
Microindentation Hardness of Protein Crystals under Controlled Relative Humidity

Abstract
:
1. Introduction
2. Results and Discussion
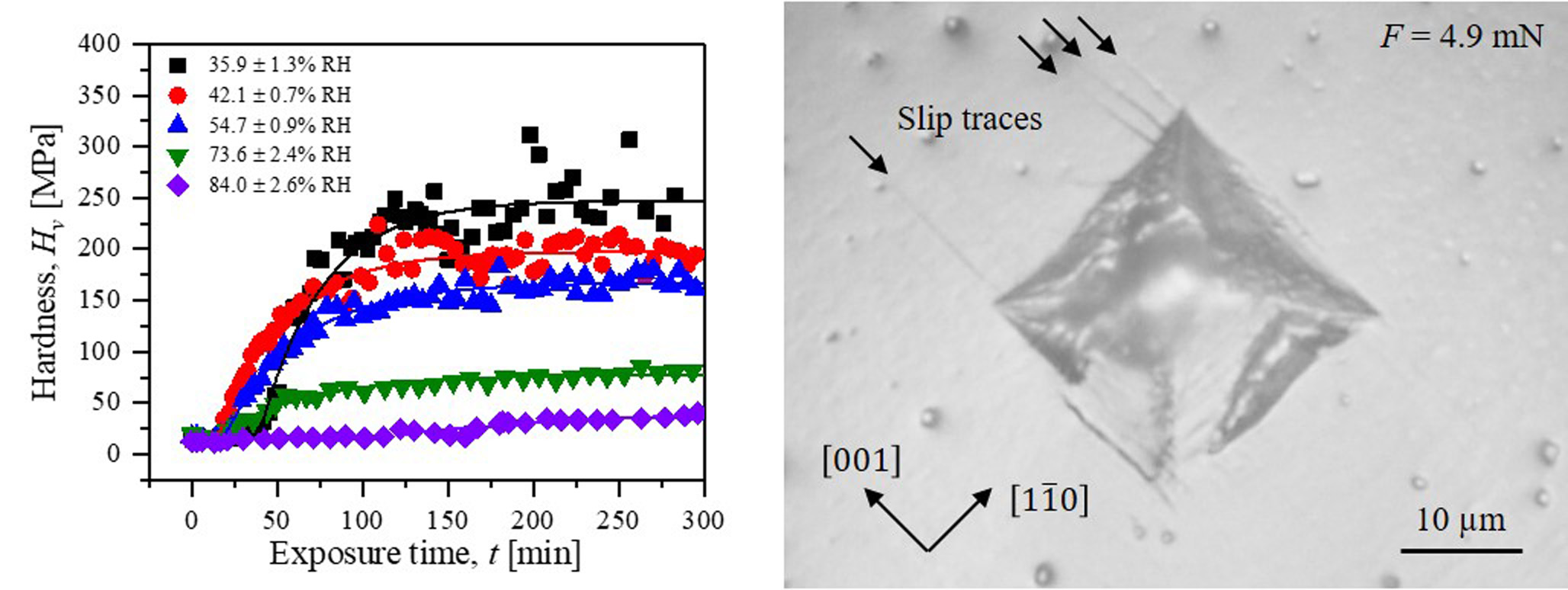
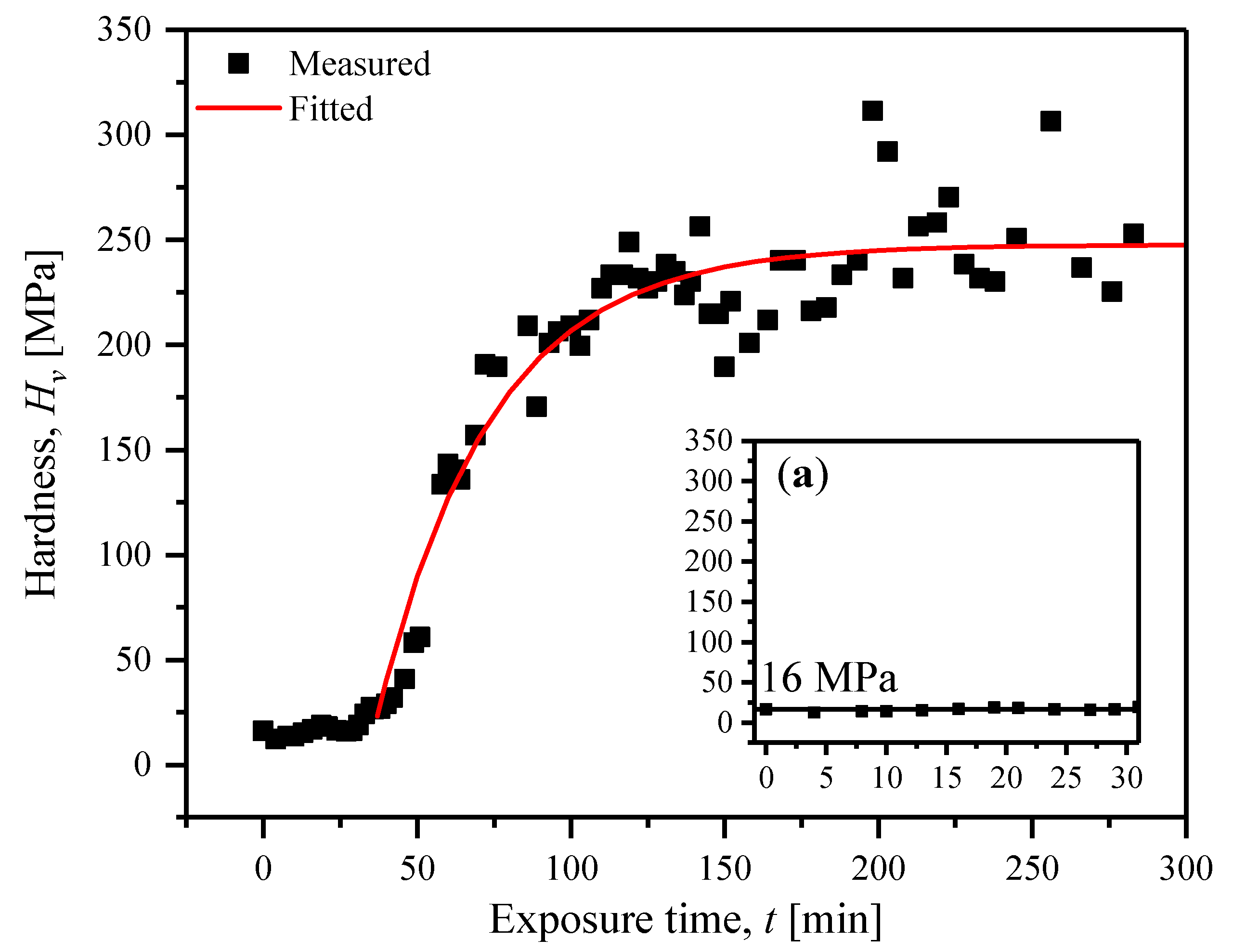
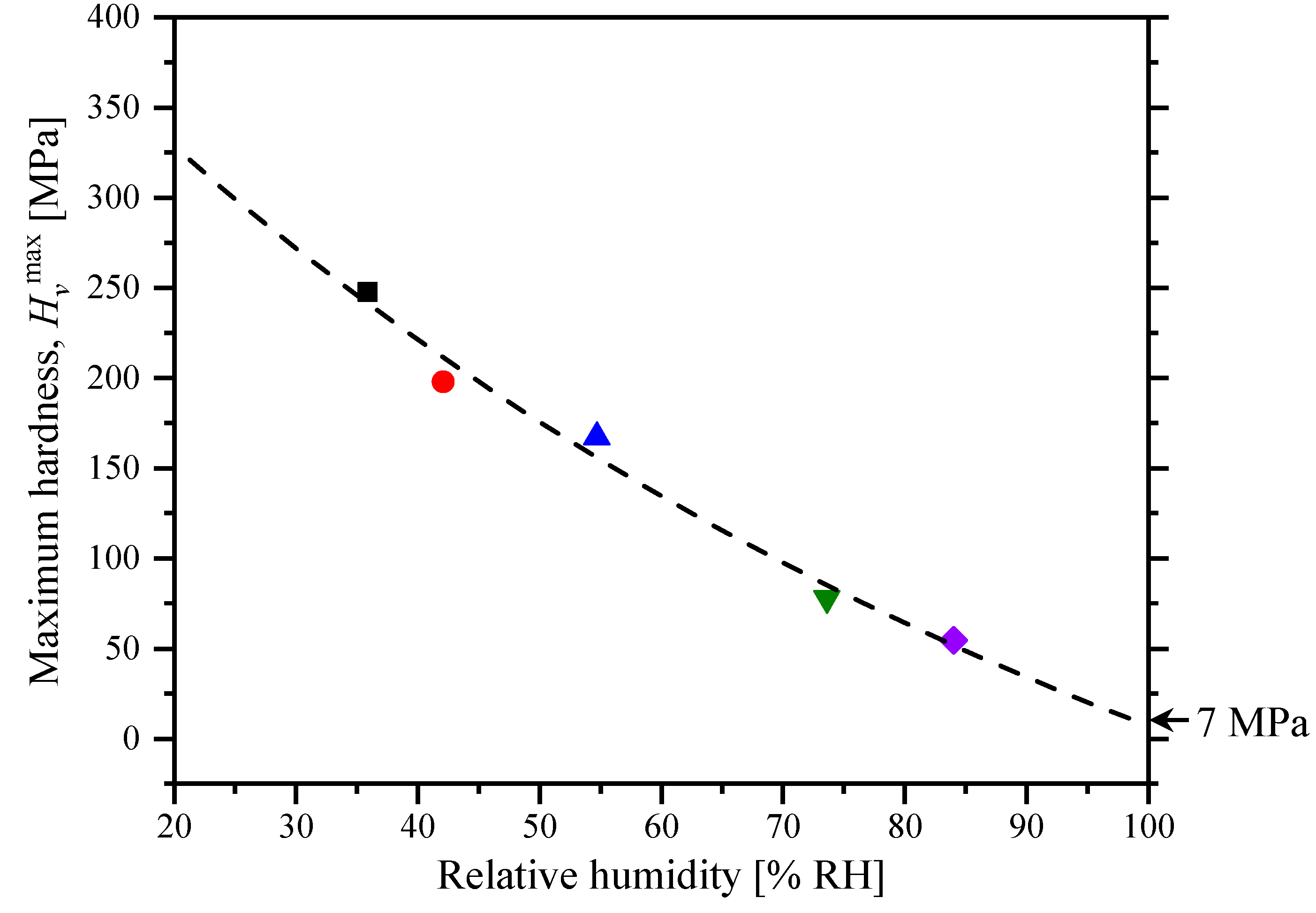
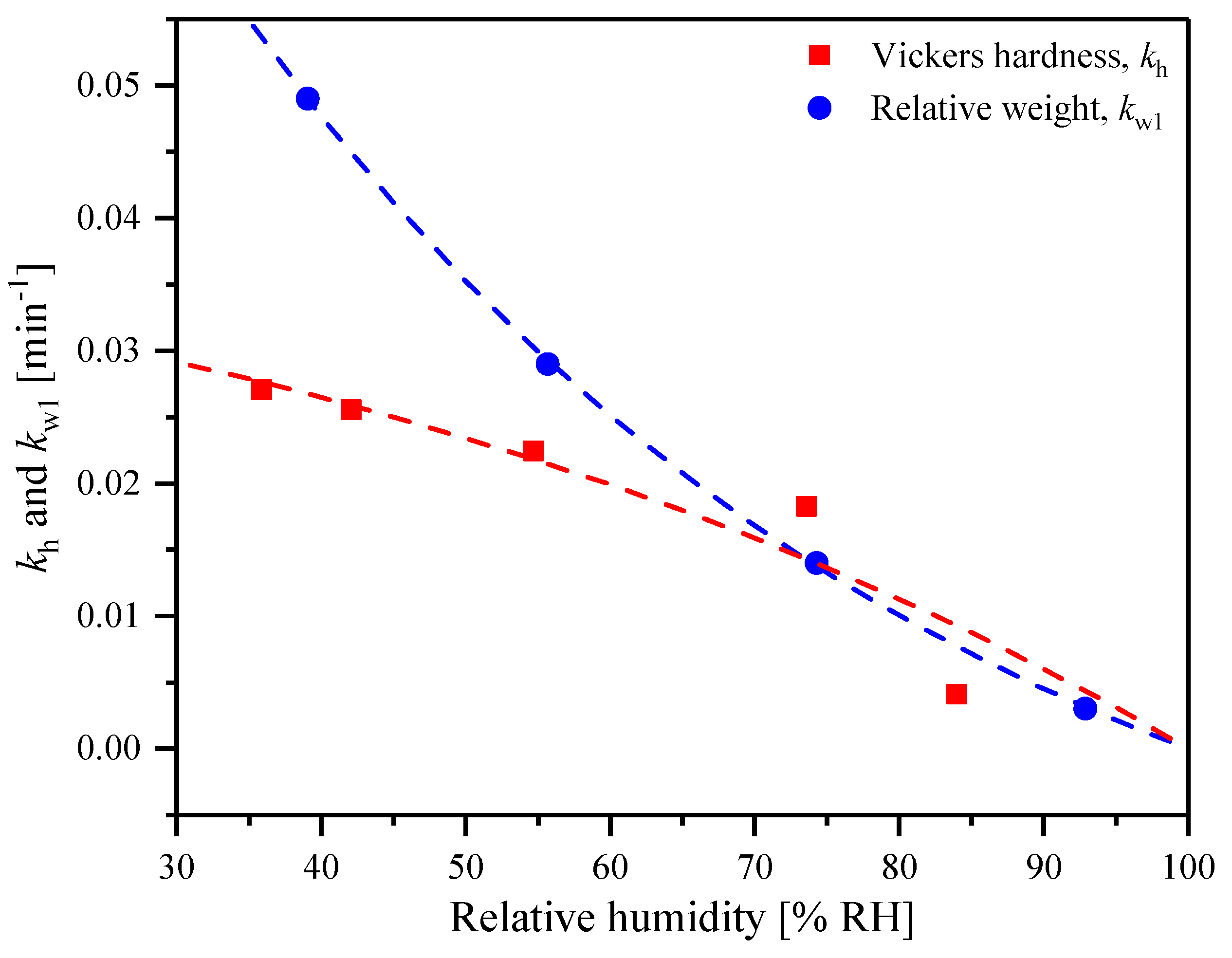
2.1. Hardness at Controlled Humidity
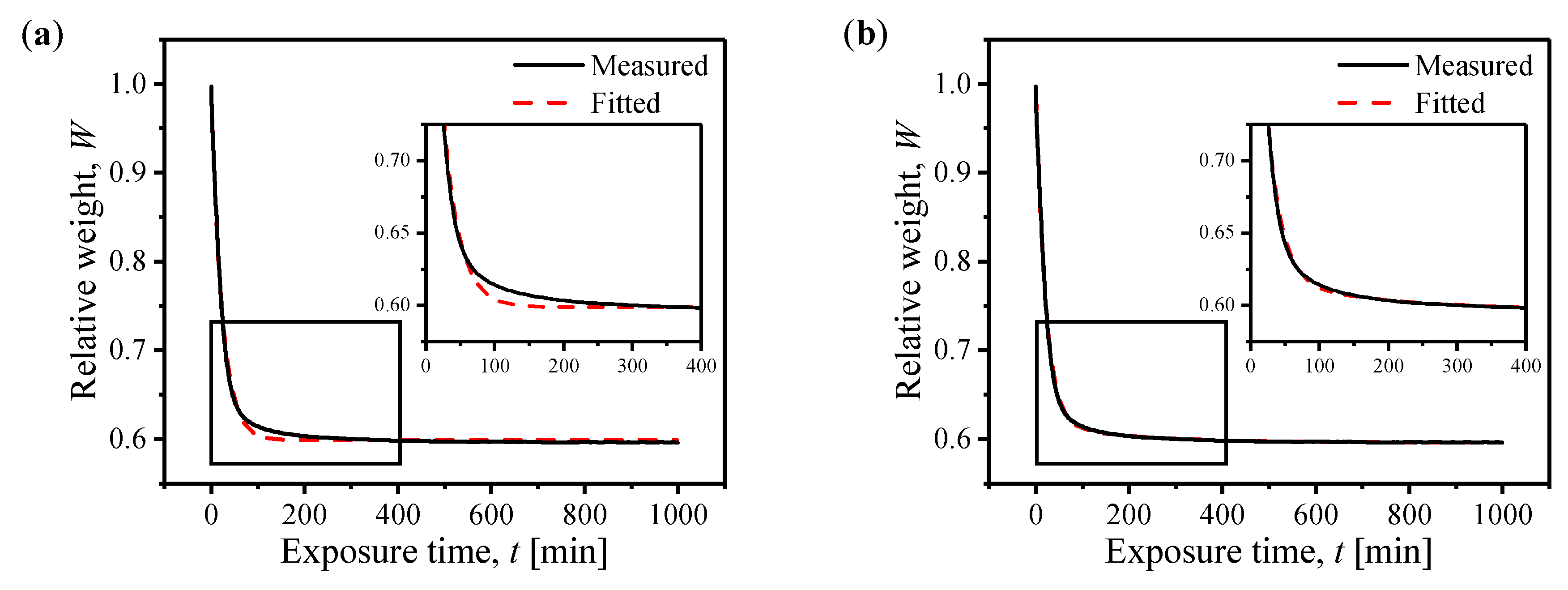
2.2. Evaporation of Intracrystalline Water
2.3. Dislocations and Peierls Stress
3. Materials and Methods
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Meyers, M.A.; Chawla, K.K. Mechanical Behavior of Materials, 2nd ed.; Cambridge University Press: Cambridge, UK, 2008; ISBN 9780521866750. [Google Scholar]
- Gilman, J.J. Chemistry and Physics of Mechanical Hardness, 1st ed.; John Wiley and Sons: Hoboken, NJ, USA, 2009; ISBN 9780470226520. [Google Scholar]
- Bembey, A.K.; Oyen, M.L.; Bushby, A.J.; Boyde, A. Viscoelastic properties of bone as a function of hydration state determined by nanoindentation. Philos. Mag. 2006, 86, 5691–5703. [Google Scholar] [CrossRef]
- Bembey, A.K.; Bushby, A.J.; Boyde, A.; Ferguson, V.L.; Oyen, M.L. Hydration effects on the micro-mechanical properties of bone. J. Mater. Res. 2006, 21, 1962–1968. [Google Scholar] [CrossRef]
- Oyen, M.L. Poroelastic nanoindentation responses of hydrated bone. J. Mater. Res. 2008, 23, 1307–1314. [Google Scholar] [CrossRef]
- Oyen, M.L. Nanoindentation of hydrated materials and tissues. Curr. Opin. Solid. State Mater. Sci. 2015, 19, 317–323. [Google Scholar] [CrossRef]
- McPherson, A. Crystallization of Biological Macromolecules; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1999; ISBN 9780879695279. [Google Scholar]
- Matthews, B.W. Solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–497. [Google Scholar] [CrossRef]
- Vilenchik, L.Z.; Griffith, J.P.; Clair, N.S.; Navia, M.A.; Margolin, A.L. Protein crystals as novel microporous materials. J. Am. Chem. Soc. 1998, 120, 4290–4294. [Google Scholar] [CrossRef]
- Margolin, A.L.; Navia, M.A. Protein crystals as novel catalytic materials. Angew. Chem. Int. Ed. 2001, 40, 2204–2222. [Google Scholar] [CrossRef]
- Abe, S.; Ueno, T. Design of protein crystals in the development of solid biomaterials. RSC Adv. 2015, 5, 21366–21375. [Google Scholar] [CrossRef]
- Otting, G.; Liepinish, E.; Wuthrich, K. Protein hydration in aqueous solution. Science 1991, 254, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.N.; Kachalova, G.S.; Evtodienko, V.U.; Lanina, N.F.; Morozova, T.Y. Permeability of lysozyme tetragonal crystals to water. Eur. Biophys. J. 1995, 24, 93–98. [Google Scholar] [CrossRef]
- Jones, M.J.; Ulrich, J. Are different protein crystal modifications polymorphs? A discussion. Chem. Eng. Technol. 2010, 33, 1571–1576. [Google Scholar] [CrossRef]
- Morozov, V.N.; Morozova, T.Y. Viscoelastic properties of protein crystals: Triclinic crystals of hen egg white lysozyme in different conditions. Biopolymers 1981, 20, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.N.; Morozova, T.Y.; Kachalova, G.S.; Myachin, E.T. Interpretation of water desorption isotherms of lysozyme. Int. J. Biol. Macromol. 1988, 10, 329–336. [Google Scholar] [CrossRef]
- Zenchenko, T.A.; Pozharskii, E.V.; Morozov, V.N. A magnetic micromethod to measure Young’s modulus of protein crystals and other polymer materials. J. Biochem. Biophys. Methods 1996, 33, 207–215. [Google Scholar] [CrossRef]
- Tachibana, M.; Kojima, K.; Ikuyama, R.; Kobayashi, Y.; Ataka, M. Sound velocity and dynamic elastic constants of lysozyme single crystals. Chem. Phys. Lett. 2000, 332, 259–264. [Google Scholar] [CrossRef]
- Tachibana, M.; Kojima, K.; Ikuyama, R.; Kobayashi, Y.; Ataka, M. Erratum to: Sound velocity and dynamic elastic constants of lysozyme single crystals. Chem. Phys. Lett. 2002, 354, 360. [Google Scholar] [CrossRef]
- Tachibana, M.; Koizumi, H.; Kojima, K. Effect of intracrystalline water on longitudinal sound velocity in tetragonal hen-egg-white lysozyme crystals. Phys. Rev. E 2004, 69, 051921. [Google Scholar] [CrossRef] [PubMed]
- Caylor, C.L.; Speziale, S.; Kriminski, S.; Duffy, T. Measuring the elastic properties of protein crystals by Brillouin scattering. J. Cryst. Growth 2001, 232, 498–501. [Google Scholar] [CrossRef]
- Speziale, S.; Jiang, F.; Caylor, C.L.; Kriminski, S.; Zha, C.S.; Thorne, R.E.; Duffy, T.S. Sound velocity and elasticity of tetragonal lysozyme crystals by Brillouin spectroscopy. Biophys. J. 2003, 85, 3202–3213. [Google Scholar] [CrossRef]
- Hashimoto, E.; Aoki, Y.; Seshimo, Y.; Sasanuma, K.; Ike, Y.; Kojima, S. Dehydration process of protein crystals by micro-brillouin scattering. Jpn. J. Appl. Phys. 2008, 47, 3839–3842. [Google Scholar] [CrossRef]
- Koizumi, H.; Tachibana, M.; Kojima, K. Elastic constants in tetragonal hen egg-white lysozyme crystals containing large amount of water. Phys. Rev. E 2009, 79, 061917. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, N.; Tsukashima, S.; Fujii, D.; Tachibana, M.; Koizumi, H.; Wako, K.; Kojima, K. Elastic constants in orthorhombic hen egg-white lysozyme crystals. Phys. Rev. E 2014, 89, 012714. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, H.; Tachibana, M.; Kojima, K. Observation of all the components of elastic constants using tetragonal hen egg-white lysozyme crystals dehydrated at 42% relative humidity. Phys. Rev. E 2006, 73. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, M.; Kobayashi, Y.; Shimazu, T.; Ataka, M.; Kojima, K. Growth and mechanical properties of lysozyme crystals. J. Cryst. Growth 1999, 198, 661–664. [Google Scholar] [CrossRef]
- Koizumi, H.; Tachibana, M.; Kawamoto, H.; Kojima, K. Temperature dependence of microhardness of tetragonal hen-egg-white lysozyme single crystals. Philos. Mag. 2004, 84, 2961–2968. [Google Scholar] [CrossRef]
- Koizumi, H.; Kawamoto, H.; Tachibana, M.; Kojima, K. Effect of intracrystalline water on micro-Vickers hardness in tetragonal hen egg-white lysozyme single crystals. J. Phys. D 2008, 41, 074019. [Google Scholar] [CrossRef]
- Tachibana, M.; Koizumi, H.; Izumi, K.; Kajiwara, K.; Kojima, K. Identification of dislocations in large tetragonal hen egg-white lysozyme crystals by synchrotron white-beam topography. J. Synchrotron Radiat. 2003, 10, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Mukobayashi, Y.; Kitajima, N.; Yamamoto, Y.; Kajiwara, K.; Sugiyama, H.; Hirano, K.; Kojima, K.; Tachibana, M. Observation of dislocations in hen egg-white lysozyme crystals by synchrotron monochromatic-beam X-ray topography. Phys. Stat. Sol. 2009, 206, 1825–1828. [Google Scholar] [CrossRef]
- Suzuki, R.; Kishi, T.; Tsukashima, S.; Tachibana, M.; Wako, K.; Kojima, K. Hardness and slip systems of orthorhombic hen egg-white lysozyme crystals. Philos. Mag. 2016, 96, 2930–2942. [Google Scholar] [CrossRef]
- Tait, S.; White, E.T.; Litster, J.D. Mechanical characterization of protein crystals. Part. Part. Syst. Charact. 2008, 25, 266–276. [Google Scholar] [CrossRef]
- Nanev, C.N.; Dimitrov, I.; Tsekova, D. Adhesion of protein crystals: Measurement of the detachment force. Cryst. Res. Technol. 2006, 41, 505–509. [Google Scholar] [CrossRef]
- Nanev, C.N. Brittleness of protein crystals. Cryst. Res. Technol. 2012, 47, 922–927. [Google Scholar] [CrossRef]
- Suzuki, R.; Koizumi, H.; Kojima, K.; Fukuyama, S.; Arai, Y.; Tsukamoto, K.; Suzuki, Y.; Tachibana, M. Characterization of grown-in dislocations in high-quality glucose isomerase crystals by synchrotron monochromatic-beam X-ray topography. J. Cryst. Growth 2017, 468, 299–304. [Google Scholar] [CrossRef]
- Zamiri, A.; De, S. Modeling the mechanical response of tetragonal lysozyme crystals. Langmuir 2010, 26, 4251–4257. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.K.; Zewail, A.H. Dynamics of water in biological recognition. Chem. Rev. 2004, 104, 2099–2123. [Google Scholar] [CrossRef] [PubMed]
- Bray, Y.L.; Prat, M. Three-dimensional pore network simulation of drying in capillary porous media. Int. J. Heat Mass Tran. 1999, 42, 4207–4224. [Google Scholar] [CrossRef]
- Yiotis, A.G.; Tsimpanogiannis, I.N.; Stubos, A.K.; Yortsos, Y.C. Pore-network study of the characteristic periods in the drying of porous materials. J. Colloid. Interface Sci. 2006, 297, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, P.; Assouline, S.; Or, D. Characteristic lengths affecting evaporative drying of porous media. Phys. Rev. E 2008, 77. [Google Scholar] [CrossRef] [PubMed]
- Yiotis, A.G.; Salin, D.; Tajer, E.S.; Yortsos, Y.C. Drying in porous media with gravity-stabilized fronts: Experimental results. Phys. Rev. E 2012, 86. [Google Scholar] [CrossRef] [PubMed]
- Ebenstein, D.M.; Pruitt, L.A. Nanoindentation of biological materials. Nano Today 2006, 1, 26–33. [Google Scholar] [CrossRef]
- Ferguson, V.L. Deformation partitioning provides insight into elastic, plastic, and viscous contributions to bone materials behavior. J. Mech. Behav. Biomed. 2009, 2, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Oyen, M.L. Nanoindentation of biological and biomimetic materials. Exp. Tech. 2013, 37, 73–87. [Google Scholar] [CrossRef]
- Hirth, J.P.; Lothe, J. Theory of Dislocations, 2nd ed.; Wiley: Hoboken, NY, USA, 1982; ISBN 9780894646171. [Google Scholar]
- Hull, D.; Bacon, D.J. Introduction to Dislocations, 5th ed.; Butterworth-Heinemann: Oxford, UK, 2011; ISBN 9780080966724. [Google Scholar]
- Tachibana, M.; Kojima, K. Growth, Defects and mechanical properties of protein single crystals. Curr. Top. Cryst. Growth Res. 2002, 6, 35–49. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Humidity [% RH] | [min−1] | [MPa] |
|---|---|---|
| 35.9 | 0.027 | 247.6 |
| 42.1 | 0.026 | 197.8 |
| 54.7 | 0.022 | 167.2 |
| 73.6 | 0.018 | 77.8 |
| 84.0 | 0.004 | 54.7 |
| Relative Humidity [%] | [min−1] | [min−1] |
|---|---|---|
| 39.1 | 0.049 | 0.006 |
| 55.7 | 0.029 | - |
| 74.3 | 0.014 | - |
| 92.9 | 0.003 | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kishi, T.; Suzuki, R.; Shigemoto, C.; Murata, H.; Kojima, K.; Tachibana, M. Microindentation Hardness of Protein Crystals under Controlled Relative Humidity. Crystals 2017, 7, 339. https://doi.org/10.3390/cryst7110339
Kishi T, Suzuki R, Shigemoto C, Murata H, Kojima K, Tachibana M. Microindentation Hardness of Protein Crystals under Controlled Relative Humidity. Crystals. 2017; 7(11):339. https://doi.org/10.3390/cryst7110339
Chicago/Turabian StyleKishi, Takeharu, Ryo Suzuki, Chika Shigemoto, Hidenobu Murata, Kenichi Kojima, and Masaru Tachibana. 2017. "Microindentation Hardness of Protein Crystals under Controlled Relative Humidity" Crystals 7, no. 11: 339. https://doi.org/10.3390/cryst7110339
APA StyleKishi, T., Suzuki, R., Shigemoto, C., Murata, H., Kojima, K., & Tachibana, M. (2017). Microindentation Hardness of Protein Crystals under Controlled Relative Humidity. Crystals, 7(11), 339. https://doi.org/10.3390/cryst7110339