

Tryptophan, an Amino-Acid Endowed with Unique Properties and Its Many Roles in Membrane Proteins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Trp in Membrane Proteins
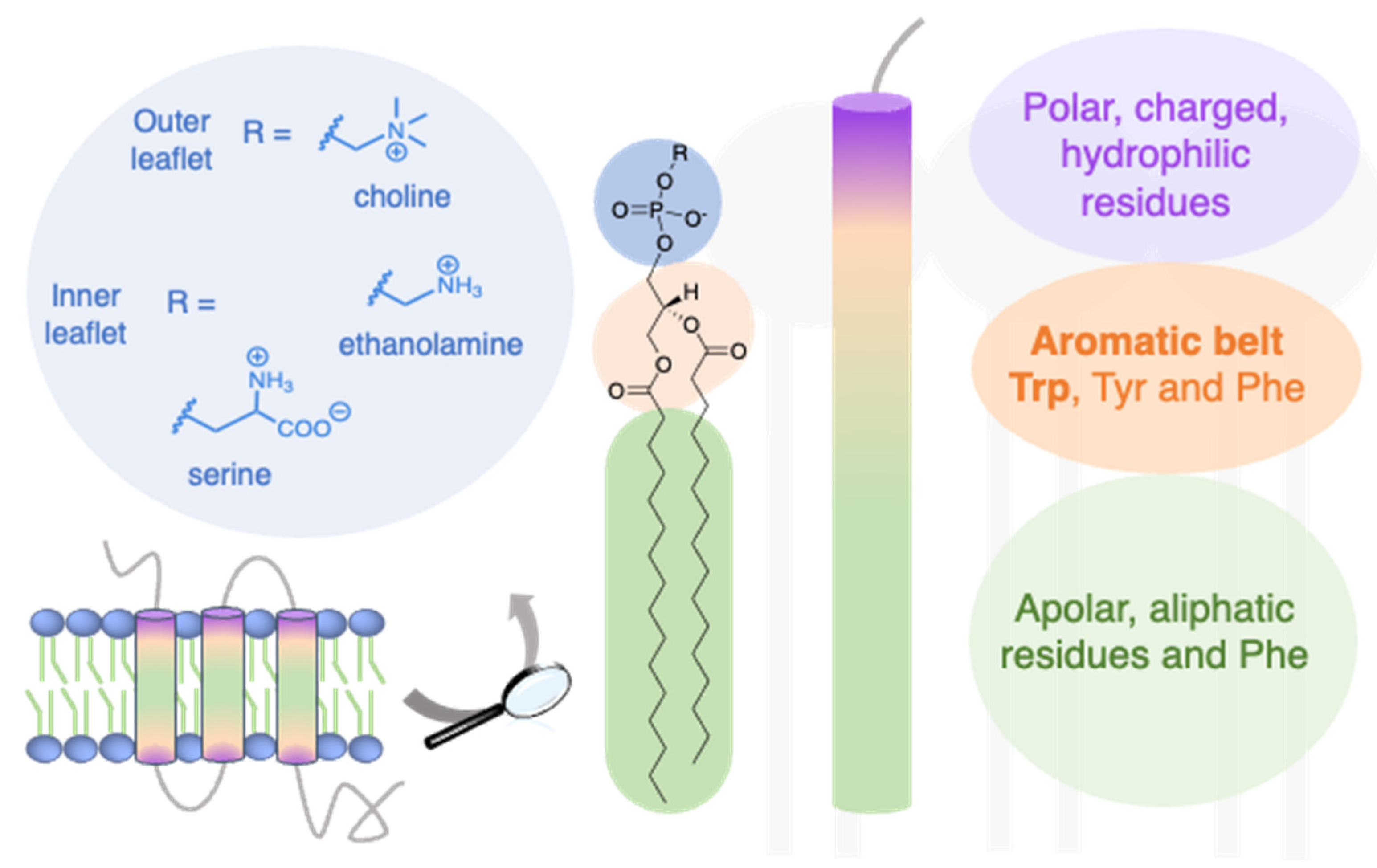
2.1. Trp Localization at the Lipid/Water Interface of Membrane Proteins
2.2. Trp in the Stabilization of Membrane Proteins
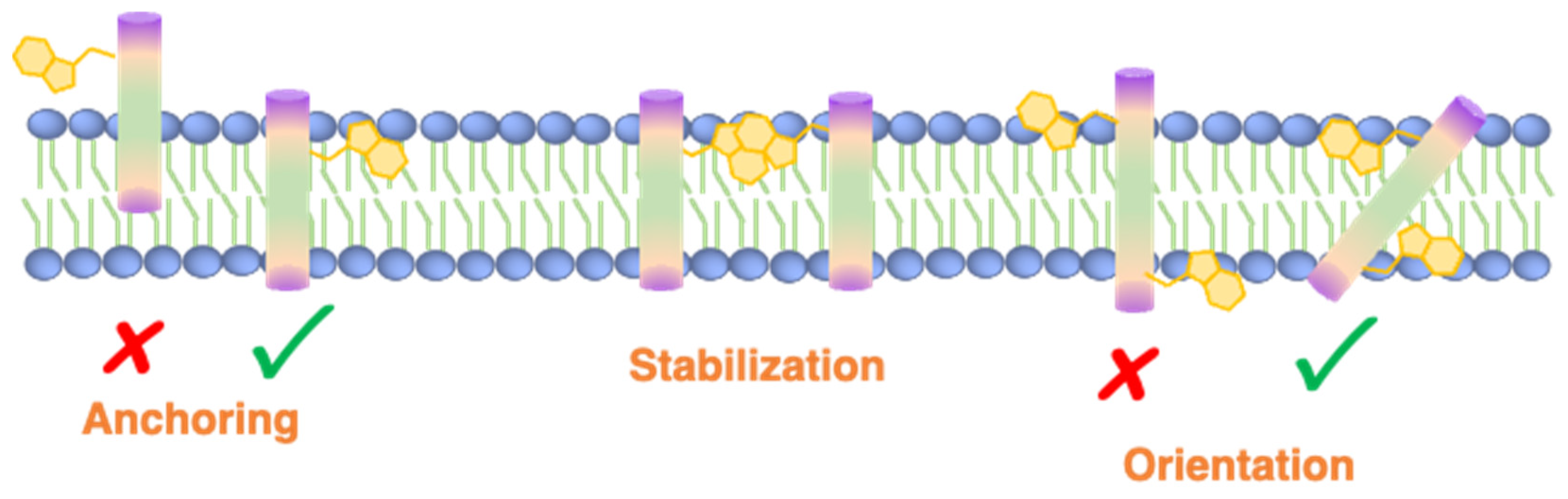
2.3. The Role of Trp in Anchoring and Orientation of Membrane Proteins
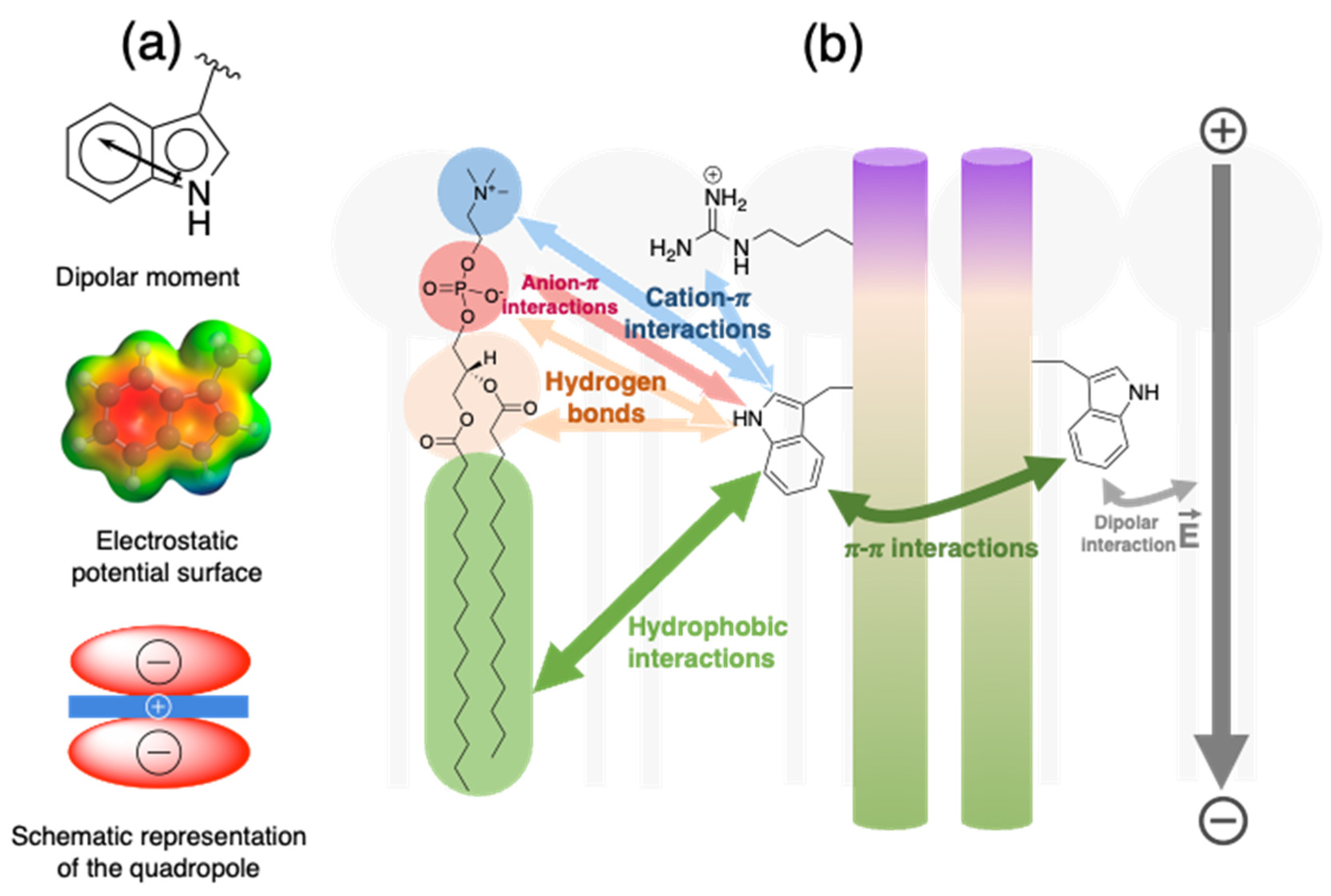
3. Noncovalent Interactions Engaging Trp
3.1. Trp Interactions with Lipids
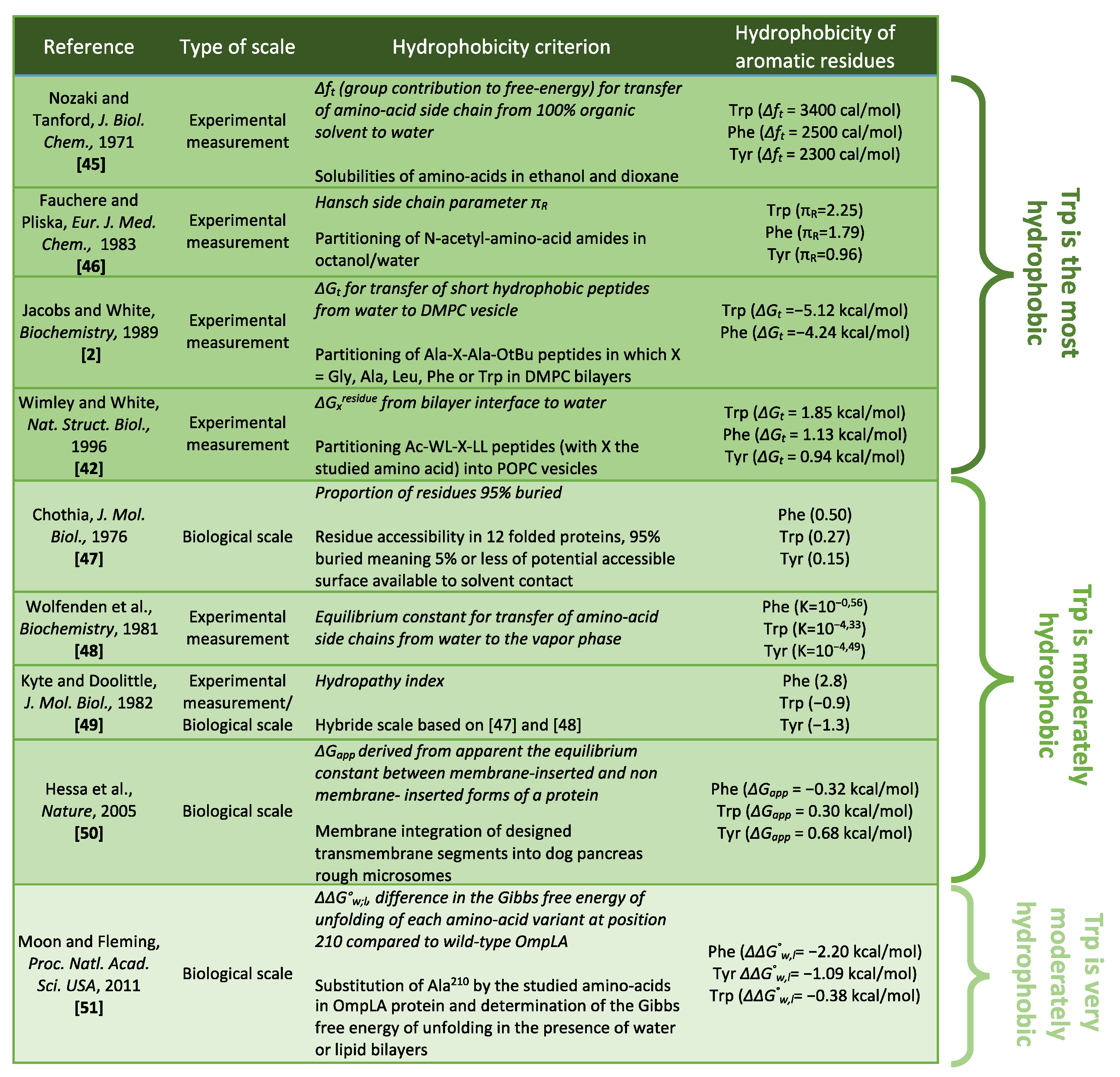
3.2. Hydrophobicity and Hydrophobic Effects
3.3. Electrostatic Interactions
3.4. Hydrogen Bonds
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- von Heijne, G. The distribution of positively charged residues in bacterial inner membrane proteins correlates with the trans-membrane topology. EMBO J. 1986, 5, 3021–3027. [Google Scholar] [CrossRef]
- Jacobs, R.E.; White, S.H. The nature of the hydrophobic binding of small peptides at the bilayer interface: Implications for the insertion of transbilayer helices. Biochemistry 1989, 28, 3421–3437. [Google Scholar] [CrossRef] [PubMed]
- Wallin, E.; Tsukihara, T.; Yoshikawa, S.; Von Heijne, G.; Elofsson, A. Architecture of helix bundle membrane proteins: An analysis of cytochrome c oxidase from bovine mitochondria. Protein Sci. 1997, 6, 808–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Planque, M.R.R.; Kruijtzer, J.A.W.; Liskamp, R.M.J.; Marsh, D.; Greathouse, D.V.; Koeppe, R.E.; De Kruijff, B.; Killian, J.A. Different membrane anchoring positions of tryptophan and lysine in synthetic transmembrane α-helical peptides. J. Biol. Chem. 1999, 274, 20839–20846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, R.E.; White, S.H. Mixtures of a series of homologous hydrophobic peptides with lipid bilayers: A simple model system for examining the protein-lipid interface. Biochemistry 1986, 25, 2605–2612. [Google Scholar] [CrossRef]
- Norman, K.E.; Nymeyer, H. Indole localization in lipid membranes revealed by molecular simulation. Biophys. J. 2006, 91, 2046–2054. [Google Scholar] [CrossRef] [Green Version]
- Ulmschneider, M.B.; Sansom, M.S.P. Amino acid distributions in integral membrane protein structures. Biochim. Biophys. Acta-Biomembr. 2001, 1512, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Landolt-Marticorena, C.; Williams, K.A.; Deber, C.M.; Reithmeier, R.A.F. Non-random distribution of amino acids in the transmembrane segments of human type I single span membrane proteins. J. Mol. Biol. 1993, 229, 602–608. [Google Scholar] [CrossRef]
- Sanderson, J.M. Peptide-lipid interactions: Insights and perspectives. Org. Biomol. Chem. 2005, 3, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Yau, W.M.; Wimley, W.C.; Gawrisch, K.; White, S.H. The preference of tryptophan for membrane interfaces. Biochemistry 1998, 37, 14713–14718. [Google Scholar] [CrossRef] [Green Version]
- Gaede, H.C.; Yau, W.M.; Gawrisch, K. Electrostatic contributions to indole-lipid interactions. J. Phys. Chem. B 2005, 109, 13014–13023. [Google Scholar] [CrossRef]
- Killian, J.A.; Von Heijne, G. How proteins adapt to a membrane-water interface. Trends Biochem. Sci. 2000, 25, 429–434. [Google Scholar] [CrossRef]
- White, S.H.; Wimley, W.C. Membrane Protein Folding and Satbility: Physical Principles. Annu. Rev. Biophys. Biomol. Struct. 1999, 28, 319–365. [Google Scholar] [CrossRef]
- Schiffer, M.; Chang, C.-H.; Stevens, F.J. The function of tryptophan residues in membrane proteins. Protein Eng. Des. Sel. 1992, 5, 213–214. [Google Scholar] [CrossRef]
- Chang, Y.; Zajicek, J.; Castellino, F.J. Role of tryptophan-63 of the kringle 2 domain of tissue-type plasminogen activator in its thermal stability, folding, and ligand binding properties. Biochemistry 1997, 36, 7652–7663. [Google Scholar] [CrossRef] [PubMed]
- Perraut, C.; Clottes, E.; Leydier, C.; Vial, C.; Marcillat, O. Role of quaternary structure in muscle creatine kinase stability: Tryptophan 210 is important for dimer cohesion. Proteins Struct. Funct. Genet. 1998, 32, 43–51. [Google Scholar] [CrossRef]
- Skoging, U.; Liljeström, P. Role of the C-terminal tryptophan residue for the structure-function of the alphavirus capsid protein. J. Mol. Biol. 1998, 279, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, P.; Aronsson, G.; Carlsson, U.; Jonsson, B.H. Tertiary structure formation at specific tryptophan side chains in the refolding of human carbonic anhydrase II. Biochemistry 1997, 36, 5142–5148. [Google Scholar] [CrossRef] [PubMed]
- Samanta, U.; Pal, D.; Chakrabarti, P. Environment of tryptophan side chains in proteins. Proteins Struct. Funct. Genet. 2000, 38, 288–300. [Google Scholar] [CrossRef]
- Harel, M.; Kleywegt, G.J.; Ravelli, R.B.; Silman, I.; Sussman, J.L. Crystal structure of an acetylcholinesterase-fasciculin complex: Interaction of a three-fingered toxin from snake venom with its target. Structure 1995, 3, 1355–1366. [Google Scholar] [CrossRef] [Green Version]
- Katz, B.A. Binding to Protein Targets of Peptidic Leads Discovered by Phage Display: Crystal Structures of Streptavidin-Bound Linear and Cyclic Peptide Ligands Containing the HPQ Sequence. Biochemistry 1995, 34, 15421–15429. [Google Scholar] [CrossRef]
- Yengo, C.M.; Fagnant, P.M.; Chrin, L.; Rovner, A.S.; Berger, C.L. Smooth muscle myosin mutants containing a single tryptophan reveal molecular interactions at the actin-binding interface. Proc. Natl. Acad. Sci. USA 1998, 95, 12944–12949. [Google Scholar] [CrossRef] [Green Version]
- Zeltins, A.; Schrempf, H. Specific interaction of the Streptomyces chitin-binding protein CHB1 with α-chitin The role of individual tryptophan residues. Eur. J. Biochem. 1997, 246, 557–564. [Google Scholar] [CrossRef]
- Langosch, D.; Heringa, J. Interaction of transmembrane helices by a knobs-into-holes packing characteristic of soluble coiled coils. Proteins Struct. Funct. Genet. 1998, 31, 150–159. [Google Scholar] [CrossRef]
- Arkin, I.T.; Brunger, A.T. Statistical analysis of predicted transmembrane α-helices. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 1998, 1429, 113–128. [Google Scholar] [CrossRef] [Green Version]
- Cowan, S.W.; Schirmer, T.; Rummel, G.; Steiert, M.; Ghosh, R.; Pauptit, R.A.; Jansonius, J.N.; Rosenbusch, J.P. Crystal structures explain functional properties of two E. coli porins. Nature 1992, 358, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Situ, A.J.; Kang, S.M.; Frey, B.B.; An, W.; Kim, C.; Ulmer, T.S. Membrane Anchoring of α-Helical Proteins: Role of Tryptophan. J. Phys. Chem. B 2018, 122, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, D.; Mahalakshmi, R. Position—Specific contribution of interface tryptophans on membrane protein energetics. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 451–457. [Google Scholar] [CrossRef]
- McDonald, S.K.; Fleming, K.G. Aromatic Side Chain Water-to-Lipid Transfer Free Energies Show a Depth Dependence across the Membrane Normal. J. Am. Chem. Soc. 2016, 138, 7946–7950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, H.; Rinehart, D.; Tamm, L.K. Membrane depth-dependent energetic contribution of the tryptophan side chain to the stability of integral membrane proteins. Biochemistry 2013, 52, 4413–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridder, A.; Skupjen, P.; Unterreitmeier, S.; Langosch, D. Tryptophan supports interaction of transmembrane helices. J. Mol. Biol. 2005, 354, 894–902. [Google Scholar] [CrossRef]
- Bogan, A.A.; Thorn, K.S. Anatomy of hot spots in protein interfaces. J. Mol. Biol. 1998, 280, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killian, J.A.; Salemink, I.; De Planque, M.R.R.; Lindblom, G.; Koeppe, R.E.; Greathouse, D.V. Induction of nonbilayer structures in diacylphosphatidylcholine model membranes by transmembrane α-helical peptides: Importance of hydrophobic mismatch and proposed role of tryptophans. Biochemistry 1996, 35, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Killian, J.A. Synthetic peptides as models for intrinsic membrane proteins. FEBS Lett. 2003, 555, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Petersen, F.N.R.; Jensen, M.; Nielsen, C.H. Interfacial tryptophan residues: A role for the cation-π effect? Biophys. J. 2005, 89, 3985–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Planque, M.R.R.; Greathouse, D.V.; Koeppe, R.E.; Schäfer, H.; Marsh, D.; Killian, J.A. Influence of lipid/peptide hydrophobic mismatch on the thickness of diacylphosphatidylcholine bilayers. A 2H NMR and ESR study using designed transmembrane α-helical peptides and gramicidin A. Biochemistry 1998, 37, 9333–9345. [Google Scholar] [CrossRef] [PubMed]
- Braun, P.; Von Heijne, G. The aromatic residues Trp and phe have different effects on the positioning of a transmembrane helix in the microsomal membrane. Biochemistry 1999, 38, 9778–9782. [Google Scholar] [CrossRef] [PubMed]
- De Jesus, A.J.; Allen, T.W. The role of tryptophan side chains in membrane protein anchoring and hydrophobic mismatch. Biochim. Biophys. Acta-Biomembr. 2013, 1828, 864–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vostrikov, V.V.; Koeppe, R.E. Response of GWALP transmembrane peptides to changes in the tryptophan anchor positions. Biochemistry 2011, 50, 7522–7535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinnokrot, M.O.; Valeev, E.F.; Sherrill, C.D. Estimates of the ab initio limit for π-π interactions: The benzene dimer. J. Am. Chem. Soc. 2002, 124, 10887–10893. [Google Scholar] [CrossRef]
- Williams, D.H.; Stephens, E.; O’Brien, D.P.; Zhou, M. Understanding noncovalent interactions: Ligand binding energy and catalytic efficiency from ligand-induced reductions in motion within receptors and enzymes. Angew. Chem. Int. Ed. 2004, 43, 6596–6616. [Google Scholar] [CrossRef]
- Carter-Fenk, K.; Herbert, J.M. Electrostatics does not dictate the slip-stacked arrangement of aromatic π-π Interactions. Chem. Sci. 2020, 11, 6758–6765. [Google Scholar] [CrossRef] [PubMed]
- Frontera, A.; Gamez, P.; Mascal, M.; Mooibroek, T.J.; Reedijk, J. Putting anion-π interactions into perspective. Angew. Chem. Int. Ed. 2011, 50, 9564–9583. [Google Scholar] [CrossRef] [PubMed]
- Dawson, R.E.; Hennig, A.; Weimann, D.P.; Emery, D.; Ravikumar, V.; Montenegro, J.; Takeuchi, T.; Gabutti, S.; Mayor, M.; Mareda, J.; et al. Experimental evidence for the functional relevance of anion-π interactions. Nat. Chem. 2010, 2, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Mareda, J.; Matile, S. Anion-π slides for transmembrane transport. Chem. A Eur. J. 2009, 15, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Gorteau, V.; Julliard, M.D.; Matile, S. Hydrophilic anchors for transmembrane anion-π slides. J. Memb. Sci. 2008, 321, 37–42. [Google Scholar] [CrossRef]
- Cotelle, Y.; Lebrun, V.; Sakai, N.; Ward, T.R.; Matile, S. Anion-π enzymes. ACS Cent. Sci. 2016, 2, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, K.; Humbert-Droz, M.; Letrun, R.; Vauthey, E.; Wesolowski, T.A.; Sakai, N.; Matile, S. Ion Pair-π Interactions. J. Am. Chem. Soc. 2015, 137, 11047–11056. [Google Scholar] [CrossRef] [PubMed]
- Chuard, N.; Fujisawa, K.; Morelli, P.; Saarbach, J.; Winssinger, N.; Metrangolo, P.; Resnati, G.; Sakai, N.; Matile, S. Activation of Cell-Penetrating Peptides with Ionpair-π Interactions and Fluorophiles. J. Am. Chem. Soc. 2016, 138, 11264–11271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walrant, A.; Bauzá, A.; Girardet, C.; Alves, I.D.; Lecomte, S.; Illien, F.; Cardon, S.; Chaianantakul, N.; Pallerla, M.; Burlina, F.; et al. Ionpair-π interactions favor cell penetration of arginine/tryptophan-rich cell-penetrating peptides. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183098. [Google Scholar] [CrossRef]
- Purushotham, U.; Sastry, G.N. A comprehensive conformational analysis of tryptophan, its ionic and dimeric forms. J. Comput. Chem. 2014, 35, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Oliveira, A.F.; Batista, P.R.; Ducati, L.C.; Correra, T.C. Analyzing the N–H+⋯ π interactions of protonated tryptophan and phenylalkylamines using QTAIM, NCI, and NBO. Theor. Chem. Acc. 2020, 139, 1–9. [Google Scholar] [CrossRef]
- Khan, H.M.; MacKerell, A.D.; Reuter, N. Cation-π Interactions between Methylated Ammonium Groups and Tryptophan in the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2019, 15, 7–12. [Google Scholar] [CrossRef]
- Johnston, A.J.; Zhang, Y.; Busch, S.; Pardo, L.C.; Imberti, S.; McLain, S.E. Amphipathic solvation of indole: Implications for the role of tryptophan in membrane proteins. J. Phys. Chem. B 2015, 119, 5979–5987. [Google Scholar] [CrossRef] [Green Version]
- Iyer, B.R.; Zadafiya, P.; Vetal, P.V.; Mahalakshmi, R. Energetics of side-chain partitioning of β-signal residues in unassisted folding of a transmembrane β-barrel protein. J. Biol. Chem. 2017, 292, 12351–12365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimley, W.C.; White, S.H. Experimentally determined hydrophobicity scale for proteins at membrane interfaces. Nat. Struct. Biol. 1996, 3, 842–848. [Google Scholar] [CrossRef]
- White, S.H.; Wimley, W.C. Hydrophobic interactions of peptides with membrane interfaces. Biochim. Biophys. Acta-Rev. Biomembr. 1998, 1376, 339–352. [Google Scholar] [CrossRef] [Green Version]
- Klein-Seetharaman, J.; Oikawa, M.; Grimshaw, S.B.; Wirmer, J.; Duchardt, E.; Ueda, T.; Imoto, T.; Smith, L.J.; Dobson, C.M.; Schwalbe, H. Long-range interactions within a nonnative protein. Science 2002, 295, 1719–1722. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, Y.; Tanford, C. The solubility of amino acids and two glycine peptides in aqueous ethanol and dioxane solutions. Establishment of a hydrophobicity scale. J. Biol. Chem. 1971, 246, 2211–2217. [Google Scholar] [CrossRef]
- Fauchere, J.-L.; Pliska, V. Hydrophobic parameters pi of amino-acid side chains from the partitioning of N-acetyl-amino-acid amides. Eur. J. Med. Chem 1983, 18, 369–375. [Google Scholar]
- Chothia, C. The nature of the accessible and buried surfaces in proteins. J. Mol. Biol. 1976, 105, 1–12. [Google Scholar] [CrossRef]
- Wolfenden, R.; Andersson, L.; Cullis, P.M.; Southgate, C.C.B. Affinities of Amino Acid Side Chains for Solvent Water. Biochemistry 1981, 20, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Hessa, T.; Kim, H.; Bihlmaier, K.; Lundin, C.; Boekel, J.; Andersson, H.; Nilsson, I.M.; White, S.H.; Von Heijne, G. Recognition of transmembrane helices by the endoplasmic reticulum translocon. Nature 2005, 433, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.P.; Fleming, K.G. Side-chain hydrophobicity scale derived from transmembrane protein folding into lipid bilayers. Proc. Natl. Acad. Sci. USA 2011, 108, 10174–10177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfenden, R. Experimental measures of amino acid hydrophobicity and the topology of transmembrane and globular proteins. J. Gen. Physiol. 2007, 129, 357–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacCallum, J.L.; Drew Bennett, W.F.; Peter Tieleman, D. Distribution of amino acids in a lipid bilayer from computer simulations. Biophys. J. 2008, 94, 3393–3404. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.H.; Charlton, J.P. Interactions of Phosphatidylcholine Vesicles with 2-p-Toluidinylnaphthalene-6-sulfonate. Biochemistry 1972, 11, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Wimley, W.C.; White, S.H. Membrane Partitioning: Distinguishing Bilayer Effects from the Hydrophobic Effect. Biochemistry 1993, 32, 6307–6312. [Google Scholar] [CrossRef]
- Fernández-Vidal, M.; White, S.H.; Ladokhin, A.S. Membrane partitioning: “classical” and “nonclassical” hydrophobic effects. J. Membr. Biol. 2011, 239, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Mahadevi, A.S.; Sastry, G.N. Cation-π interaction: Its role and relevance in chemistry, biology, and material science. Chem. Rev. 2013, 113, 2100–2138. [Google Scholar] [CrossRef]
- Dougherty, D.A. The cation-π interaction. Acc. Chem. Res. 2013, 46, 885–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.C.; Dougherty, D.A. The cation-π interaction. Chem. Rev. 1997, 97, 1303–1324. [Google Scholar] [CrossRef]
- Schottel, B.L.; Chifotides, H.T.; Dunbar, K.R. Anion-π interactions. Chem. Soc. Rev. 2008, 37, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Gallivan, J.P.; Dougherty, D.A. Cation-π interactions in structural biology. Proc. Natl. Acad. Sci. USA 1999, 96, 9459–9464. [Google Scholar] [CrossRef] [Green Version]
- Flocco, M.M.; Mowbray, S.L. Planar stacking interactions of arginine and aromatic side-chains in proteins. J. Mol. Biol. 1994, 235, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Lucas, X.; Bauzá, A.; Frontera, A.; Quiñonero, D. A thorough anion-π interaction study in biomolecules: On the importance of cooperativity effects. Chem. Sci. 2016, 7, 1038–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarty, S.; Ung, A.R.; Moore, B.; Shore, J.; Alshamrani, M. A Comprehensive Analysis of Anion-Quadrupole Interactions in Protein Structures. Biochemistry 2018, 57, 1852–1867. [Google Scholar] [CrossRef]
- Mbaye, M.N.; Hou, Q.; Basu, S.; Teheux, F.; Pucci, F.; Rooman, M. A comprehensive computational study of amino acid interactions in membrane proteins. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Greathouse, D.V.; Andersen, O.S.; Koeppe, R.E. The preference of tryptophan for membrane interfaces: Insights from N-methylation of tryptophans in gramicidin channels. J. Biol. Chem. 2008, 283, 22233–22243. [Google Scholar] [CrossRef] [Green Version]
- Blaser, G.; Sanderson, J.M.; Wilson, M.R. Free-energy relationships for the interactions of tryptophan with phosphocholines. Org. Biomol. Chem. 2009, 7, 5119–5128. [Google Scholar] [CrossRef] [Green Version]
- Ippolito, J.A.; Alexander, R.S.; Christianson, D.W. Hydrogen bond stereochemistry in protein structure and function. J. Mol. Biol. 1990, 215, 457–471. [Google Scholar] [CrossRef]
- Bouchard, M.; Auger, M. Solvent history dependence of gramicidin-lipid interactions: A Raman and infrared spectroscopic study. Biophys. J. 1993, 65, 2484–2492. [Google Scholar] [CrossRef] [Green Version]
- Meulendijks, G.H.W.M.; Sonderkamp, T.; Dubois, J.E.; Nielen, R.J.; Kremers, J.A.; Buck, H.M. The different influences of ether and ester phospholipids on the conformation of gramicidin A. A molecular modelling study. Biochim. Biophys. Acta 1989, 979, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Salom, D.; Pérez-Payá, E.; Pascal, J.; Abad, C. Environment- and sequence-dependent modulation of the double-stranded to single-stranded conformational transition of gramicidin A in membranes. Biochemistry 1998, 37, 14279–14291. [Google Scholar] [CrossRef] [PubMed]
- Wimley, W.C.; White, S.H.; Chang, G.G. Partitioning of Tryptophan Side-Chain Analogs between Water and Cyclohexane. Biochemistry 1992, 31, 12813–12818. [Google Scholar] [CrossRef]
- Ladokhin, A.S.; Jayasinghe, S.; White, S.H. How to measure and analyze tryptophan fluorescence in membranes properly, and why bother? Anal. Biochem. 2000, 285, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Tower, S.J.; Hetcher, W.J.; Myers, T.E.; Kuehl, N.J.; Taylor, M.T. Selective Modification of Tryptophan Residues in Peptides and Proteins Using a Biomimetic Electron Transfer Process. J. Am. Chem. Soc. 2020, 142, 9112–9118. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Dai, H.; Li, J.; Huang, X.; Wei, Z. The effects of biological environments on the electron-relay functionality of tryptophan residues in proteins. ChemPhysChem 2012, 13, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Korendovych, I.V.; Broos, J. Biosynthetic incorporation of the azulene moiety in proteins with high efficiency. Amino Acids 2015, 47, 213–216. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khemaissa, S.; Sagan, S.; Walrant, A. Tryptophan, an Amino-Acid Endowed with Unique Properties and Its Many Roles in Membrane Proteins. Crystals 2021, 11, 1032. https://doi.org/10.3390/cryst11091032
Khemaissa S, Sagan S, Walrant A. Tryptophan, an Amino-Acid Endowed with Unique Properties and Its Many Roles in Membrane Proteins. Crystals. 2021; 11(9):1032. https://doi.org/10.3390/cryst11091032
Chicago/Turabian StyleKhemaissa, Sonia, Sandrine Sagan, and Astrid Walrant. 2021. "Tryptophan, an Amino-Acid Endowed with Unique Properties and Its Many Roles in Membrane Proteins" Crystals 11, no. 9: 1032. https://doi.org/10.3390/cryst11091032
APA StyleKhemaissa, S., Sagan, S., & Walrant, A. (2021). Tryptophan, an Amino-Acid Endowed with Unique Properties and Its Many Roles in Membrane Proteins. Crystals, 11(9), 1032. https://doi.org/10.3390/cryst11091032