.H.; Lane, T.J.; White, T.A.; O’Grady, C.; Kuhn, M.; Aplin, S.; Koglin, J.; Barty, A.; et al. OnDA: Online data analysis and feedback for serial X-ray imaging. J. Appl. Crystallogr. 2016, 49, 1073–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gati, C.; Bourenkov, G.; Klinge, M.; Rehders, D.; Stellato, F.; Oberthür, D.; Yefanov, O.; Sommer, B.P.; Mogk, S.; Duszenko, M.; et al. Serial crystallography on in vivo grown microcrystals using synchrotron radiation. IUCrJ 2014, 1, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Stellato, F.; Oberthür, D.; Liang, M.; Bean, R.; Gati, C.; Yefanov, O.; Barty, A.; Burkhardt, A.; Fischer, P.; Galli, L.; et al. Room-temperature macromolecular serial crystallography using synchrotron radiation. IUCrJ 2014, 1, 204–212. [Google Scholar] [CrossRef]
- Heymann, M.; Opthalage, A.; Wierman, J.L.; Akella, S.; Szebenyi, D.M.E.; Gruner, S.M.; Fraden, S. Room-temperature serial crystallography using a kinetically optimized microfluidic device for protein crystallization and on-chip X-ray diffraction. IUCrJ 2014, 1, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coquelle, N.; Brewster, A.S.; Kapp, U.; Shilova, A.; Weinhausen, B.; Burghammer, M.; Colletier, J.-P. Raster-scanning serial protein crystallography using micro- and nano-focused synchrotron beams. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 1184–1196. [Google Scholar] [CrossRef] [Green Version]
- Murray, T.D.; Lyubimov, A.Y.; Ogata, C.M.; Vo, H.; Uervirojnangkoorn, M.; Brunger, A.T.; Berger, J.M. A high-transparency, micro-patternable chip for X-ray diffraction analysis of microcrystals under native growth conditions. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 1987–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-Y.; Olieric, V.; Ma, P.; Panepucci, E.; Diederichs, K.; Wang, M.; Caffrey, M. In meso in situ serial X-ray crystallography of soluble and membrane proteins. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 1238–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zander, U.; Bourenkov, G.; Popov, A.N.; de Sanctis, D.; Svensson, O.; McCarthy, A.A.; Round, E.; Gordeliy, V.; Mueller-Dieckmann, C.; Leonard, G.A. MeshAndCollect: An automated multi-crystal data-collection workflow for synchrotron macromolecular crystallography beamlines. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 2328–2343. [Google Scholar] [CrossRef] [Green Version]
- Mueller, C.; Marx, A.; Epp, S.W.; Zhong, Y.; Kuo, A.; Balo, A.R.; Soman, J.; Schotte, F.; Lemke, H.T.; Owen, R.L.; et al. Fixed target matrix for femtosecond time-resolved and in situ serial micro-crystallography. Struct. Dyn. 2015, 2, 54302. [Google Scholar] [CrossRef] [Green Version]
- Roedig, P.; Vartiainen, I.; Duman, R.; Panneerselvam, S.; Stübe, N.; Lorbeer, O.; Warmer, M.; Sutton, G.; Stuart, D.I.; Weckert, E.; et al. A micro-patterned silicon chip as sample holder for macromolecular crystallography experiments with minimal background scattering. Sci. Rep. 2015, 5, 10451. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Olieric, V.; Ma, P.; Howe, N.; Vogeley, L.; Liu, X.; Warshamanage, R.; Weinert, T.; Panepucci, E.; Kobilka, B.; et al. In meso in situ serial X-ray crystallography of soluble and membrane proteins at cryogenic temperatures. Acta Crystallogr. Sect. D Struct. Biol. 2016, 72, 93–112. [Google Scholar] [CrossRef] [Green Version]
- Roedig, P.; Duman, R.; Sanchez-Weatherby, J.; Vartiainen, I.; Burkhardt, A.; Warmer, M.; David, C.; Wagner, A.; Meents, A. Room-temperature macromolecular crystallography using a micro-patterned silicon chip with minimal background scattering. J. Appl. Crystallogr. 2016, 49, 968–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, K.; Yamashita, K.; Murai, T.; Nuemket, N.; Hirata, K.; Ueno, G.; Ago, H.; Nakatsu, T.; Kumasaka, T.; Yamamoto, M. Development of a dose-limiting data collection strategy for serial synchrotron rotation crystallography. J. Synchrotron. Radiat. 2017, 24, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Martin-Garcia, J.M.; Conrad, C.E.; Nelson, G.; Stander, N.; Zatsepin, N.A.; Zook, J.; Zhu, L.; Geiger, J.; Chun, E.; Kissick, D.; et al. Serial millisecond crystallography of membrane and soluble protein microcrystals using synchrotron radiation. IUCrJ 2017, 4, 439–454. [Google Scholar] [CrossRef]
- Meents, A.; Wiedorn, M.O.; Srajer, V.; Henning, R.; Sarrou, I.; Bergtholdt, J.; Barthelmess, M.; Reinke, P.Y.A.; Dierksmeyer, D.; Tolstikova, A.; et al. Pink-beam serial crystallography. Nat. Commun. 2017, 8, 1281. [Google Scholar] [CrossRef] [Green Version]
- Beyerlein, K.R.; Dierksmeyer, D.; Mariani, V.; Kuhn, M.; Sarrou, I.; Ottaviano, A.; Awel, S.; Knoska, J.; Fuglerud, S.; Jönsson, O.; et al. Mix-and-diffuse serial synchrotron crystallography. IUCrJ 2017, 4, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Weinert, T.; Olieric, N.; Cheng, R.; Brünle, S.; James, D.; Ozerov, D.; Gashi, D.; Vera, L.; Marsh, M.; Jaeger, K.; et al. Serial millisecond crystallography for routine room-temperature structure determination at synchrotrons. Nat. Commun. 2017, 8, 542. [Google Scholar] [CrossRef]
- Kovácsová, G.; Grünbein, M.L.; Kloos, M.; Barends, T.R.M.; Schlesinger, R.; Heberle, J.; Kabsch, W.; Shoeman, R.L.; Doak, R.B.; Schlichting, I. Viscous hydrophilic injection matrices for serial crystallography. IUCrJ 2017, 4, 400–410. [Google Scholar] [CrossRef] [Green Version]
- Boudes, M.; Garriga, D.; Fryga, A.; Caradoc-Davies, T.; Coulibaly, F. A pipeline for structure determination of in vivo -grown crystals using in cellulo diffraction. Acta Crystallogr. Sect. D Struct. Biol. 2016, 72, 576–585. [Google Scholar] [CrossRef] [Green Version]
- Foos, N.; Seuring, C.; Schubert, R.; Burkhardt, A.; Svensson, O.; Meents, A.; Chapman, H.N.; Nanao, M.H. X-ray and UV radiation-damage-induced phasing using synchrotron serial crystallography. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Botha, S.; Baitan, D.; Jungnickel, K.E.J.; Oberthür, D.; Schmidt, C.; Stern, S.; Wiedorn, M.O.; Perbandt, M.; Chapman, H.N.; Betzel, C. De novo protein structure determination by heavy-atom soaking in lipidic cubic phase and SIRAS phasing using serial synchrotron crystallography. IUCrJ 2018, 5, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Xu, W.; Shi, W.; Soares, A.; Jakoncic, J.; Myers, S.; Martins, B.; Skinner, J.; Liu, Q.; Bernstein, H.; et al. High-speed raster-scanning synchrotron serial microcrystallography with a high-precision piezo-scanner. J. Synchrotron. Radiat. 2018, 25, 1362–1370. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Fuchs, M.R.; Shi, W.; Skinner, J.; Berman, E.; Ogata, C.M.; Hendrickson, W.A.; McSweeney, S.; Liu, Q. Sample manipulation and data assembly for robust microcrystal synchrotron crystallography. IUCrJ 2018, 5, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Gicquel, Y.; Schubert, R.; Kapis, S.; Bourenkov, G.; Schneider, T.; Perbandt, M.; Betzel, C.; Chapman, H.N.; Heymann, M. Microfluidic Chips for In Situ Crystal X-ray Diffraction and In Situ Dynamic Light Scattering for Serial Crystallography. J. Vis. Exp. 2018, 57133. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Ayhan, M.; Bandara, S.; Bowatte, K.; Kumarapperuma, I.; Gunawardana, S.; Shin, H.; Wang, C.; Zeng, X.; Yang, X. Crystal-on-crystal chips for in situ serial diffraction at room temperature. Lab Chip. 2018, 18, 2246–2256. [Google Scholar] [CrossRef] [PubMed]
- Schulz, E.C.; Mehrabi, P.; Müller-Werkmeister, H.M.; Tellkamp, F.; Jha, A.; Stuart, W.; Persch, E.; De Gasparo, R.; Diederich, F.; Pai, E.F.; et al. The hit-and-return system enables efficient time-resolved serial synchrotron crystallography. Nat. Methods 2018, 15, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Kaminski, J.W.; Panepucci, E.; Huang, C.-Y.; Warshamanage, R.; Wang, M.; Wojdyla, J.A. Automated data collection and real-time data analysis suite for serial synchrotron crystallography. J. Synchrotron. Radiat. 2019, 26, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Martin-Garcia, J.M.; Zhu, L.; Mendez, D.; Lee, M.-Y.; Chun, E.; Li, C.; Hu, H.; Subramanian, G.; Kissick, D.; Ogata, C.; et al. High-viscosity injector-based pink-beam serial crystallography of microcrystals at a synchrotron radiation source. IUCrJ 2019, 6, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, P.; Schulz, E.C.; Agthe, M.; Horrell, S.; Bourenkov, G.; von Stetten, D.; Leimkohl, J.-P.; Schikora, H.; Schneider, T.R.; Pearson, A.R.; et al. Liquid application method for time-resolved analyses by serial synchrotron crystallography. Nat. Methods 2019, 16, 979–982. [Google Scholar] [CrossRef]
- Mehrabi, P.; Schulz, E.C.; Dsouza, R.; Müller-Werkmeister, H.M.; Tellkamp, F.; Miller, R.J.D.; Pai, E.F. Time-resolved crystallography reveals allosteric communication aligned with molecular breathing. Science 2019, 365, 1167–1170. [Google Scholar] [CrossRef]
- Monteiro, D.C.F.; Vakili, M.; Harich, J.; Sztucki, M.; Meier, S.M.; Horrell, S.; Josts, I.; Trebbin, M. A microfluidic flow-focusing device for low sample consumption serial synchrotron crystallography experiments in liquid flow. J. Synchrotron. Radiat. 2019, 26, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Aumonier, S.; Santoni, G.; Gotthard, G.; Von Stetten, D.; Leonard, G.A.; Royant, A. Millisecond time-resolved serial oscillation crystallography of a blue-light photoreceptor at a synchrotron. IUCrJ 2020, 7, 728–736. [Google Scholar] [CrossRef]
- Monteiro, D.C.F.; von Stetten, D.; Stohrer, C.; Sans, M.; Pearson, A.R.; Santoni, G.; van der Linden, P.; Trebbin, M. 3D-MiXD: 3D-printed X-ray-compatible microfluidic devices for rapid, low-consumption serial synchrotron crystallography data collection in flow. IUCrJ 2020, 7, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.Z.; Sun, B.; Yu, L.; Xiao, Q.J.; Wang, Z.J.; Chen, L.L.; Liang, H.; Wang, Q.S.; He, J.H.; Yin, D.C. A novel sample delivery system based on circular motion for: In situ serial synchrotron crystallography. Lab Chip. 2020, 20, 3888–3898. [Google Scholar] [CrossRef] [PubMed]
- de la Mora, E.; Coquelle, N.; Bury, C.S.; Rosenthal, M.; Holton, J.M.; Carmichael, I.; Garman, E.F.; Burghammer, M.; Colletier, J.-P.; Weik, M. Radiation damage and dose limits in serial synchrotron crystallography at cryo- and room temperatures. Proc. Natl. Acad. Sci. USA 2020, 117, 4142–4151. [Google Scholar] [CrossRef] [Green Version]
- Kovalev, K.; Astashkin, R.; Gushchin, I.; Orekhov, P.; Volkov, D.; Zinovev, E.; Marin, E.; Rulev, M.; Alekseev, A.; Royant, A.; et al. Molecular mechanism of light-driven sodium pumping. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Shahsavar, A.; Stohler, P.; Bourenkov, G.; Zimmermann, I.; Siegrist, M.; Guba, W.; Pinard, E.; Sinning, S.; Seeger, M.A.; Schneider, T.R.; et al. Structural insights into the inhibition of glycine reuptake. Nature 2021, 591, 677–681. [Google Scholar] [CrossRef]
- Hasegawa, K.; Baba, S.; Kawamura, T.; Yamamotob, M.; Kumasakaa, T. Evaluation of the data-collection strategy for room-Temperature micro-crystallography studied by serial synchrotron rotation crystallography combined with the humid air and glue-coating method. Acta Crystallogr. Sect. D Struct. Biol. 2021, 77, 300–312. [Google Scholar] [CrossRef]
- Mehrabi, P.; Bücker, R.; Bourenkov, G.; Ginn, H.M.; von Stetten, D.; Müller-Werkmeister, H.M.; Kuo, A.; Morizumi, T.; Eger, B.T.; Ou, W.L.; et al. Serial femtosecond and serial synchrotron crystallography can yield data of equivalent quality: A systematic comparison. Sci. Adv. 2021, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, J.M.; Dyer, D.H.; Scott, W.G.; Singer, P.; Sweet, R.M.; Jr, D.E.K.; Stoddard, B.L.; Lyon, S.; Bolduc, J.M.; Dyer, D.H.; et al. Mutagenesis and Laue Structures of Enzyme Intermediates: Isocitrate Dehydrogenase Published by: American Association for the Advancement of Science Mutagenesis and Laue Structures of Enzyme Intermediates: Isocitrate Dehydrogenase. Science 2021, 268, 1312–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddard, B.L.; Farber, G.K. Direct measurement of reactivity in the protein crystal. Structure 1995, 3, 991–996. [Google Scholar] [CrossRef] [Green Version]
- Helliwell, J.R.; Nieh, Y.P.; Habash, J.; Faulder, P.F.; Raftery, J.; Cianci, M.; Wulff, M.; Hädener, A. Time-resolved and static-ensemble structural chemistry of hydroxymethylbilane synthase. Faraday Discuss 2002, 122, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Ursby, T.; Weik, M.; Fioravanti, E.; Delarue, M.; Goeldner, M.; Bourgeois, D. Cryophotolysis of caged compounds: A technique for trapping intermediate states in protein crystals. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Stoddard, B.L.; Cohen, B.E.; Brubaker, M.; Mesecar, A.D.; Koshland, D.E. Millisecond laue structures of an enzyme-product complex using photocaged substrate analogs. Nat. Struct. Biol. 1998, 5, 891–897. [Google Scholar] [CrossRef]
- Schlichting, I.; Rapp, G.; John, J.; Wittinghofer, A.; Pai, E.F.; Goody, R.S. Biochemical and crystallographic characterization of a complex of c-Ha-ras p21 and caged GTP with flash photolysis. Proc. Natl. Acad. Sci. USA 1989, 86, 7687–7690. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M. Mix and Inject: Reaction Initiation by Diffusion for Time-Resolved Macromolecular Crystallography. Adv. Condens. Matter Phys. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Weierstall, U.; James, D.; Wang, C.; White, T.A.; Wang, D.; Liu, W.; Spence, J.C.H.; Bruce Doak, R.; Nelson, G.; Fromme, P.; et al. Lipidic cubic phase injector facilitates membrane protein serial femtosecond crystallography. Nat. Commun. 2014, 5, 3309. [Google Scholar] [CrossRef] [Green Version]
- Ishigami, I.; Lewis-Ballester, A.; Echelmeier, A.; Brehm, G.; Zatsepin, N.A.; Grant, T.D.; Coe, J.D.; Lisova, S.; Nelson, G.; Zhang, S.; et al. Snapshot of an oxygen intermediate in the catalytic reaction of cytochrome c oxidase. Proc. Natl. Acad. Sci. USA 2019, 116, 3572–3577. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Kumar, S. Microbial enzyme in food biotechnology. Enzym. Food Biotechnol. Prod. Appl. Futur. Prospect. 2018, 19–28. [Google Scholar] [CrossRef]
- Lavie, A.; Allen, K.N.; Petsko, G.A.; Ringe, D. X-ray Crystallographic Structures of d-Xylose Isomerase-Substrate Complexes Position the Substrate and Provide Evidence for Metal Movement during Catalysis. Biochemistry 1994, 33, 5469–5480. [Google Scholar] [CrossRef]
- Kovalevsky, A.; Fisher, Z.; Johnson, H.; Mustyakimov, M.; Waltman, M.J.; Langan, P. Macromolecular neutron crystallography at the Protein Crystallography Station (PCS). Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Blake, C.C.F.; Johnson, L.N.; Mair, G.A.; North, A.C.T.; Phillips, D.C.; Sarma, V.R. Crystallographic studies of the activity of hen egg-white lysozyme. Proc. R. Soc. L B Biol. Sci. 1967, 167, 378–388. [Google Scholar]
- Vocadlo, D.J.; Davies, G.J.; Laine, R.; Withers, S.G. Catalysis by hen egg-white lysozyme proceeds via a covalent intermediate. Nature 2001, 412, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Kaiserli, E.; Sullivan, S.; Jones, M.A.; Feeney, K.A.; Christie, J.M. Domain swapping to assess the mechanistic basis of Arabidopsis phototropin 1 receptor kinase activation and endocytosis by blue light. Plant. Cell 2009, 21, 3226–3244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasahara, M.; Swartz, T.E.; Olney, M.A.; Onodera, A.; Mochizuki, N.; Fukuzawa, H.; Asamizu, E.; Tabata, S.; Kanegae, H.; Takano, M.; et al. Photochemical properties of the flavin mononucleotide-binding domains of the phototropins from Arabidopsis, rice, and Chlamydomonas reinhardtii. Plant. Physiol. 2002, 129, 762–773. [Google Scholar] [CrossRef] [Green Version]
- Vanhaelewyn, L.; Schumacher, P.; Poelman, D.; Fankhauser, C.; Van Der Straeten, D.; Vandenbussche, F. REPRESSOR OF ULTRAVIOLET-B PHOTOMORPHOGENESIS function allows efficient phototropin mediated ultraviolet-B phototropism in etiolated seedlings. Plant. Sci. 2016, 252, 215–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, H.N.; Fromme, P.; Barty, A.; White, T.A.; Kirian, R.A.; Aquila, A.; Hunter, M.S.; Schulz, J.; DePonte, D.P.; Weierstall, U.; et al. Femtosecond X-ray protein nanocrystallography. Nature 2011, 470, 73–77. [Google Scholar] [CrossRef]
- Grünbein, M.L.; Kovacs, G.N. Sample delivery for serial crystallography at free-electron lasers and synchrotrons. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.; Boutet, S.; Carini, G.; Dubrovin, M.; Duda, B.; Fritz, D.; Haller, G.; Herbst, R.; Herrmann, S.; Kenney, C.; et al. The CSPAD megapixel x-ray camera at LCLS. In SPIE Optical Engineering + Applications; Moeller, S.P., Yabashi, M., Hau-Riege, S.P., Eds.; International Society for Optics and Photonics: Bellingham, WA, USA, 2012; p. 85040C. [Google Scholar]
- Denes, P.; Schmitt, B. Pixel detectors for diffraction-limited storage rings. J. Synchrotron. Radiat. 2014, 21, 1006–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hülsen, G.; Broennimann, C.; Eikenberry, E.F.; Wagner, A. Protein crystallography with a novel large-area pixel detector. J. Appl. Crystallogr. 2006, 39, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Allahgholi, A.; Becker, J.; Bianco, L.; Delfs, A.; Dinapoli, R.; Goettlicher, P.; Graafsma, H.; Greiffenberg, D.; Hirsemann, H.; Jack, S.; et al. AGIPD, a high dynamic range fast detector for the European XFEL. J. Instrum. 2015, 10, C01023. [Google Scholar] [CrossRef] [Green Version]
- Casanas, A.; Warshamanage, R.; Finke, A.D.; Panepucci, E.; Olieric, V.; Nöll, A.; Tampé, R.; Brandstetter, S.; Förster, A.; Mueller, M.; et al. EIGER detector: Application in macromolecular crystallography: Acta Crystallogr. Sect. D Struct. Biol. 2016, 72, 1036–1048. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Garcia, J.M. Protein Dynamics and Time Resolved Protein Crystallography at Synchrotron Radiation Sources: Past, Present and Future. Crystals 2021, 11, 521. https://doi.org/10.3390/cryst11050521
Martin-Garcia JM. Protein Dynamics and Time Resolved Protein Crystallography at Synchrotron Radiation Sources: Past, Present and Future. Crystals. 2021; 11(5):521. https://doi.org/10.3390/cryst11050521
Chicago/Turabian StyleMartin-Garcia, Jose M. 2021. "Protein Dynamics and Time Resolved Protein Crystallography at Synchrotron Radiation Sources: Past, Present and Future" Crystals 11, no. 5: 521. https://doi.org/10.3390/cryst11050521
APA StyleMartin-Garcia, J. M. (2021). Protein Dynamics and Time Resolved Protein Crystallography at Synchrotron Radiation Sources: Past, Present and Future. Crystals, 11(5), 521. https://doi.org/10.3390/cryst11050521