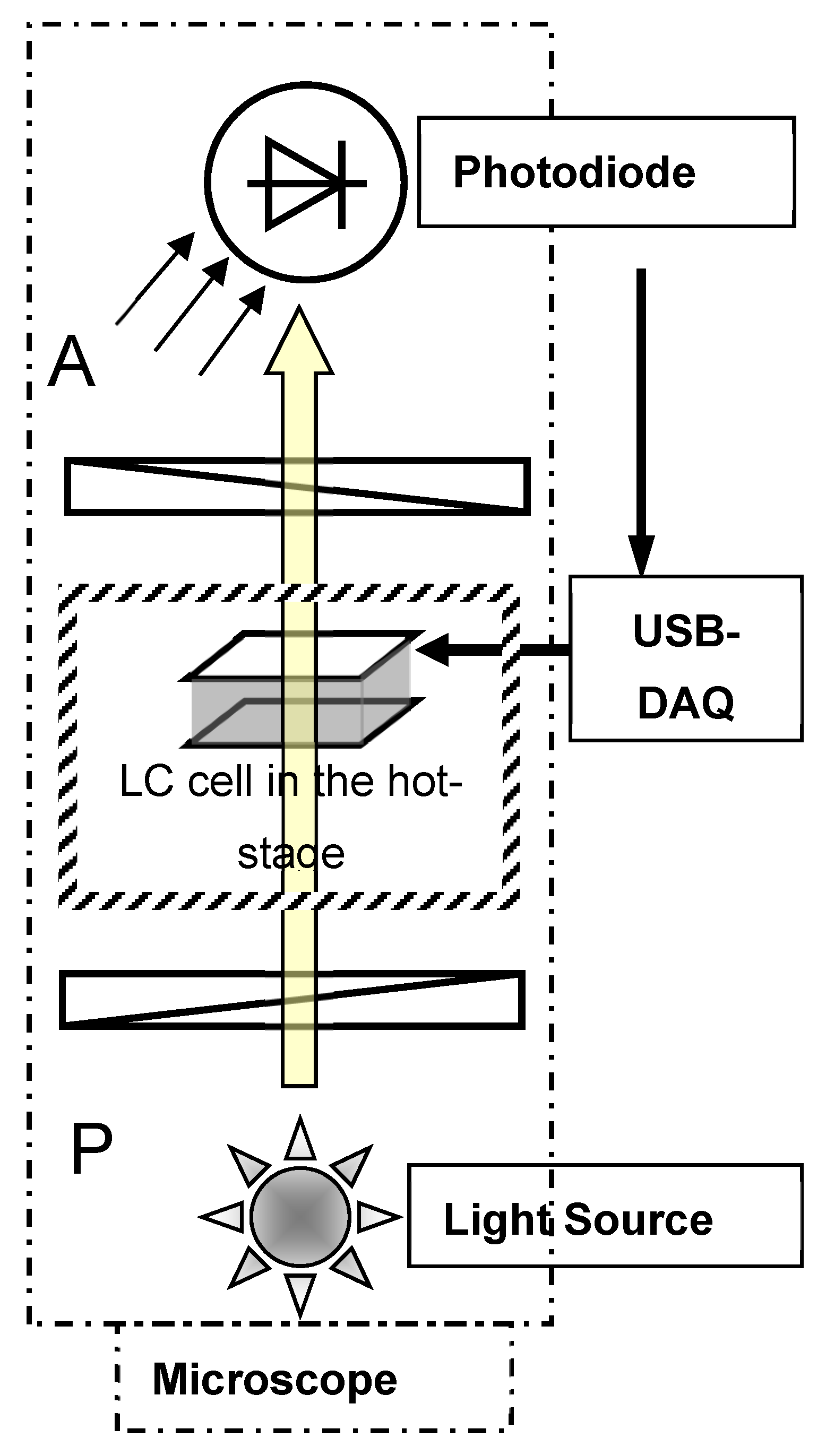
3.2. Electro-Optical Response
Results for the electro-optical experiment show that for temperatures lower than for a conventional nematic, the odd harmonics of the signal are of measurable amplitudes in the Nx phase [
16]. This is possible only if switching is polar in an LC cell. When the square waveform is applied, the signals for the fundamental and third harmonic continue to satisfy the condition V
1F = 3 V
3F, even when the frequency of the signal applied across the sample cell is as large as 2 kHz. This means that the switching time is at least as fast as the third harmonic of the applied field. The results of the experiment are a direct demonstration of the polar switching that could conceivably arise from the domain of a single chirality present in the field of view of the microscope. This, indeed, was an extremely interesting observation recorded in the experiment on the nematic phase formed from achiral molecules. The observation raised an important question for us to answer—how did we obtain the structure of a single chirality over the macroscopic area of a ‘parallel rubbed planar-aligned’ material of achiral molecules in an LC cell?
The domains of single chirality were found to vary in size and shape depending on the surface conditions of an LC cell, the temperature gradient, the cooling/heating rate during the N–N
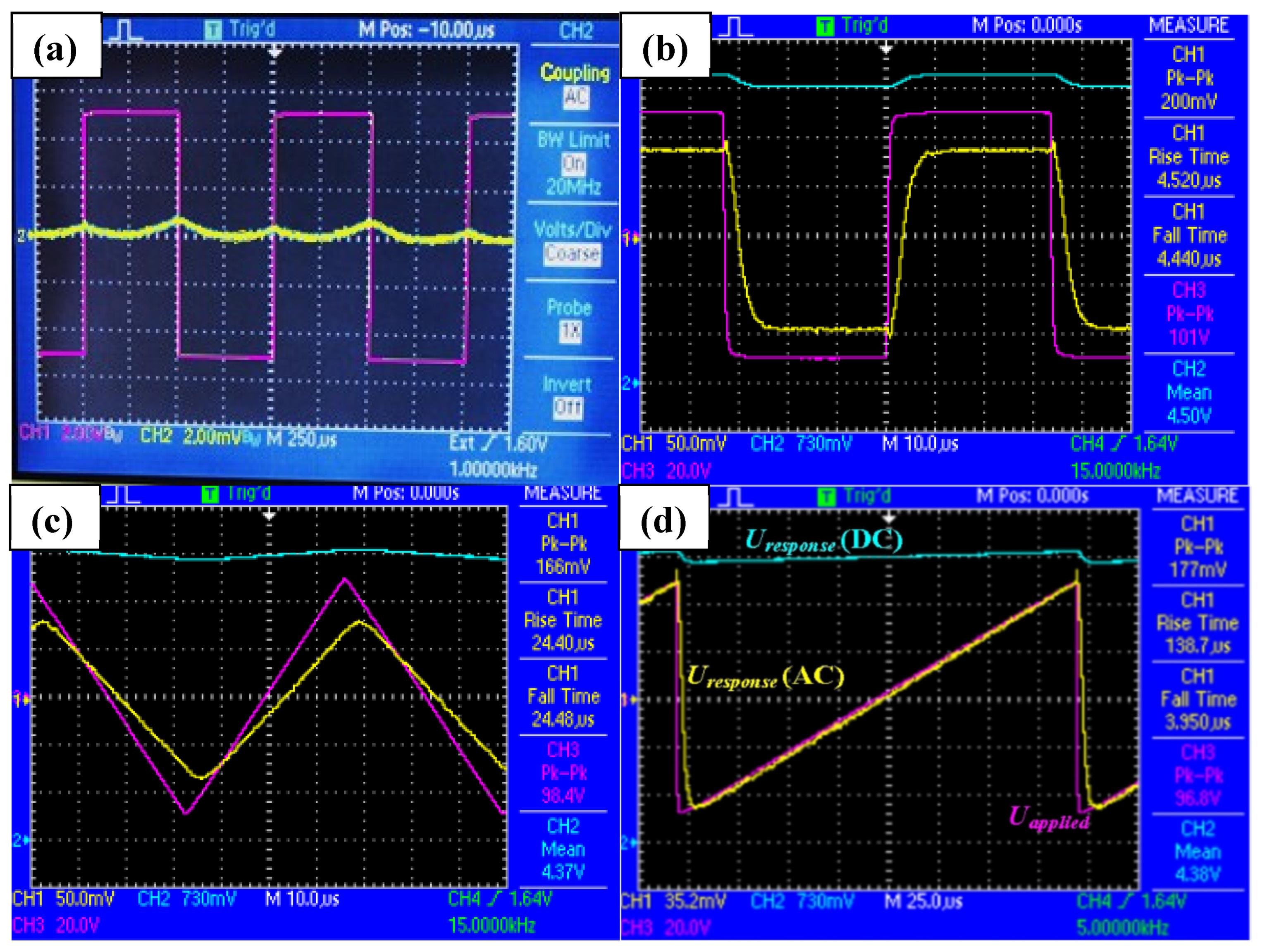
X phase transition and the sample history. In some cells, the domains of dimensions as large as 2 mm in length/breadth are observed. Repeated slow heating and cooling of the sample cell under electric fields of a single polarity could have played an important role in determining the outcome of this experiment. DC and AC coupled signals are fed to the separate channels of the oscilloscope, and the varying (switching) component of the signal relative to the input can be zoomed due to AC coupling. A close similarity of the applied and output waveforms (
Figure 3b–d) confirmed the linear nature of the switching.
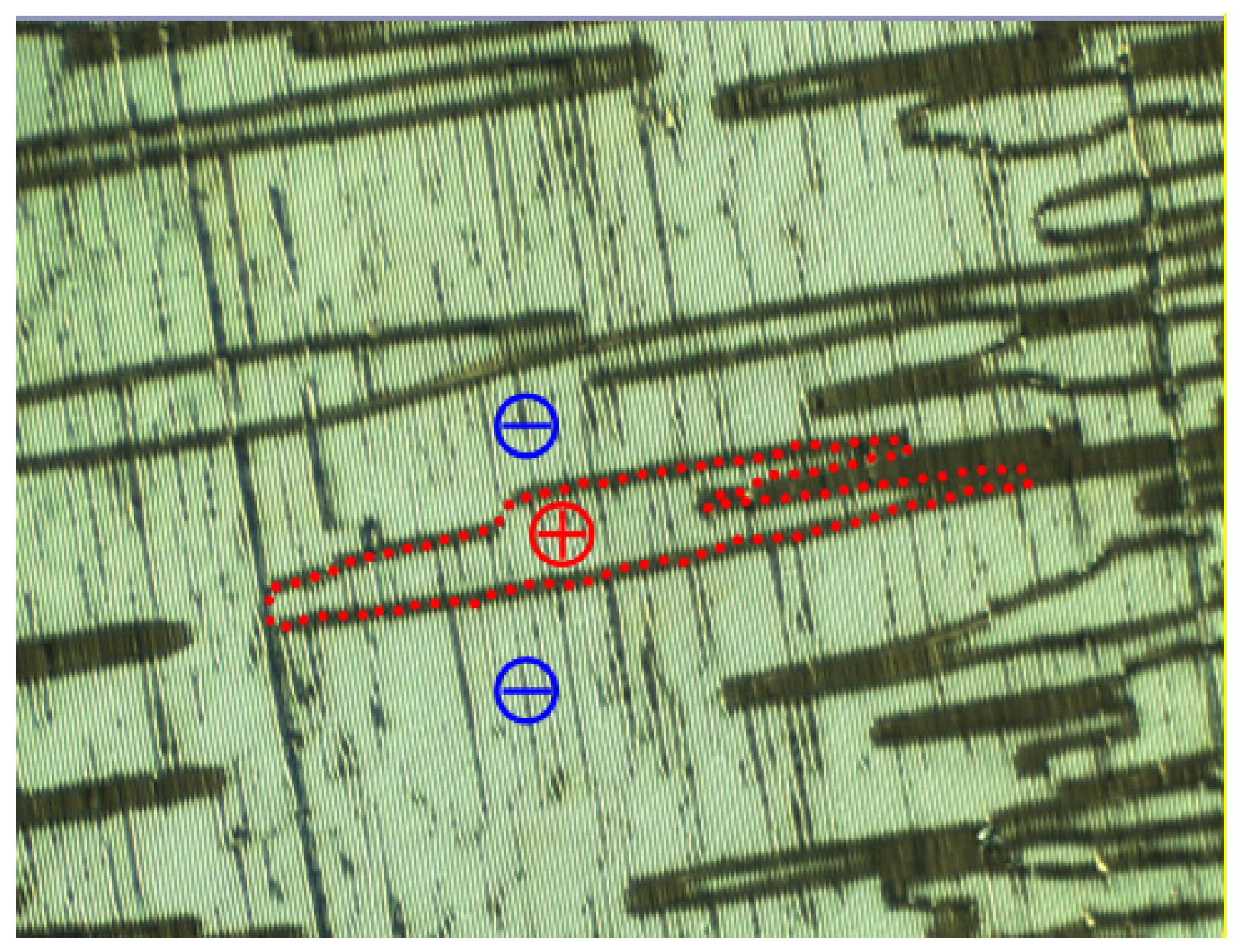
The results of the electro-optical experiment clearly confirm that the domain of a nematic phase in an LC cell under the field of view of a microscope is of single chirality [
16,
18]. Furthermore, the domain exhibits unusually fast electro-optic dynamics. To determine how such a chiral domain originated in the cell in the first instance, we examine the emergence of the domains observed under the polarising microscope in greater detail (
Figure 4). The domain boundaries are identified as ‘discontinuities’ in a spontaneous emergence of the stripe pattern [
15,
16,
17,
18]. The two neighbouring domains gave an opposite switching response to the electric field applied [
16,
17,
18]. The overall symmetry of the sample is consistent with the achiral nature of the material in a planar-aligned cell if the chiral domains of opposite chirality in a nematic phase also appear in the cell. The phase in question was proven to be nematic by X-ray scattering, as already been stated [
15,
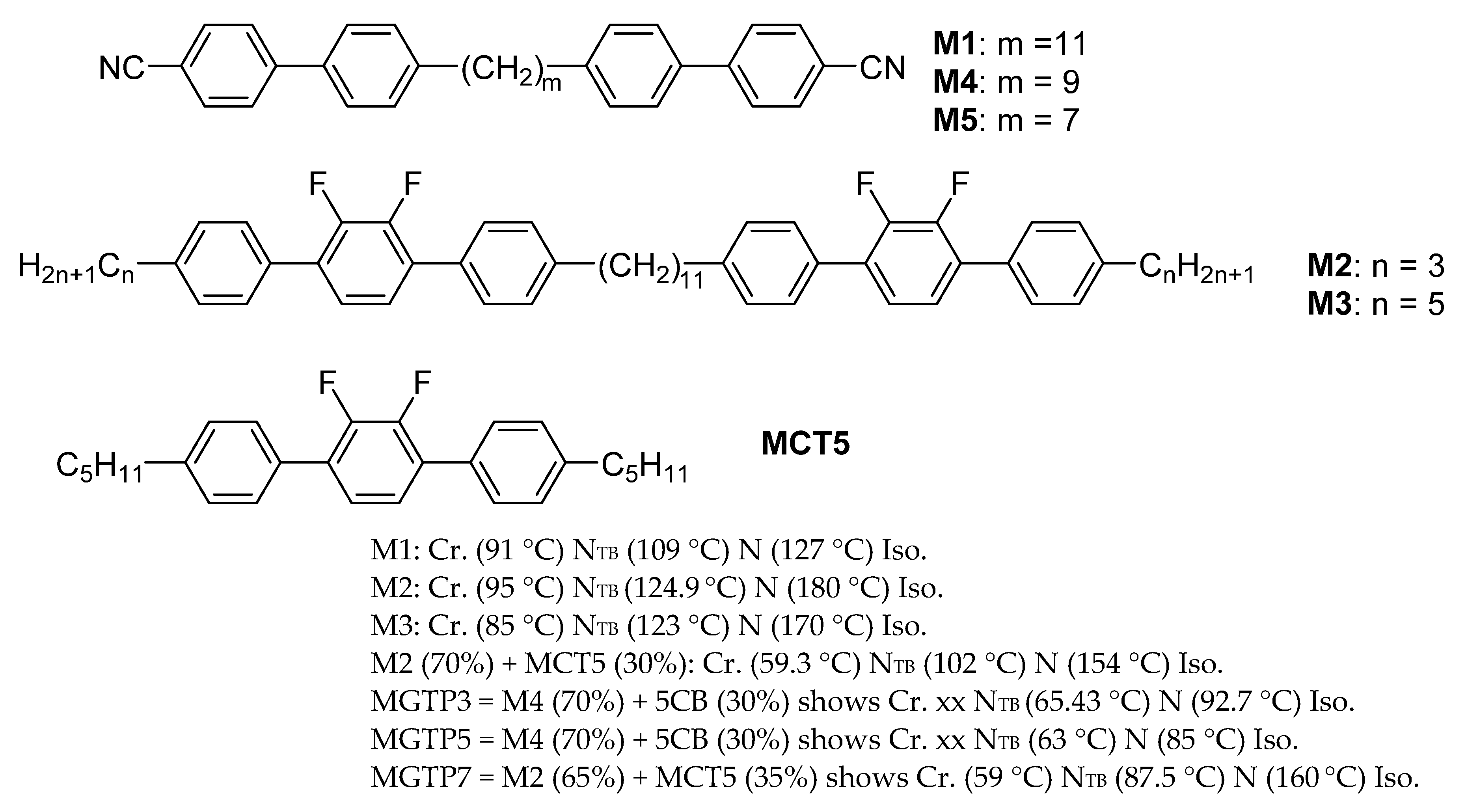
19]. Such observations were repeatable and reproducible for the compounds and the mixtures, composed of achiral molecules, investigated as listed in
Figure 2.
When an LC cell is rotated by a small angle from its dark position clockwise or anticlockwise, a variation in the transmitted intensity (from dark to bright or reverse) is observed in between the two neighbouring domains [
16]. Similar observations were made by Cestari et al. [
20] soon after; however, these authors reported that neighbouring stripes (and not the neighbouring domains) were of opposite chirality. In our experiments, the domains were of large enough dimensions that each contained a significantly large number of stripes of the same chirality. Moreover, we found that the application of sufficiently large electric fields at high frequencies led to the formation of periodic patterns of such domains [
17]. The domains of opposite chirality were also confirmed to exist using
2H nuclear magnetic resonance experiments later by Emsley et al. [
21]. They converted the domains of opposite chirality or seeded to ‘the domain of a single chirality’ by doping the bimesogen with a chiral solute (S)-1-phenylethanol.
In our experiments, the system was cooled/heated slowly (0.2 °C per minute) under a weak DC electric field. The polar switching, as explained above, was observed with a switching time of a few microseconds. This meant that a domain of ‘a single chirality’ existed stably over the field of view of a polarising microscope, without having the need to seed it with a chiral dopant. A repeated slow heating and cooling of the sample cell could have resulted in the biasing of all the molecules, thus acquiring the bent shape of a single configuration (R) or (S). This could have been a major step forward for forming a single chirality domain.
3.3. Identification of NX Phase with the NTB Phase
A procedure of obtaining the domain of a single chirality without needing to use a chiral dopant offers significant potential in fabricating devices using chiral materials derived from achiral molecules. This method dispenses with following the most expensive route of synthesising chiral materials directly. Dozov had predicted that the twist-bend structure of weakly bent molecules in the nematic phase could lead to a chiral structure [
22]. It now turns out that the formation and the stability of this structure are due to a minimisation of the excluded volume [
23]. This, together with other evidence, has enabled the identification of this phase as the ‘twist-bend nematic’, ‘N
TB’ [
2,
3,
4,
18,
19,
22]. Henceforth, we shall refer to the N
x phase as N
TB. Essentially, it is a chiral structure created by the symmetry breaking of achiral molecules. Molecules are bent and twisted to form the twist-bend structure. The phase, however, is still nematic.
It may be restated that Dozov’s work [
22] emphasised the emergence of spontaneous chirality in a nematic phase from the twist-bend structure formed by achiral molecules with a bend elastic constant K
33 decreasing to almost zero and going to a negative value [
24]. This results in a spontaneous bend distortion of the director. The twist provides stability with a higher packing efficiency and a minimisation of the excluded volume, as is conjectured by Goodby [
23]. Whether the structure formed is splay-bend or twist-bend depends on the relative ratios of the other two elastic constants. However, the splay-bend phase does not lead to chirality. Since the domain observed was chiral, the ‘splay-bend nematic phase’ in its ground state is ruled out. We may add that the optical stripes observed in parallel-rubbed planar-aligned cells are not related to the chirality. The twist-bend of the molecules leads to a heliconical structure on the nm scale. The helical pitch of 8 nm was determined initially by Freeze Fracture Transmission Electron Microscopy (FF-TEM) [
2,
3], and it has recently been confirmed using other techniques of soft resonant carbon, selenium and sulphur K-edge X-rays [
25,
26,
27]. The structure of the phase at the micrometre scale [
28] is far more complex and intriguing than the locally biaxial helical structure on the nanometre (nm) scale would have suggested. A complicated hierarchy of structures on several different length scales [
18,
28,
29] requires detailed investigations to be carried out using techniques that enable high resolution, a wide field of view and 3D imaging of the structure. As the electric field is applied across the parallel-rubbed planar-aligned cell, the N
TB phase is converted to a splay-bend for fields larger than a few V/μm depending on the Δε and optical biaxiality. This was observed for the first time in [
30] especially for a bent-core LC, and was theoretically predicted by Longa et al. [
31,
32] soon after. This has recently been verified by Meyer et al. [
33] through birefringence measurements, which suggest that molecular biaxiality plays a key role in not only stabilising the N
TB phase but also in producing a field-induced splay-bend phase. Merkel et al. show through polarised IR measurements that biaxiality order parameters increase significantly in the N
TB phase with a reduction in temperature [
34].
The measurement of the birefringence [
16,
18,
19] gives both the deviation angle
ϕ per unit electric field and the switching time
τ for a uniformly lying helix. The angle
ϕ by which the optic axis deviates from its initial position in the plane of the cell normal to
E is given by:
The switching time τ is given by:
Our experiments yield values of
ϕ ~ 0.1° to 1.0° for
E = 10 to 20 V/µm [
16,
18,
19]. In Equations (1) and (2),
e is the flexoelectric coefficient,
p is the pitch of the helix,
K is the effective elastic constant,
γ is the viscosity, and
θc is the heliconical angle. The parameters
e,
k and
τ can be measured [
16,
35], and
can be estimated. Based on the experimental results, we estimated at an earlier stage that
p << the wavelength of UV light [
16];
p << 100 nm [
19,
35]. Coles et al. [
36] used a set of Equations (1) and (2) for the cholesteric nematic phase, except they used
K =
K1 +
K3 instead of
K =
K1 +
K2 [
4] for the N
TB phase.
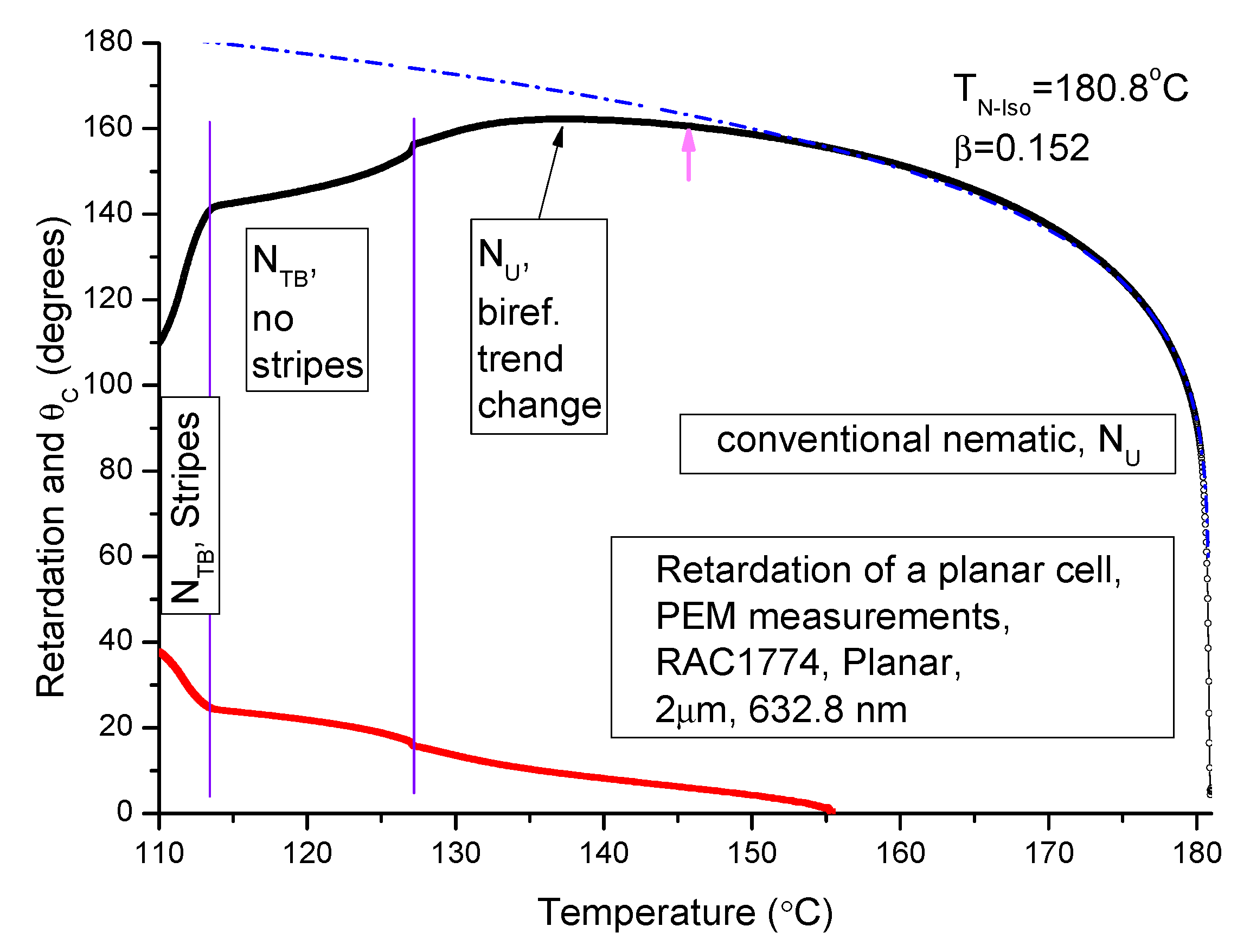
3.4. The Optical Retardation
The heliconical angle
θc can be determined from the optical retardation (OPR) or the phase difference between the output and input signals. The OPR is given by:
Here,
is birefringence,
d is the cell thickness, and
is the wavelength of the light beam for which the optical retardation is measured. The OPR (in degrees) measured by the PEM set-up designed by one of the authors (VPP) [
14] is plotted as a function of temperature in
Figure 5. We used a planar-aligned cell of cell spacing 2 μm. The material used in this experiment is fluorinated bimesogen denoted as M2.
Interestingly, a change in the trend of the ‘dependence of OPR on temperature’ within the high temperature nematic phase but closer to the N–NTB phase transition temperature is observed. The decrement in OPR starts at a temperature approximately 10 °C just above the NTB-N phase transition temperature using the procedure given below. In a similar manner, we estimate θc = 24.3° at 113.9 °C (in the NTB at a temperature just above the onset of the striped pattern). However, caution in the interpretation of the results needs to be exercised for two reasons:
An extrapolation of the OPR over a wide temperature range depends significantly on the temperature range selected in the power law fitting formula of Haller [
37]. The starting point of the temperature range is marked by an arrow (in colour magenta) in
Figure 5, and;
An extremely complicated structure of the N
TB phase may invalidate the use of the simple Haller formula [
37].
The birefringence
in the N
TB phase is given in terms of the heliconical angle
θc:
Here,
is the measured birefringence value at a temperature of interest. Δ
nextr is the extrapolated value at this temperature provided the transformation from N to N
TB had not occurred. Δ
nextr is obtained from Haller’s formula [
37]:
where the parameters Δn
o, T* and
β are obtained by fitting the experimental birefringence data acquired in the high temperature conventional nematic phase. Here, Δ
n0 corresponds to the birefringence value at the absolute zero temperature,
T* is the temperature of the Iso-N phase transition and
β is a critical exponent. On having determined these parameters, one can obtain Δ
nextr for temperatures corresponding to the N
TB phase using Equation (5) and then calculate
θc using Equation (4).
We find that
θc, calculated from the results of the OPR data, starts rising in the high temperature N phase (see
Figure 5 red line). The heliconical angle is ~ 10° at a temperature of 14 °C above the N–N
TB transition temperature. At first, this angle increases linearly with a reduction in temperature, followed by a jump at the N–N
TB transition temperature. The observation is reminiscent of the first order phase transition. The important finding here is that the ‘heliconical angle’ emerges in the high temperature N phase itself, and it continues to increase with a decrease in temperature. The result is consistent with that obtained from NMR [
38] and by dielectric spectroscopy [
30]. A change in the nature of the high temperature N phase above the N–N
TB transition temperature may be due to the pretransition effect caused by the first order N–N
TB transition [
19].
3.5. The Fréedericksz Transition in the NTB Phase
In this section, we examine the electro-optical response in the N
TB phase following the application of an electric field across a planar-aligned cell, filled with achiral bimesogens having Δ
ε > 0. The response is found to be of two types: the (a) linear and (b) non-linear. The latter is due to the Fréedericksz transition, of a type shown here. This is different in the N
TB that in the N phase. The linear response has already been discussed in greater detail in
Section 3.2.
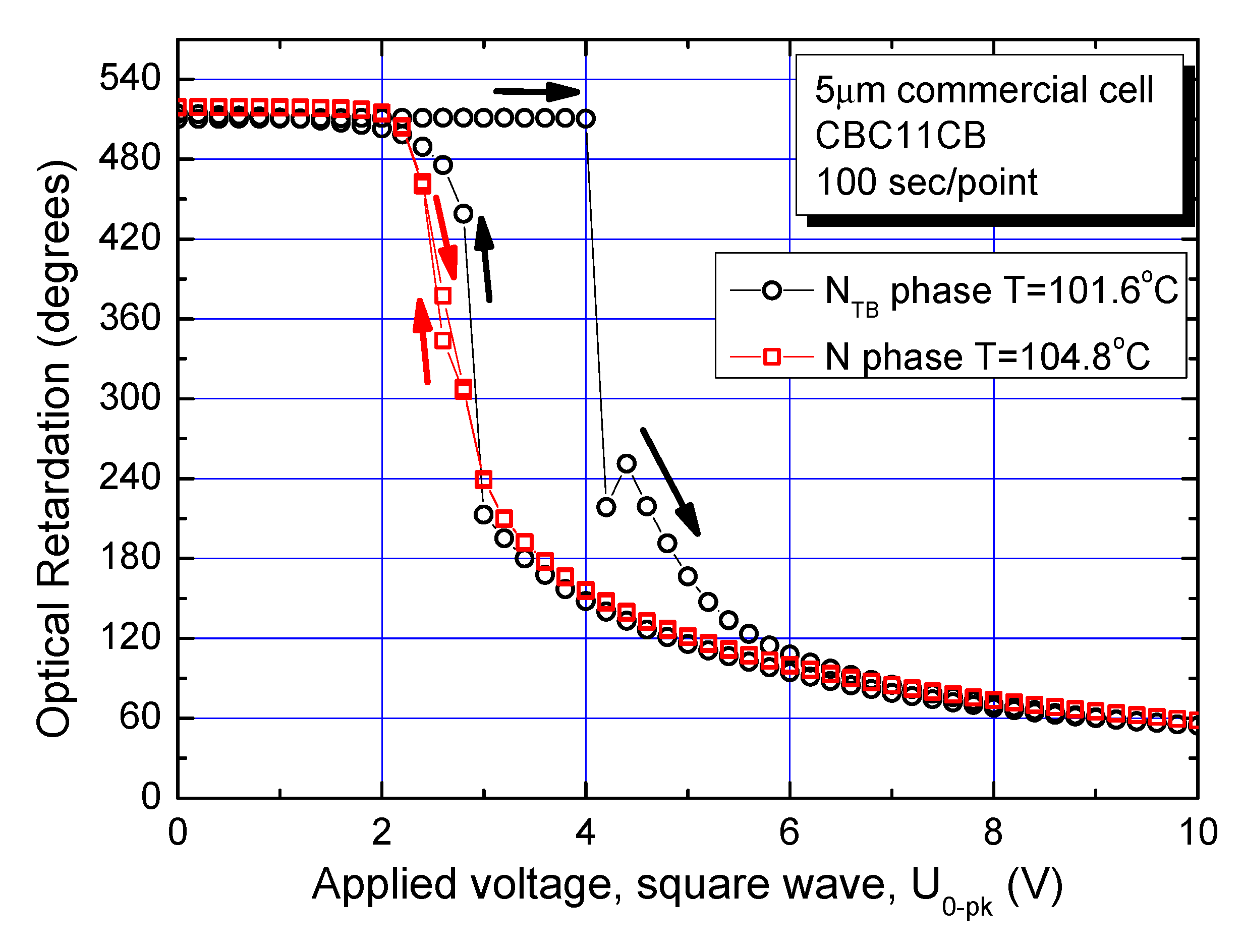
In terms of the differences in the characteristics of the two phases, the Fréedericksz transition is investigated for both N and N
TB phases of a planar-aligned cell. The optical retardation (OPR) is measured using a photo-elastic modulator (PEM) system [
14] following an application of square waveform across a commercial parallel-rubbed planar-aligned cell of sample thickness 5 μm. The cell is filled with the cyanobiphenyl bimesogen CBC11CB (M1). The dielectric anisotropy for both N and N
TB phases is finite and positive. The amplitude of a square waveform is first increased, and then decreased gradually. Each measurement point is fixed to have 100 s delay time inserted in the computer program of an automated measurement procedure to eliminate the contribution to the output from an increased viscosity of the system. Hence, the experiment takes a time span of approximately 5 h to complete. The samples in the LC cell are chemically stable over a longer period. The experimental data, recorded just ~0.5 °C below the N–N
TB phase transition, are plotted in
Figure 6 as a function of applied voltage. For temperatures much below the N–N
TB phase transition temperature, a manifold increase in the viscosity of the material makes this experiment almost impossible to conduct.
An interaction of the electric field applied across the parallel-rubbed planar-aligned cell with a material of positive dielectric anisotropy makes the constituent molecules start turning from the horizontal aligned positions to the direction vertical to the cell-windows. Consequently, the optical retardation decreases with an increase in the field strength. Furthermore, the Fréedericksz transition is mostly of the second order in the N phase. It is rarely observed as a first order transition except under the bias field [
39]. No hysteresis is observed in the optical retardation values in between the two cases of the field increasing and the field decreasing in the N phase, as the response data for both waveforms coincide within the experimental error. The data for the OPR in the N phase are shown by red squares in
Figure 6.
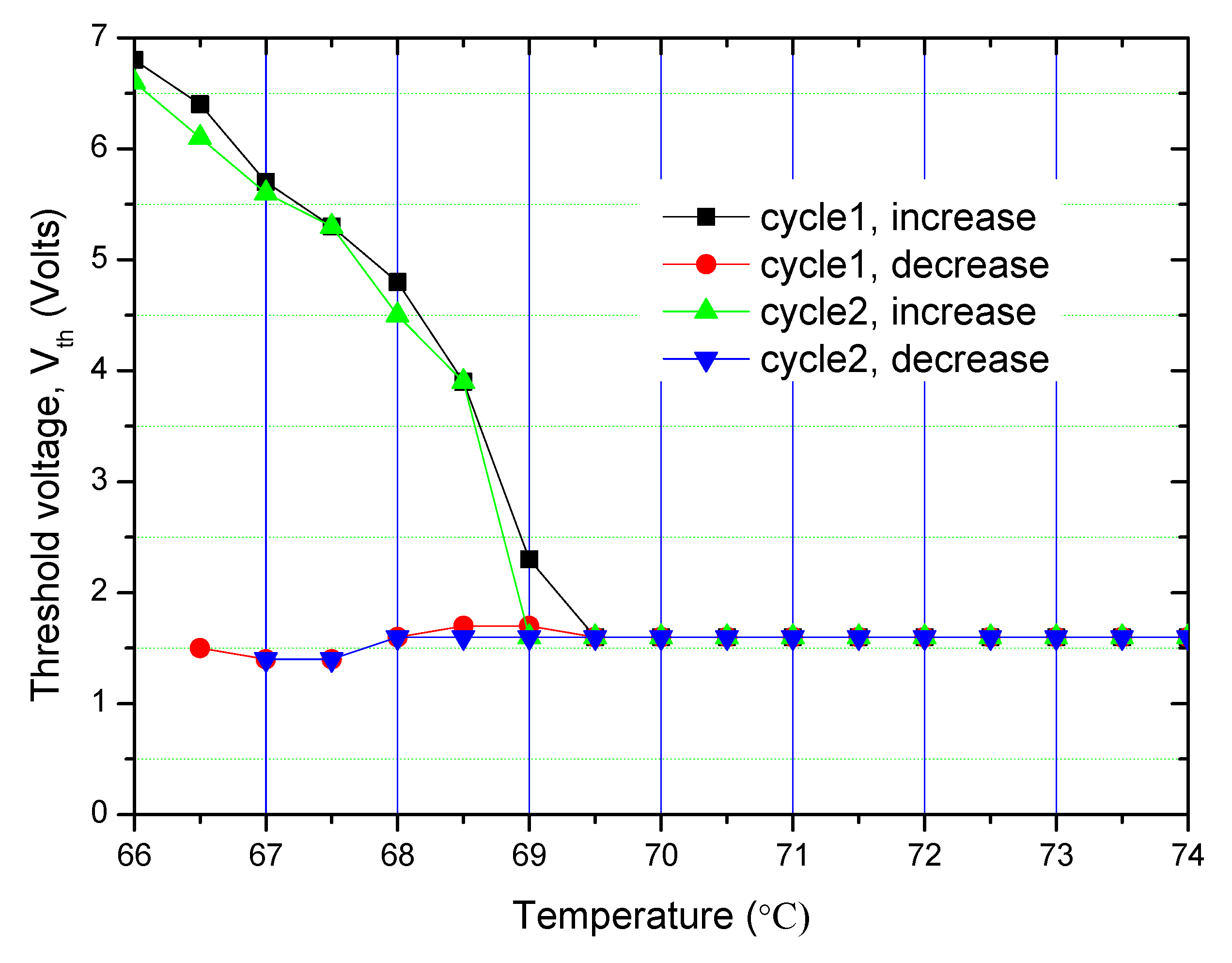
A good reproducibility in the results of the experiment is observed for repeated recycling of the input signal. The results are shown in
Figure 7.
The threshold voltage, V
th, is defined as the voltage for which OPR suddenly drops to a lower value from its initial value following the application of an electric field. A significantly large hysteresis in the threshold voltage is observed in the N
TB in contrast to almost none in the N phase at higher temperatures (
Figure 7). The Fréedericksz transition is observed as the first order in the N
TB phase as opposed to the second order in the N phase. Though a large increase in viscosity with a reduction in temperature in the N
TB phase could have played a significant role in leading to a larger hysteresis, its effect is eliminated here by conducting measurements with a long time delay inserted in between acquisitions of the successive data points. We suggest that hysteresis in the N
TB phase is due to its polar nature. The magnitude of hysteresis is relatively small when compared to the one arising from ‘bistability’ of the electric polarisation in ferroelectric liquid crystals.
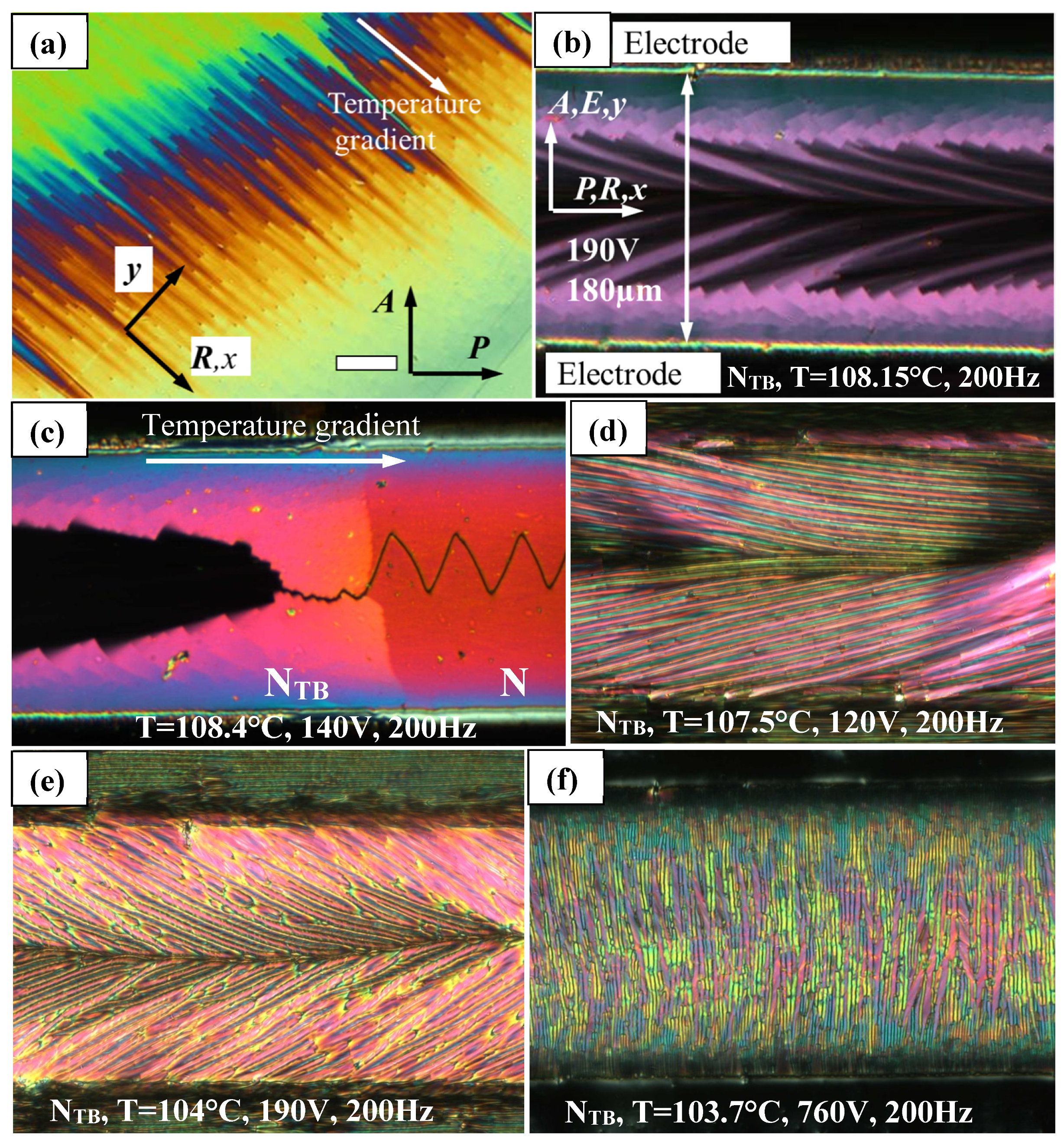
Data presented in
Figure 6 and
Figure 7 can be illustrated by POM image in
Figure 8a showing the phase transition in planar parallelrubbed sandwich cell. Due to a small temperature gradient and temperature dependence of the threshold voltage, one can observe step-wise change of color along the temperature gradient direction.
Fascinating images can be obtained using in-plane electrodes in planar rubbed cell (
Figure 8b–f) by applying a variety of voltages in the vicinity of the N–N
TB phase transition. Once again, sharp change is visible between switched (bright color) and non-switched states (dull color) demonstrate the first order of the transition (
Figure 8b,c). Difference between the phases is clearly visible due to a small temperature gradient present in
Figure 8c. When cooling the sample with a voltage applied, the self-deformation striped pattern appears with the stripes extending at an angle to the original rubbing direction (
Figure 8d–f). Since for this material dielectric anisotropy is positive, this is one more confirmation that the stripes develop along the average director. Finally, for large fields,
Figure 8f, the stripes are observed parallel to the electric field.
Based on the results of the experiment shown in
Figure 8 and
Figure 9, we conclude that a homeotropic aligned-cell containing materials/mixtures of negative Δε listed in
Figure 2, with the AC field, applied across the cell shows a first order Fréedericksz reorientation in the N
TB phase. In contrast, a classical N phase normally exhibits a second order transition [
3], except when a large DC bias field is applied across the cell [
39]. Here, the optical axis is realigned gradually and uniformly across the sample as the field exceeds the threshold. In the N
TB phase, a reorientation of the optic axis seems to start only at isolated sites, and each site is presumably associated with the dust particle/s or a surface irregularity. The nucleating regions in the shape of asymmetric focal conic domains [
40] coexist with the unswitched homeotropic state. Furthermore, the threshold voltage exhibits a significant hysteresis (
Figure 9 has almost 3V difference between the left and the right columns). According to the data obtained for the same homeotropic geometry using PolScope [
3], the deformations of the optical axis are of splay and saddle-splay type in the N
TB phase.
3.6. Stripes in the NTB Phase
- (i)
Planar-aligned cell
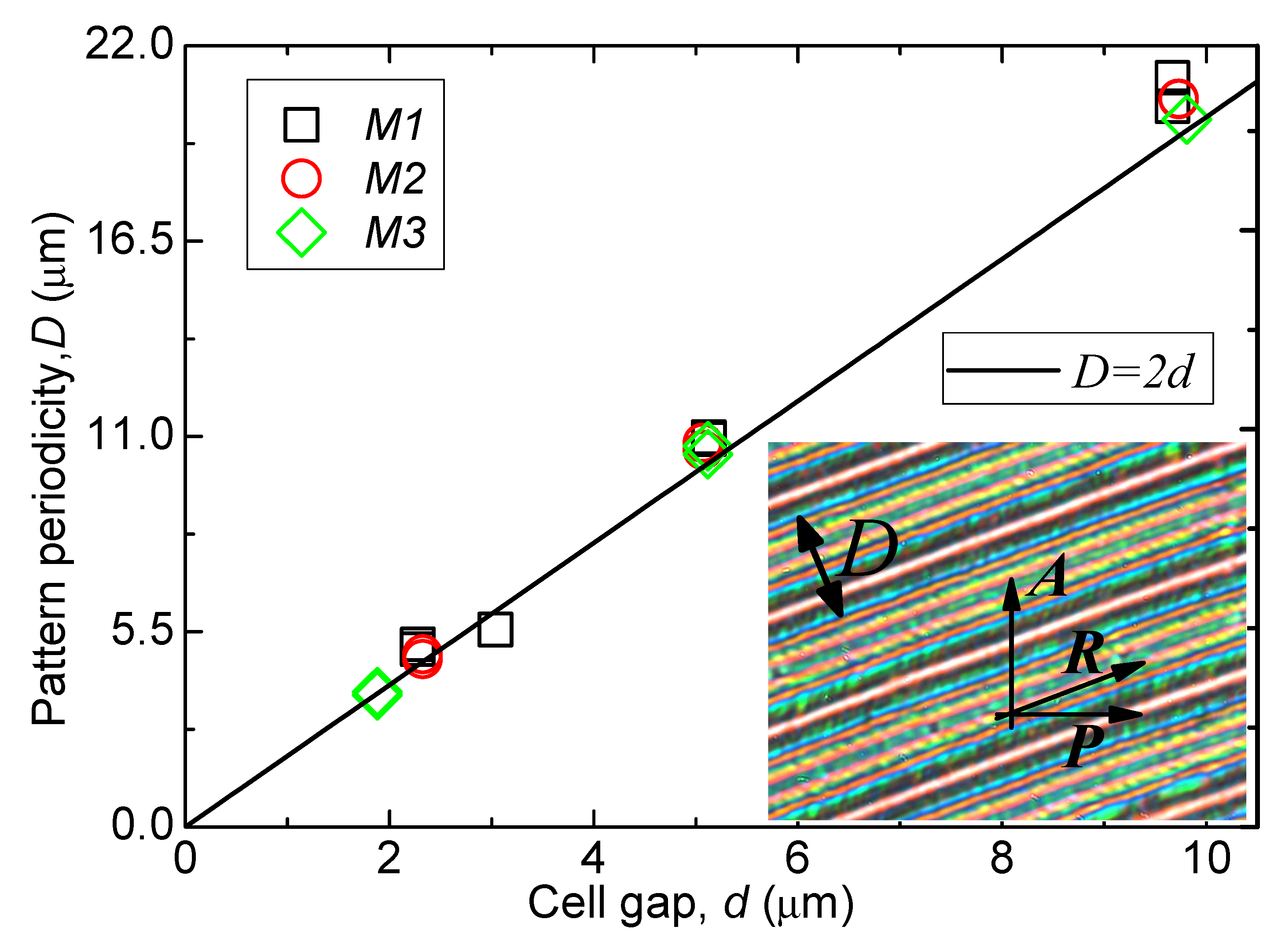
Beautiful stripes as observed in a thin planar-aligned cell under crossed-polarisers are shown in
Figure 10 [
15]. To explore these stripes further, the cell thickness in the experiment is varied from 2 μm to 10 μm. In the N
TB phase, three materials (M1, M2, M3) exhibit uniform stripes parallel to the rubbing direction in each case. For each cell thickness, the width of a stripe or distance between the successive stripes, called ‘the stripe periodicity’, is 2 times the cell thickness. Periodicity is dependent only on the cell thickness, but is independent of temperature. Three bimesogen/dimer systems investigated here showed similar behaviour. The phenomenon is material-independent so long as it satisfies the criterion of exhibiting a twist-bend nematic phase. At much lower temperatures within the N
TB, the stripe texture is too complex to be understood. For cell thicknesses greater than 12 µm, focal conic textures are observed. These are reminiscent of the typical texture of a smectic phase, discussed later. Nevertheless, an absence of smectic layers was confirmed by X-ray experiments [
15] in this phase. For this reason, it was initially labelled as Nx (identified later as N
TB). We refer to the virtual layers in the N
TB as the pseudo-layers.
The stripes arise presumably as a direct consequence of mechanical Helfrich–Hurault instability due to an increase in the heliconical angle with a reduction in temperature, as discussed in
Section 3.4,
Figure 5 (red squares). This leads to a correlated decrease in the pseudo-layer thickness. These domains may have also arisen from an interaction of the flexoelectricity with the electric field produced by the surface polarisation. The pyroelectric data do show a rather strong surface polarisation signal both in the twisted and hybrid cell (planar alignment on one electrode and homeotropic on the second), even in the isotropic phase of the chemical systems investigated [
36].
- (ii)
Twisted liquid crystal cell
Not only do the planar-aligned cells exhibit beautiful stripes in the N
TB phase discussed above, the twisted planar-aligned cells exhibit even more intriguing behaviour. The rubbing direction on the top electrode is at right angles to that on the bottom electrode. A delicate balance among the following variables: surfaces, material properties and the sample history, leads to display of a wide variety of patterns. Intriguingly, in some cases it is difficult to investigate patterns on account of the large sensitivity of their formation in experimental conditions. The material used in the LC cell is MGTP7, the composition of which is given in
Figure 2, and it is characterised by a large negative Δε. The stripe periodicity is temperature dependent in a twisted cell in contrast to the temperature independence in a parallel-rubbed planar-aligned cell.
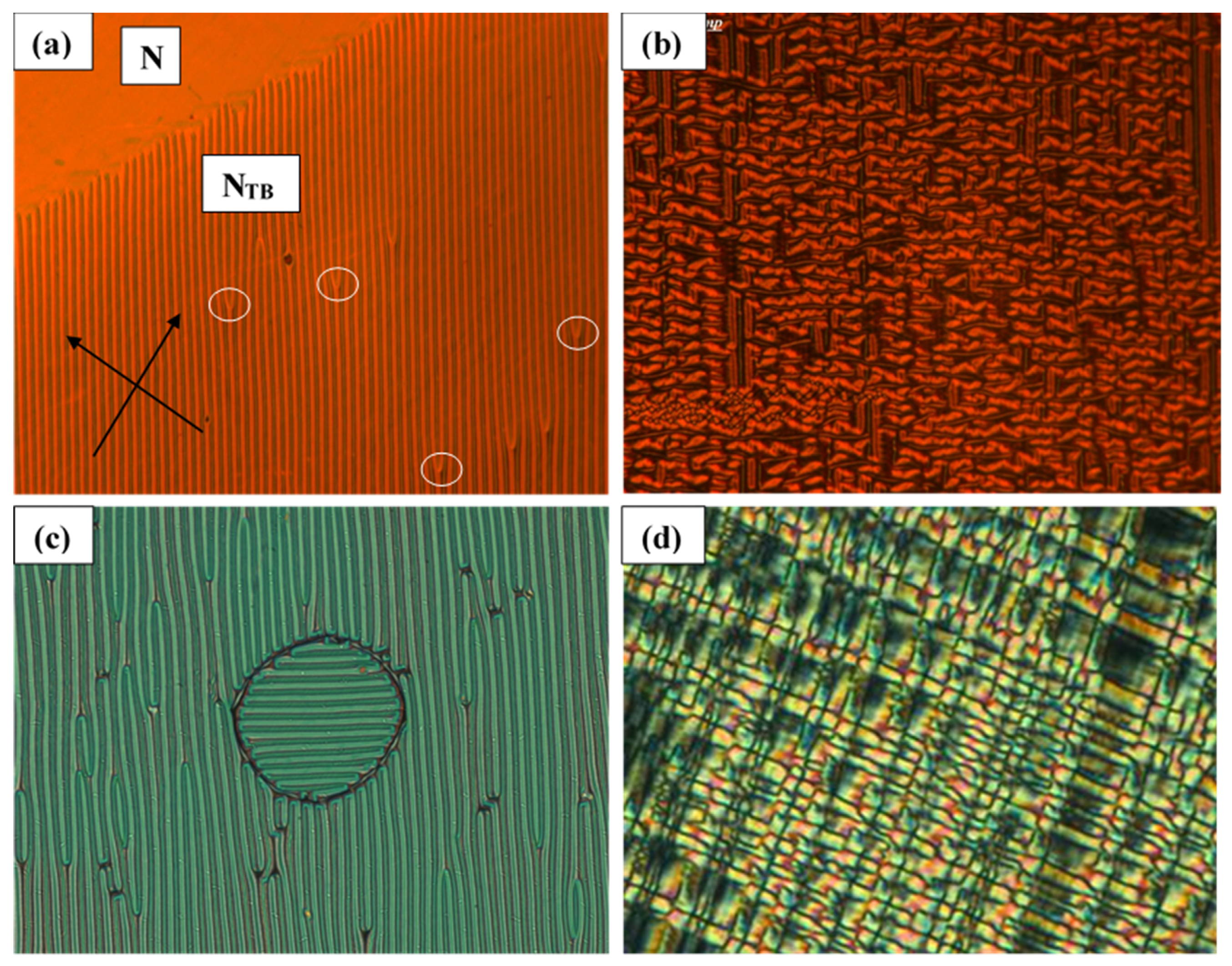
Due to the presence of a small temperature gradient in the cell, the top part of
Figure 11a is at the highest temperature and is in the N phase. Some stripes discontinue (fork defects are created at places marked by white circles), this provides a space for the increase in the periodicity of the pattern at lower temperatures. The plot demonstrating detailed temperature dependence of the stripe periodicity in twisted cells is shown in
Figure 12.
The texture in
Figure 11b is recorded at a lower temperature of 4 °C than the transition temperature. The pattern formed is rather complex and only some stripes manage to survive. Two domains at right angles to each other are observed in a twisted cell (
Figure 11c). An alternative pattern of ‘crosses’ is observed after an electric field is applied to a material with positive Δε planar-aligned in a twisted cell of thickness 8 μm. The temperature here is 5 °C below the N–N
TB phase transition temperature.
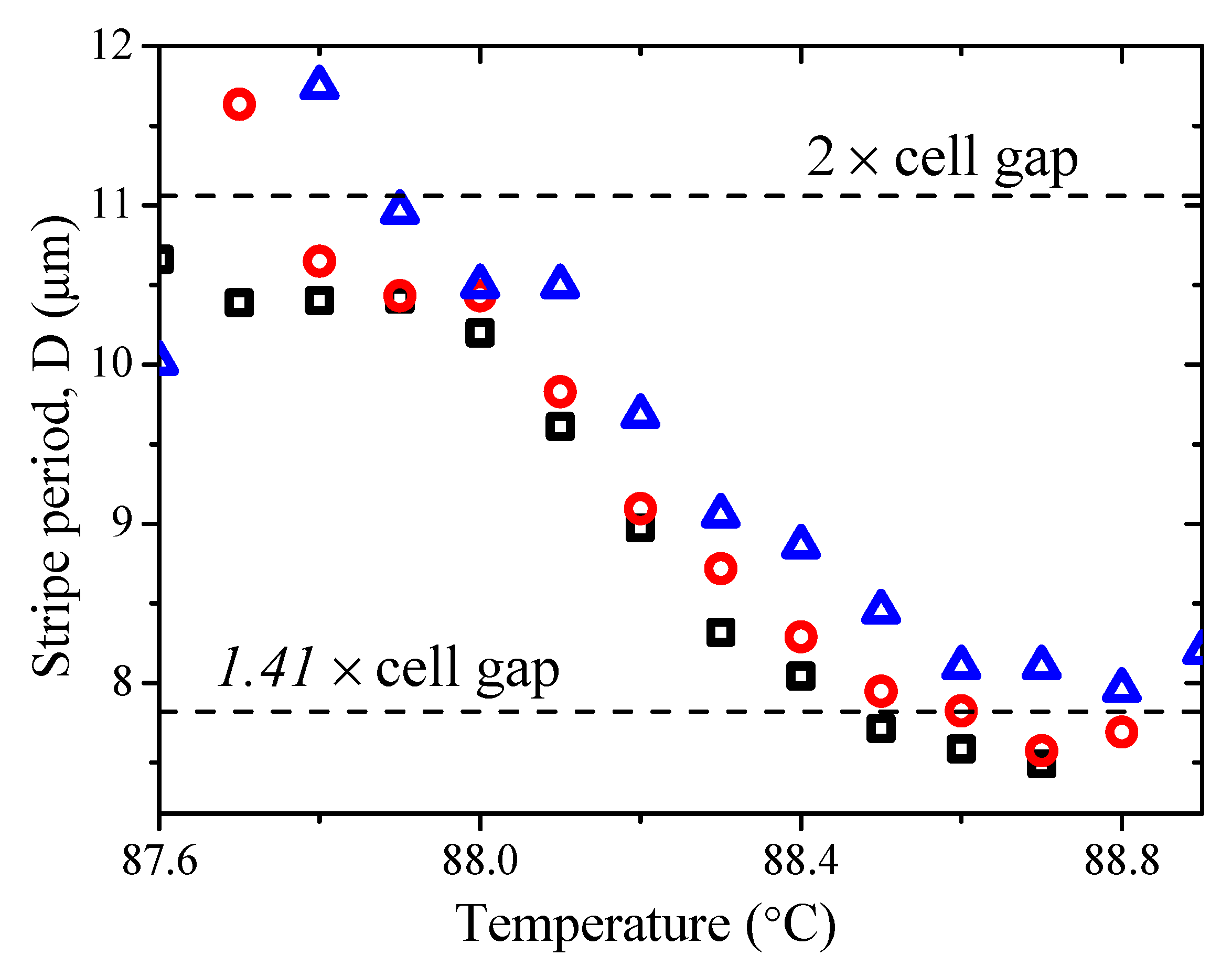
The stripe periodicity as a function of temperature in a twisted cell is shown in
Figure 12. In a twisted cell, we note that the stripe periodicity
D is temperature dependent, and it decreases with a reduction in temperature in the N
TB. It would seem that twist distortions in the deformations of molecules ‘enable’ the formation of stripes with a periodicity different from the double cell gap. Here, the stripe periodicity varies between 1.4 and 2 times the cell gap,
d. This behaviour is different from cells with parallel rubbing, where the liquid crystalline structure at micrometer hierarchy level demonstrates “binary” behaviour, either staying uniformly aligned, or forming double-cell-gap periodic pattern.
When we apply a square wave of frequency 10 Hz and amplitude 2 V at a temperature of 87.5 °C (just 0.5 °C) below the N–N
TB transition temperature, some of the stripes start disappearing. The applied electric field seemingly suppresses the undulations of the pseudo-layers, see
Figure 13. The undulations are suggested to be the cause of the emergence of stripes. The pseudo-layers lead to kinds of Chevron structures, where the molecules are bent in some cases.
3.7. Templating the Polymerised Structure of a Liquid Crystalline Phase
The N
TB phase persists over a wide range of concentrations of the bimesogen mixed with non-N
TB materials [
28]. The latter includes the monomer and a reactive mesogen RM257 photo-initiator. The planar-aligned cell filled with the mixture shows a variety of textures depending on the temperature and the phase. The reactive mesogen is also a liquid crystalline material. The molecules of the reactive mesogen fill up the voids and thus these follow the structure of the phase. In the phase of interest, the LC mixture is polymerised by shining UV light onto the sample through an optical microscope. A sponge-like polymer template of the structure of the ‘phase of interest’ is thus formed in the cell. Unpolymerised liquid crystalline material is washed away with acetone. The polymerised sponge has the structure of the phase prior to having the polymerisation initiated. The polymer so formed is imaged using SEM.
For SEM imaging, direct access to the surface of the sample is needed. Two sample preparation methods are used:
Open cell: An appropriate mixture prepared as above is dissolved in acetone. The liquid so formed is put onto a glass substrate which had previously been sputtered by indium tin oxide (ITO), and is spin-coated by the alignment layer. As the solvent evaporates, a film of several µm in thickness is formed on the substrate. It is then brought to a temperature of the N
TB phase by heating. The film in the phase of interest is polymerised and afterwards scanned by SEM. The focal conic domains as observed are shown in
Figure 14a. For examining the structural details, the image is scanned with a higher magnification of SEM, as shown in
Figure 14b. Focal conic domains observed by POM in cells of thickness greater than 10 µm in colour are given in the inset. This method suffers from drawbacks such as the lack of control of the sample’s thickness, the effect of surface tension of the liquid on the shape of the film and the need to negate the effects of oxygen on polymerisation. One needs to perform experiments in an O
2-free atmosphere or, alternatively, significantly increase the concentration of the photo-initiator.
Sandwiched cell: In this case, a standard commercial sandwich cell is used. The cell is filled with the mixture and is polymerised in the phase/texture of interest. Though we avoid some of the drawbacks of the first method, nevertheless, this technique may damage the structure of the phase in the sponge itself while the two substrates are pulled mechanically apart. However, we do expect that the polymer template retains the structure of the phase of interest in the middle of the cell. The structure corresponding to the N
TB so formed is imaged by SEM, and images are shown in
Figure 14c,d, the former scanned with a higher magnification of SEM than the latter.
A highly conductive sample is required for the SEM imaging. In most cases, a thin metal film needs to be deposited onto a polymer. However, the electron microscope (Zeiss ULTRAplus®/Geminy®) can satisfactorily image the samples without having to coat it with a metal, provided (i) the ITO coating is present on the glass substrate and (ii) the thickness of the polymer sponge is below 2 µm.
The results of the imaging of the structures are extremely interesting. The fish-bone structure shown in
Figure 14d can explain the ‘saw-tooth’ pattern given in
Figure 8b. The average director of the structure deviates from the rubbing direction in the cell by the applied electric field by a certain angle. Hence, the two sides of fish-bone texture exhibit dramatically different intensity levels between the crossed polarisers since on one side the ‘fish-bone’ angle α is subtracted from this angle, while on the other side it is added to it. Therefore, on one side of the fish-bone structure, the optical axis is parallel to the polariser/analyser and it appears dark, while on the other, it deviates by a significantly large angle and thus it appears bright.
Moreover, an analysis of the SEM images shown in
Figure 14c,d and reported in [
28] has led us to conclude that the ‘self-deformation stripes’ exhibit ‘fish-bone’-like structures at the micrometre level. Meanwhile, at the nanoscale level of the hierarchy, the phase is helical. Hence, it is an interesting challenge to determine the relationship between the helical axis and the direction of the ‘fish-bone’ structure. Since the fish-bone angle α approaches 45° at a lower temperature in the N
TB phase, and it has a lower value closer to the N–N
TB transition temperature, one can expect it to cross-over 22.5° at some temperature. In this case, the sign of the fast electro-optic response of fundamental frequency discussed in
Section 3.2 and in [
16] should change its sign, as is explained (supplementary material of Ref. [
28]). Such a change in sign in the electro-optic response is observed in the terphenyl bimesogens M2 and M3 [
28]. Therefore, we conclude that the helical axis of the nanoscale pitch is likely to lie along the ‘fish-bone’ observed on the micrometre scale.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}













