Conversion of Wheat Bran to Xylanases and Dye Adsorbent by Streptomyces thermocarboxydus




Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Xylanase Assay
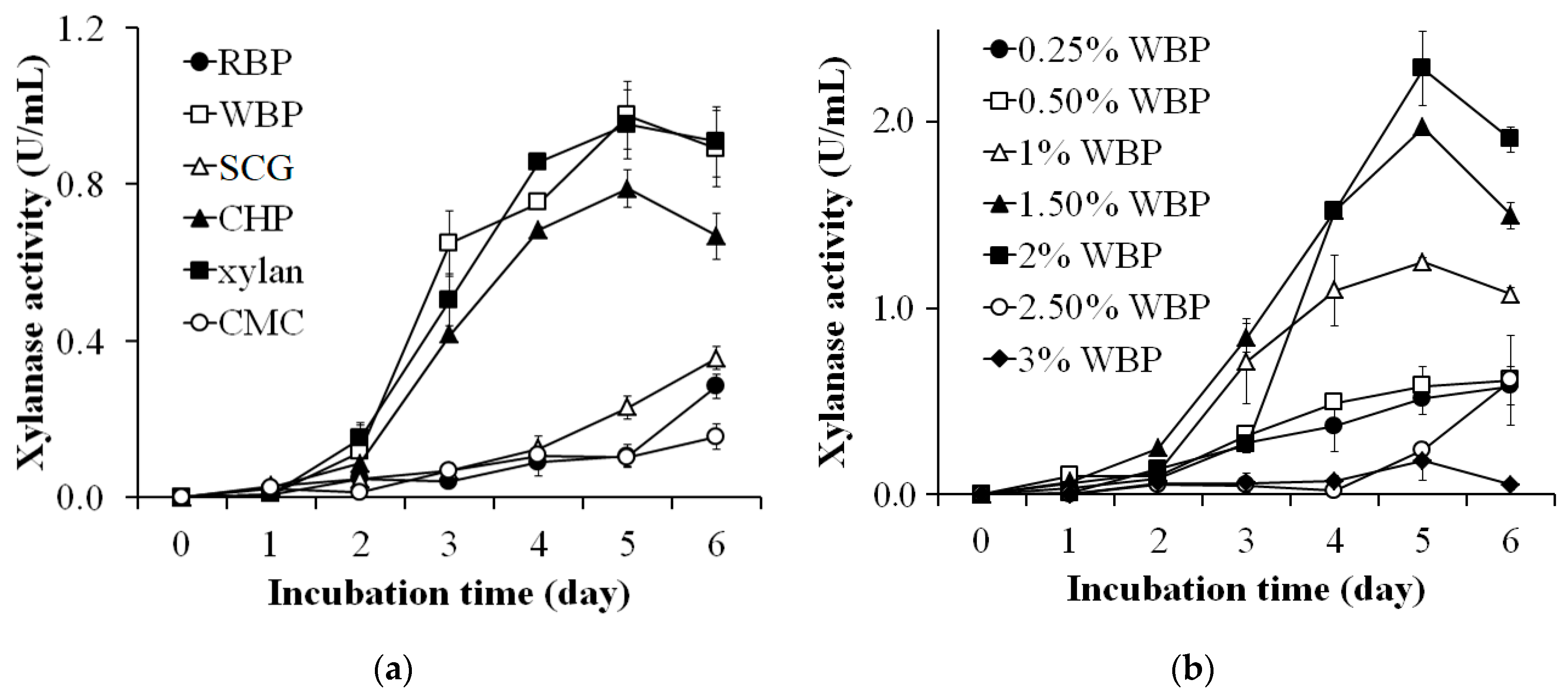
2.3. Agro-Byproducts as the Sole Carbon Source for Xylanase Production
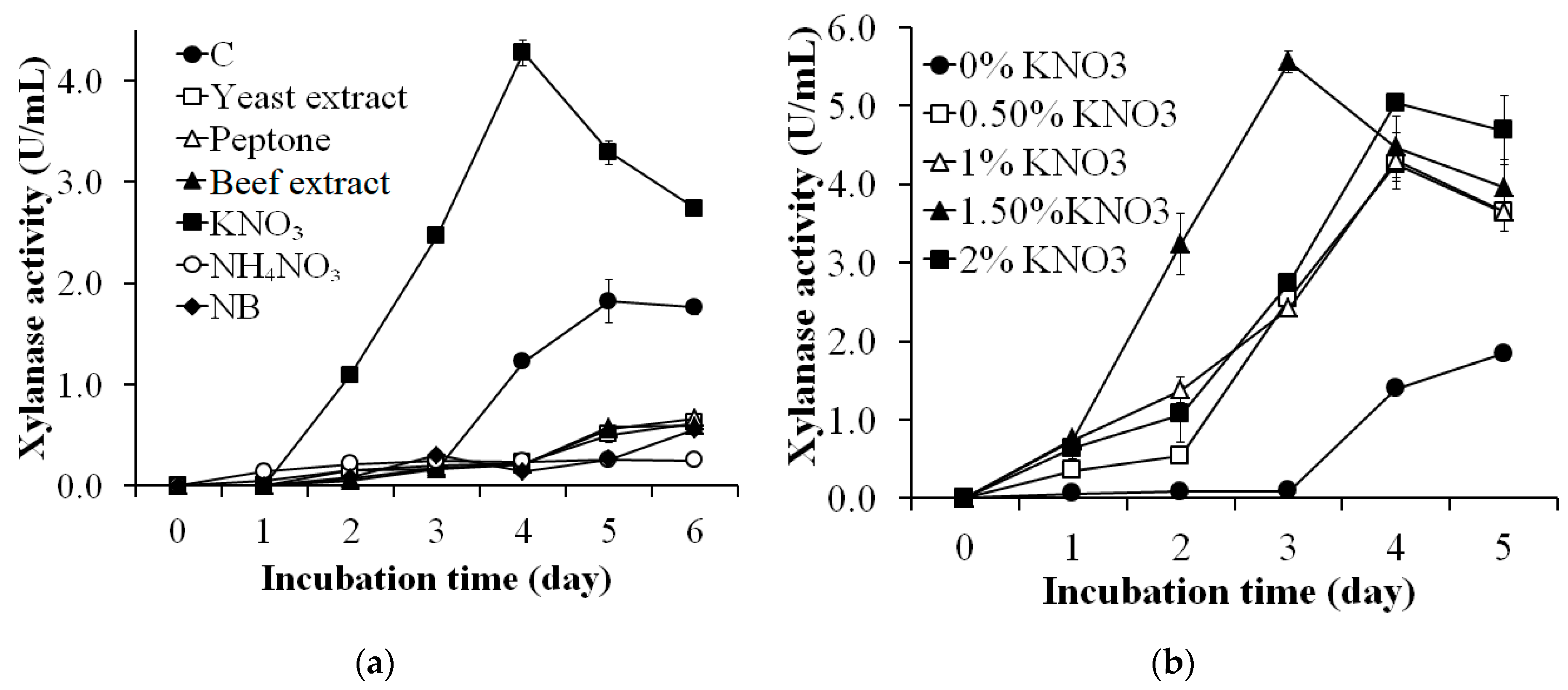
2.4. Effects of Nitrogen Sources on Xylanase Production
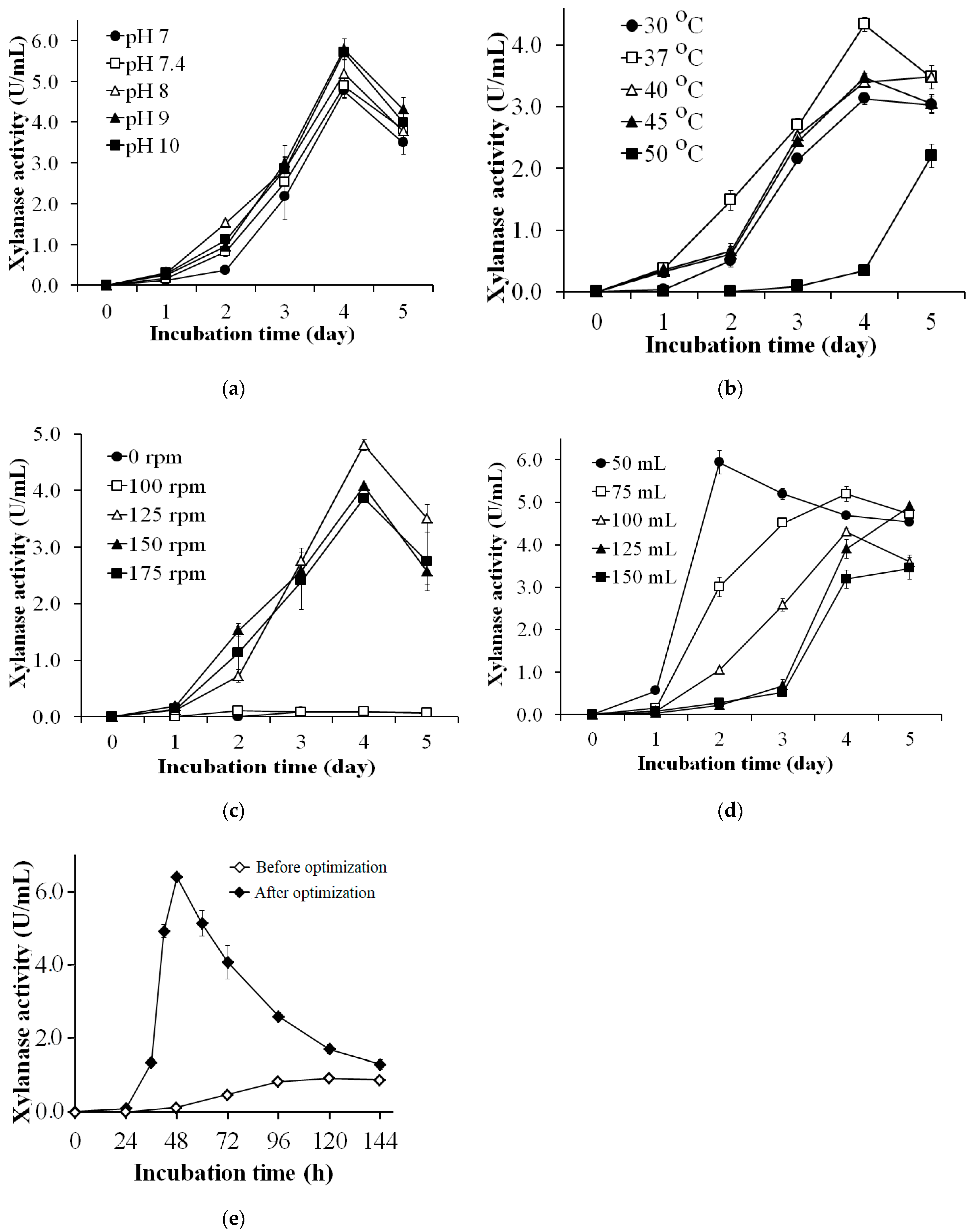
2.5. Effects of Culture Conditions on Xylanase Production
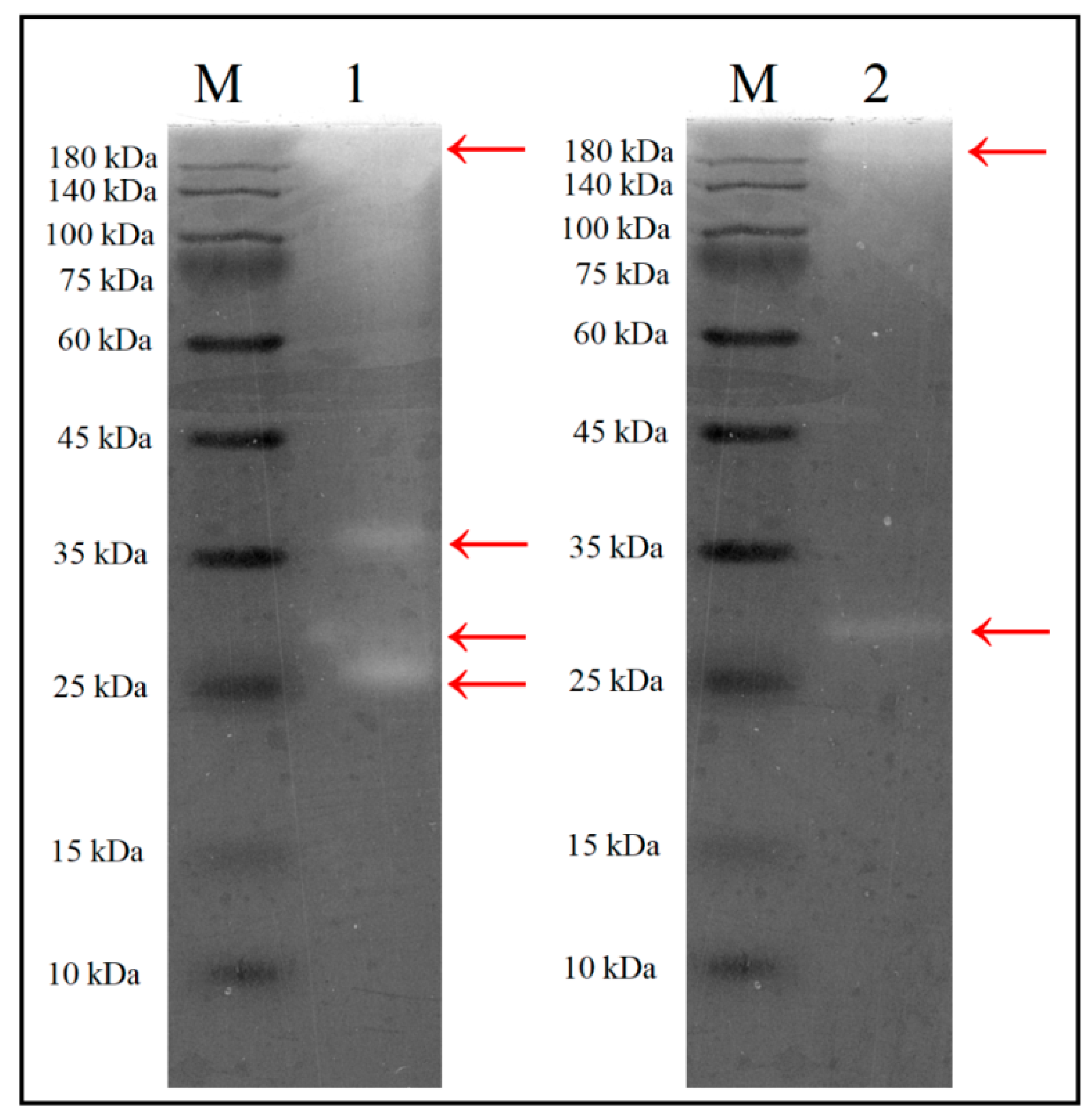
2.6. Determination of Molecular Weight (MW) of the Xylanases
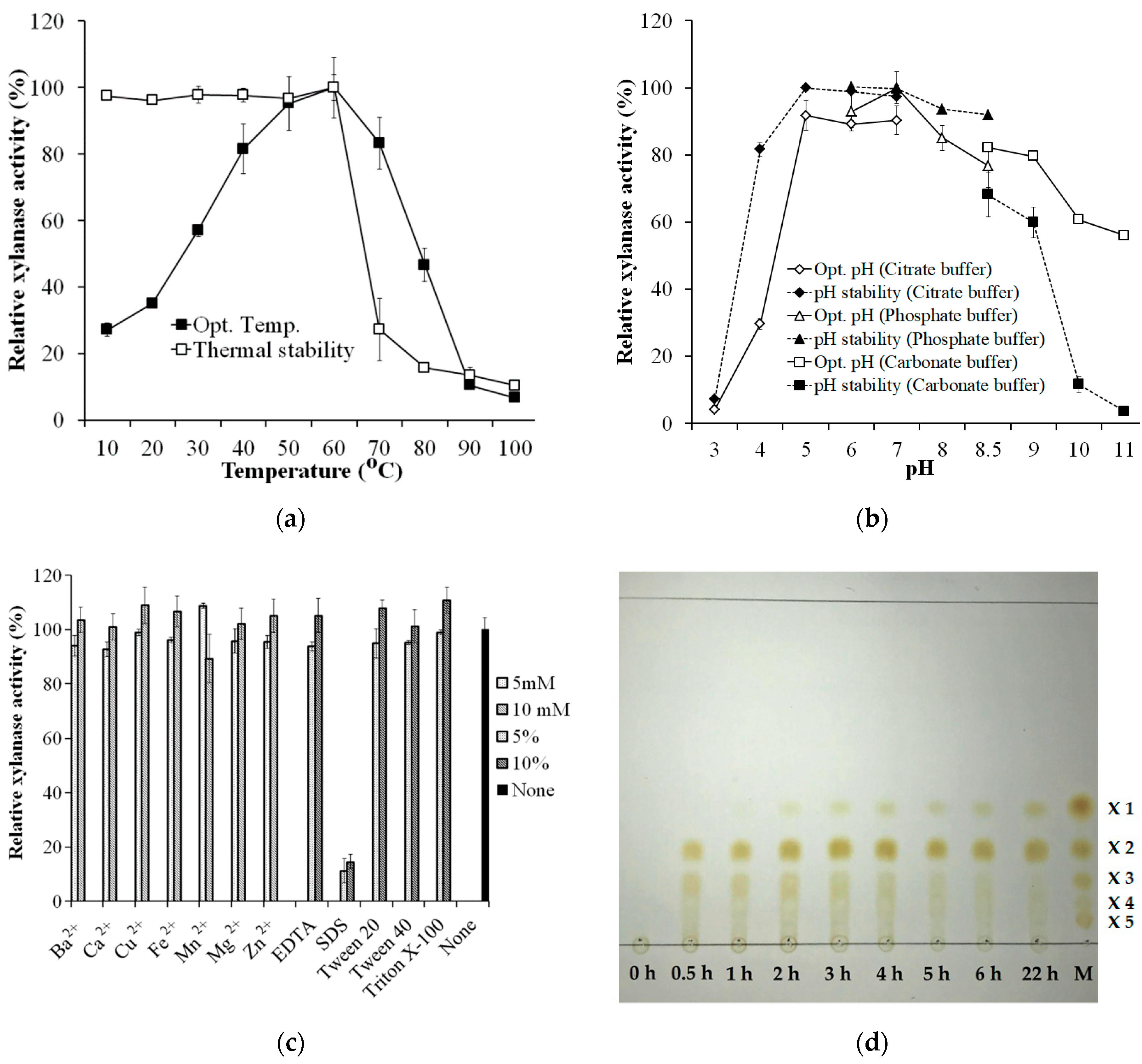
2.7. Effects of Temperature and pH
2.8. Effect of Divalent Metal Ions, Surfactants, and Ethylenediaminetetraacetic Acid (EDTA)
2.9. Xylan Hydrolysis
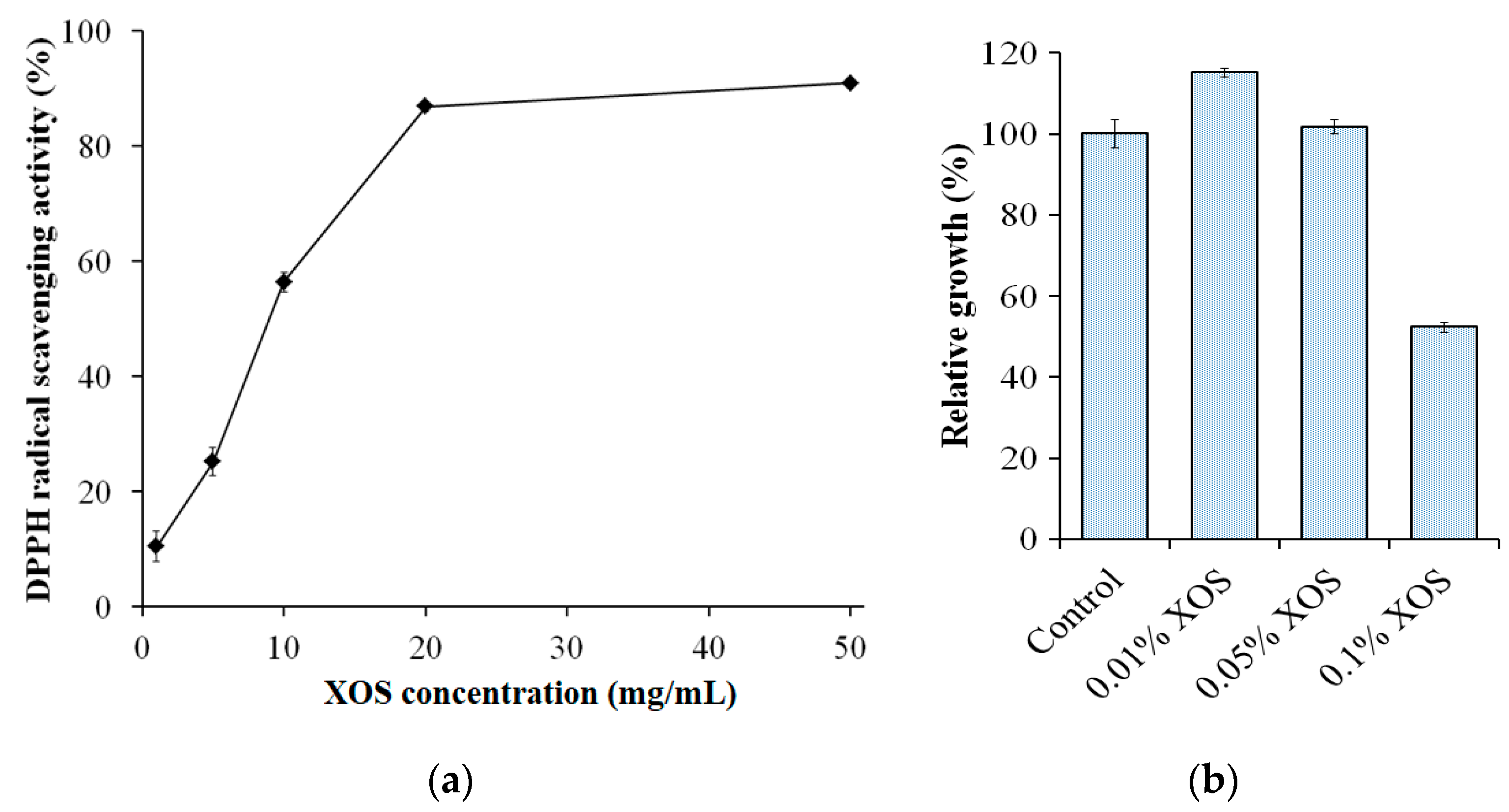
2.10. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Activity Assay
2.11. Growth Effect Assay
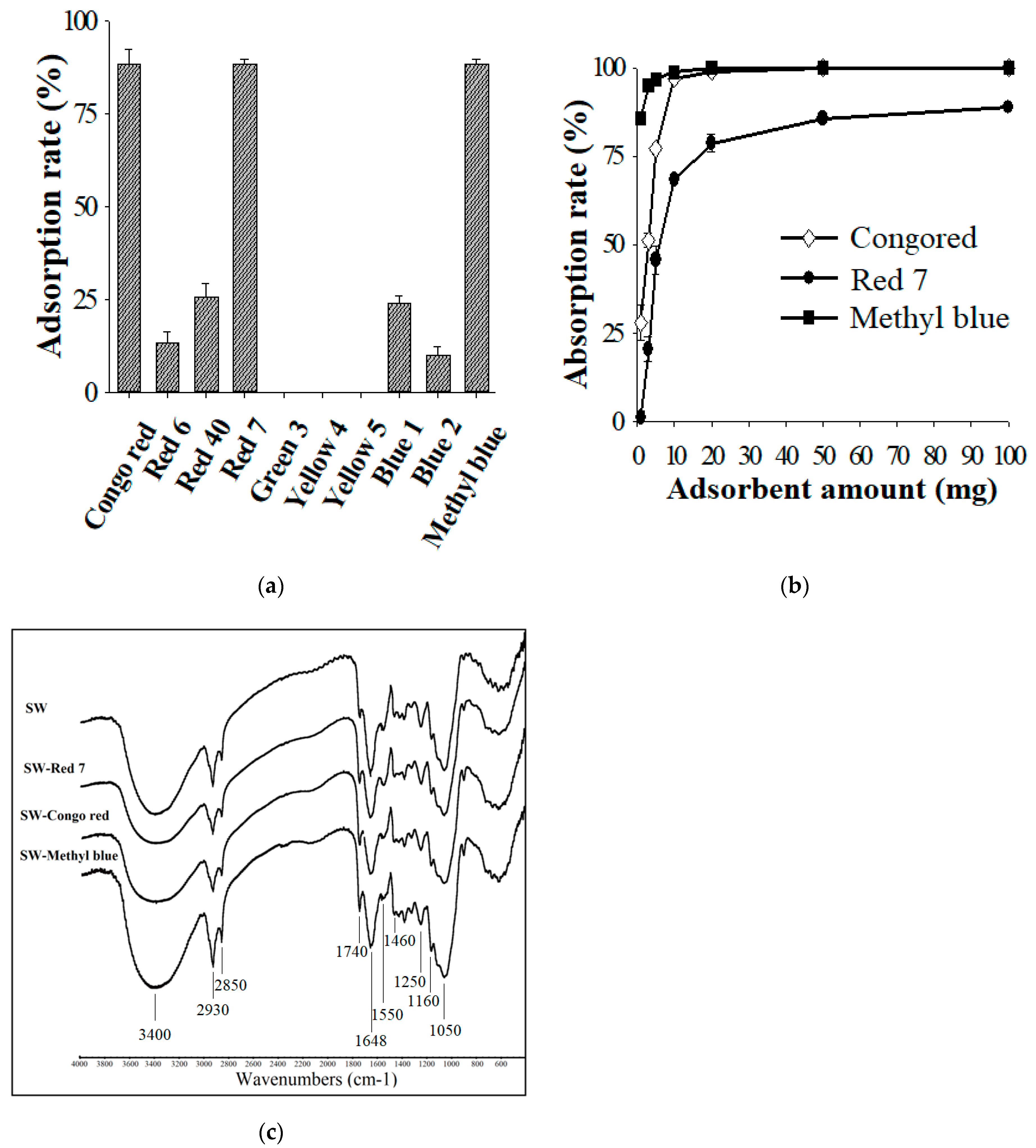
2.12. Dye Adsorption Assay
2.13. The Fourier-Transform Infrared Spectroscopy (FTIR) Analysis
2.14. Statistical Analysis
3. Results and Discussion
3.1. Agro-Byproducts as the Sole Carbon Source for the Xylanase Production
3.2. Effects of Nitrogen Source on Xylanase Production
3.3. Effects of Culture Conditions on Xylanase Production
3.4. Molecular Weight Determination of the Xylanases
3.5. Biochemical Characterization of Crude Enzyme Cocktail
3.6. Antioxidant Activity and the Growth Effect on Lactic Acid Bacteria of Xylan Hydrolysate
3.7. Utilization of Solid Waste from the WBP Fermentation as a Dye Adsorbent
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, H.; Yi, N.; Zhao, S.; Qaseem, M.F.; Zheng, B.; Li, H.; Feng, J.; Wu, A. Characterization of hemicelluloses in sugarcane (Saccharum spp. hybrids) culm during xylogenesis. Int. J. Biol. Macromol. 2020, 165, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from enzymatic degradation of cellulose and hemicellulose to fermentable sugars—A review. Biomass Bioener. 2020, 134, 105481. [Google Scholar] [CrossRef]
- Balakshin, M.; Capanema, E.; Gracz, H.; Chang, H.; Jameel, H. Quantification of lignin–carbohydrate linkages with high-resolution NMR spectroscopy. Planta 2011, 233, 1097–1110. [Google Scholar] [CrossRef]
- Malgas, S.; Mafa, M.S.; Mkabayi, L.; Pletschke, B. A mini review of xylanolytic enzymes with regards to their synergistic interactions during hetero-xylan degradation. World J. Microb. Biot. 2019, 35, 187. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Wu, J.C. Microbial exo-xylanases: A mini review. Appl. Biochem. Biotechnol. 2014, 174, 81–92. [Google Scholar] [CrossRef]
- Park, S.H.; Pham, T.T.H.; Kim, T.H. Effects of additional xylanase on saccharification and ethanol fermentation of ammonia-pretreated corn stover and rice straw. Energies 2020, 13, 4574. [Google Scholar] [CrossRef]
- Prasoulas, G.; Gentikis, A.; Konti, A.; Kalantzi, S.; Kekos, D.; Mamma, D. Bioethanol production from food waste applying the multienzyme system produced on-site by Fusarium oxysporum F3 and mixed microbial cultures. Fermentation 2020, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Dodd, D.; Cann, I.K. Enzymatic deconstruction of xylan for biofuel production. Glob. Chang. Biol. Bioenergy 2009, 1, 2–17. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Chaudhary, R.; Kaur, J.; Arya, S.K. Greener approach for pulp and paper industry by Xylanase and Laccase. Biocatal. Agric. Biotechnol. 2020, 25, 101604. [Google Scholar] [CrossRef]
- Sridevi, A.; Ramanjaneyulu, G.; Suvarnalatha Devi, P. Biobleaching of paper pulp with xylanase produced by Trichoderma asperellum. 3 Biotech. 2017, 7, 266. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, V.V.; Shrivastava, A.; Jain, R.K.; Mathur, R.M.; Gupta, R.; Kuhad, R.C. Xylanase and laccase based enzymatic kraft pulp bleaching reduces adsorbable organic halogen (AOX) in bleach effluents: A pilot scale study. Bioresour. Technol. 2014, 169, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D.; Khattak, F.; Hastie, P.; Bedford, M.R.; Olukosi, O.A. Xylanase and xylooligosaccharide prebiotic improve the growth performance and concentration of potentially prebiotic oligosaccharides in the ileum of broiler chickens. Br. Poult. Sci. 2020, 61, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordberg Karlsson, E.; Schmitz, E.; Linares-Pastén, J.A.; Adlercreutz, P. Endo-xylanases as tools for production of substituted xylooligosaccharides with prebiotic properties. Appl. Microbiol. Biot. 2018, 102, 9081–9088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valls, C.; Javier Pastor, F.I.; Vidal, T.; Blanca Roncero, M.; Díaz, P.; Martínez, J.; Valenzuela, S.V. Antioxidant activity of xylooligosaccharides produced from glucuronoxylan by Xyn10A and Xyn30D xylanases and eucalyptus autohydrolysates. Carbohyd. Polym. 2018, 194, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Shen, D.; Zhang, L.; Zhong, R.; Liu, Z.; Liu, L.; Chen, L.; Zhang, H. Supplementation of non-starch polysaccharide enzymes cocktail in a corn-miscellaneous meal diet improves nutrient digestibility and reduces carbon dioxide emissions in finishing pigs. Animals 2020, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Petry, A.L.; Patience, J.F. Xylanase supplementation in corn-based swine diets: A review with emphasis on potential mechanisms of action. J. Anim. Sci. 2020, 98, skaa318. [Google Scholar] [CrossRef]
- Kim, H.-J.; Yoo, S.-H. Effects of combined α-amylase and endo-xylanase treatments on the properties of fresh and frozen doughs and final breads. Polymers 2020, 12, 1349. [Google Scholar] [CrossRef]
- Dahiya, S.; Bajaj, B.K.; Kumar, A.; Tiwari, S.K.; Singh, B. A review on biotechnological potential of multifarious enzymes in bread making. Process Biochem. 2020, 99, 290–306. [Google Scholar] [CrossRef]
- Ghoshal, G.; Shivhare, U.S.; Banerjee, U.C. Rheological properties and microstructure of xylanase containing whole wheat bread dough. J. Food Sci. Technol. 2017, 54, 1928–1937. [Google Scholar] [CrossRef]
- Bibra, M.; Kunreddy, V.R.; Sani, R.K. Thermostable xylanase production by Geobacillus sp. strain DUSELR13, and its application in ethanol production with lignocellulosic biomass. Microorganisms 2018, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Dhruw, C.; Husain, K.; Kumar, V.; Sonawane, V.C. Novel xylanase producing Bacillus strain X2: Molecular phylogenetic analysis and its application for production of xylooligosaccharides. 3 Biotech. 2020, 10, 328. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, G.; Chapadgaonkar, S.S. Taguchi optimization and scale up of xylanase from Bacillus licheniformis isolated from hot water geyser. J. Genet. Eng. Biotechnol. 2020, 18, 65. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Cao, R.; Wang, M.; Lin, Q.; Zhan, R.; Xu, H.; Wang, S. A novel thermostable GH10 xylanase with activities on a wide variety of cellulosic substrates from a xylanolytic Bacillus strain exhibiting significant synergy with commercial Celluclast 1.5 L in pretreated corn stover hydrolysis. Biotechnol. Biofuels 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatokun, E.N.; Nwodo, U.U.; Okoh, A.I. Classical optimization of cellulase and xylanase production by a marine Streptomyces Species. Appl. Sci. 2016, 6, 286. [Google Scholar] [CrossRef] [Green Version]
- Maehara, T.; Yagi, H.; Sato, T.; Ohnishi-Kameyama, M.; Fujimoto, Z.; Kamino, K.; Kitamura, Y.; St John, F.; Yaoi, K.; Kaneko, S. GH30 Glucuronoxylan-Specific Xylanase from Streptomyces turgidiscabies C56. Appl. Environ. Microbiol. 2018, 31, 84. [Google Scholar]
- Ouephanit, C.; Boonvitthya, N.; Bozonnet, S.; Chulalaksananukul, W. High-level heterologous expression of endo-1,4-β-xylanase from Penicillium citrinum in Pichia pastoris X-33 directed through codon optimization and optimized expression. Molecules 2019, 24, 3515. [Google Scholar] [CrossRef] [Green Version]
- Duarte, G.C.; Moreira, L.R.S.; Gómez-Mendoza, D.P.; Siqueira, F.G.; Batista, L.R.; Amaral, L.I.V.; Ricart, C.A.O.; Filho, E.X.F. Use of residual biomass from the textile industry as carbon source for production of a low-molecular-weight xylanase from Aspergillus oryzae. Appl. Sci. 2012, 2, 754–772. [Google Scholar] [CrossRef]
- Hüttner, S.; Granchi, Z.; Nguyen, T.T.; Pelt, S.V.; Larsbrink, J.; Thanh, V.N.; Olsson, L. Genome sequence of Rhizomucor pusillus FCH 5.7, a thermophilic zygomycete involved in plant biomass degradation harbouring putative GH9 endoglucanases. Biotechnol. Rep. 2018, 20, e00279. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Kumar, B.; Agarwal, K.; Chaturvedi, V.; Verma, P. Purification and characterization of a thermo-acid/alkali stable xylanases from Aspergillus oryzae LC1 and its application in xylooligosaccharides production from lignocellulosic agricultural wastes. Int. J. Biol. Macromol. 2019, 122, 1191–1202. [Google Scholar] [CrossRef]
- Sukhumsirichart, W.; Deesukon, W.; Kawakami, T.; Matsumoto, S.; Seesom, W.; Sakamoto, T. Expression and characterization of recombinant GH11 xylanase from thermotolerant Streptomyces sp. SWU10. Appl. Biochem. Biotechnol. 2014, 172, 436–446. [Google Scholar] [CrossRef]
- Kim, D.Y.; Han, M.K.; Oh, H.W.; Park, D.S.; Kim, S.J.; Lee, S.G.; Shin, D.H.; Son, K.H.; Bae, K.S.; Park, H.Y. Catalytic properties of a GH10 endo-β-1, 4- xylanase from Streptomyces thermocarboxydus HY-15 isolated from the gut of Eisenia fetida. J. Mol. Catal. B Enzym. 2010, 62, 32–39. [Google Scholar] [CrossRef]
- Chi, W.J.; Lim, J.H.; Park, D.Y.; Park, J.S.; Hong, S.K. Production and characterization of a thermostable endo-type β-xylanase produced by a newly-isolated Streptomyces thermocarboxydus subspecies MW8 strain from Jeju Island. Process Biochem. 2013, 48, 1736–1743. [Google Scholar] [CrossRef]
- Deesukon, W.; Nishimura, Y.; Sakamoto, T.; Sukhumsirichart, W. Purification, characterization of GH11 endo-β-1,4-xylanase from thermotolerant Streptomyces sp. SWU10 and overexpression in Pichia pastoris KM71H. Mol. Biotechnol. 2013, 54, 37–46. [Google Scholar] [CrossRef]
- Yan, Q.; Hao, S.; Jiang, Z.; Zhai, Q.; Chen, W. Properties of a xylanase from Streptomyces matensis being suitable for xylooligosaccharides production. J. Mol. Catal. B Enzym. 2009, 58, 72–77. [Google Scholar] [CrossRef]
- Liu, L.; Xu, M.; Cao, Y.; Wang, H.; Shao, J.; Xu, M.; Zhang, Y.; Wang, Y.; Zhang, W.; Meng, X.; et al. Biochemical characterization of xylanases from Streptomyces sp. B6 and their application in the xylooligosaccharide production from viscose fiber production waste. J. Agric. Food Chem. 2020, 68, 3184–3194. [Google Scholar] [CrossRef]
- Elegir, G.; Szakács, G.; Jeffries, T.W. Purification, characterization, and substrate specificities of multiple xylanases from Streptomyces sp. strain B-12-2. Appl. Environ. Microbiol. 1994, 60, 2609–2615. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.L.; Yen, Y.H.; Shih, I.L.; Chang, A.C.; Chang, W.T.; Wu, W.C.; Chai, Y.D. Production of xylanases from rice bran by Streptomyces actuosus A-151. Enzym. Microb. Technol. 2003, 33, 917–925. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, X.; Sun, B.; Song, H.; Li, E.; Song, H. Properties of an alkaline-tolerant, thermostable xylanase from Streptomyces chartreusis L1105, suitable for xylooligosaccharide production. J. Food Sci. 2012, 2012 77, C506–C511. [Google Scholar] [CrossRef]
- Walia, A.; Guleria, S.; Mehta, P.; Chauhan, A.; Parkash, J. Microbial xylanases and their industrial application in pulp and paper biobleaching: A review. 3 Biotech. 2017, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.H.; Doan, C.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Reclamation of fishery processing waste: A mini-review. Molecules 2019, 24, 2234. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.L.; Nguyen, V.B.; Doan, C.T.; Tran, T.N.; Nguyen, M.T.; Nguyen, A.D. Production and potential applications of bioconversion of chitin and protein-containing fishery byproducts into prodigiosin: A Review. Molecules 2020, 25, 2744. [Google Scholar] [CrossRef] [PubMed]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Vo, T.P.K.; Nguyen, A.D.; Wang, S.L. Chitin extraction from shrimp waste by liquid fermentation using an alkaline protease-producing strain, Brevibacillus parabrevis. Int. J. Biol. Macromol. 2019, 131, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, R.; Hassan, S.S.; Williams, G.A.; Jaiswal, A.K. A Review on Bioconversion of Agro-Industrial Wastes to Industrially Important Enzymes. Bioengineering 2018, 5, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Reclamation of marine chitinous materials for chitosanase production via microbial conversion by Paenibacillus macerans. Mar. Drugs 2018, 16, 429. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.L.; Yu, H.T.; Tsai, M.H.; Doan, C.T.; Nguyen, V.B.; Do, V.C.; Nguyen, A.D. Conversion of squid pens to chitosanases and dye adsorbents via Bacillus cereus. Res. Chem. Intermed. 2018, 44, 4903–4911. [Google Scholar] [CrossRef]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Conversion of squid pens to chitosanases and proteases via Paenibacillus sp. TKU042. Mar. Drugs 2018, 16, 83. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.N.; Doan, C.T.; Nguyen, M.T.; Nguyen, V.B.; Vo, T.P.K.; Nguyen, A.D.; Wang, S.L. An exochitinase with N-acetyl-β-glucosaminidase-like activity from shrimp head conversion by Streptomyces speibonae and its application in hydrolyzing β-chitin powder to produce N-acetyl-D-glucosamine. Polymers 2019, 11, 1600. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, N.; Kumar, B.; Verma, P. A detailed overview of xylanases: An emerging biomolecule for current and future prospective. Bioresour. Bioprocess. 2019, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.N.; Doan, C.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. The isolation of chitinase from Streptomyces thermocarboxydus and its application in the preparation of chitin oligomers. Res. Chem. Intermed. 2019, 45, 727–742. [Google Scholar] [CrossRef]
- Silva, L.A.O.; Fanchini Terrasan, C.R.; Carmona, E.C. Purification and characterization of xylanases from Trichoderma inhamatum. Electron. J. Biotechnol. 2015, 18, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Doan, C.T.; Tran, T.N.; Wen, I.H.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Conversion of shrimp head waste for production of a thermotolerant, detergent-stable, alkaline protease by Paenibacillus sp. Catalysts 2019, 9, 798. [Google Scholar] [CrossRef] [Green Version]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Utilization of seafood processing by-products for production of proteases by Paenibacillus sp. TKU052 and their application in biopeptides’ preparation. Mar. Drugs 2020, 18, 574. [Google Scholar] [CrossRef] [PubMed]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Tran, T.D.; Nguyen, A.D.; Wang, S.L. Bioprocessing of squid pens waste into chitosanase by Paenibacillus sp. TKU047 and its application in low-molecular weight chitosan oligosaccharides production. Polymers 2020, 12, 1163. [Google Scholar] [CrossRef]
- Tran, T.N.; Doan, C.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Anti-oxidant and anti-diabetes potential of water-soluble chitosan–glucose derivatives produced by Maillard reaction. Polymers 2019, 11, 1714. [Google Scholar] [CrossRef] [Green Version]
- Doan, C.T.; Tran, T.N.; Wang, C.L.; Wang, S.L. Microbial conversion of shrimp heads to proteases and chitin as an effective dye adsorbent. Polymers 2020, 12, 2228. [Google Scholar] [CrossRef]
- Meryandini, A. Characterization of xylanase from Streptomyces spp. strain C1-3. HAYATI J. Biosci. 2007, 14, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Ai, Z.; Jiang, Z.; Li, L.; Deng, W.; Kusakabe, I.; Li, H. Immobilization of Streptomyces olivaceoviridis E-86 xylanase on Eudragit S-100 for xylo-oligosaccharide production. Process Biochem. 2005, 40, 2707–2714. [Google Scholar] [CrossRef]
- Georis, J.; Giannotta, F.; De Buyl, E.; Granier, B.; Frère, J. Purification and properties of three endo-beta-1,4-xylanases produced by Streptomyces sp. strain S38 which differ in their ability to enhance the bleaching of kraft pulps. Enzym. Microb. Technol. 2000, 26, 178–186. [Google Scholar] [CrossRef]
- Kumar, A.; Gupta, R.; Shrivastava, B.; Khasa, Y.P.; Kuhad, R.C. Xylanase production from an alkalophilic actinomycete isolate Streptomyces sp. RCK-2010, its characterization and application in saccharification of second generation biomass. J. Mol. Catal. B Enzym. 2012, 74, 170–177. [Google Scholar] [CrossRef]
- Pradeep, G.C.; Choi, Y.H.; Choi, Y.S.; Seong, C.N.; Cho, S.S.; Lee, H.J.; Yoo, J.C. A novel thermostable cellulase free xylanase stable in broad range of pH from Streptomyces sp. CS428. Process Biochem. 2013, 48, 1188–1196. [Google Scholar]
- Ninawe, S.; Kapoor, M.; Kuhad, R.C. Purification and characterization of extracellular xylanase from Streptomyces cyaneus SN32. Bioresour. Technol. 2008, 99, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Sinjaroonsak, S.; Chaiyaso, T.; H-Kittikun, A. Optimization of cellulase and xylanase productions by Streptomyces thermocoprophilus TC13W using low cost pretreated oil palm empty fruit bunch. Waste Biomass Valori. 2019, 11, 3925–3936. [Google Scholar] [CrossRef]
- Wu, H.; Cheng, X.; Zhu, Y.; Zeng, W.; Chen, G.; Liang, Z. Purification and characterization of a cellulase-free, thermostable endo-xylanase from Streptomyces griseorubens LH-3 and its use in biobleaching on eucalyptus kraft pulp. J. Biosci. Bioeng. 2018, 125, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; She, Y.; Sun, B.; Song, H.; Zhu, Y.; Lv, Y.; Song, H. Purification and characterization of a cellulase-free, thermostable xylanase from Streptomyces rameus L2001 and its biobleaching effect on wheat straw pulp. Biochem. Eng. J. 2010, 52, 71–78. [Google Scholar] [CrossRef]
- Chaiyaso, T.; Kuntiya, A.; techapun, C.; Leksawasdi, N.; Seesuriyachan, P.; Hanmoungjai, P. Optimization of cellulase-free xylanase production by thermophilic Streptomyces thermovulgaris TISTR1948 through Plackett-Burman and response surface methodological approaches. Biosci. Biotechnol. Biochem. 2011, 75, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Sanjivkumar, M.; Silambarasan, T.; Balagurunathan, R.; Immanuel, G. Biosynthesis, molecular modeling and statistical optimization of xylanase from a mangrove associated actinobacterium Streptomyces variabilis (MAB3) using Box-Behnken design with its bioconversion efficacy. Int. J. Biol. Macromol. 2018, 118, 195–208. [Google Scholar] [CrossRef]
- Coman, G.; Bahrim, G. Optimization of xylanase production by Streptomyces sp. P12-137 using response surface methodology and central composite design. Ann. Microbiol. 2011, 61, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Boonchuay, P.; Takenaka, S.; Kuntiya, A.; Techapun, C.; Leksawasdi, N.; Seesuriyachan, P.; Chaiyaso, T. Purification, characterization, and molecular cloning of the xylanase from Streptomyces thermovulgaris TISTR1948 and its application to xylooligosaccharide production. J. Mol. Catal. B Enzym. 2016, 129, 61–68. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Production of a thermostable chitosanase from shrimp heads via Paenibacillus mucilaginosus TKU032 conversion and its application in the preparation of bioactive chitosan oligosaccharides. Mar. Drugs 2019, 17, 217. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Gioia, F.; Tzortzakis, N.; Rouphael, Y.; Kyriacou, M.C.; Sampaio, S.L.; Ferreira, I.; Petropoulos, S.A. Grown to be blue—antioxidant properties and health effects of colored vegetables. Part II: Leafy, fruit, and other vegetables. Antioxidants 2020, 9, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowdhaman, D.; Ponnusami, V. Production and optimization of xylooligosaccharides from corncob by Bacillus aerophilus KGJ2 xylanase and its antioxidant potential. Int. J. Biol. Macromol. 2015, 79, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, X.; Liang, C.; Jiang, X.; Yang, G.; Xu, J.; Yong, Q. A sustainable process for procuring biologically active fractions of high-purity xylooligosaccharides and water-soluble lignin from Moso bamboo prehydrolyzate. Biotechnol. Biofuels 2019, 12, 189. [Google Scholar] [CrossRef] [Green Version]
- Rashand, M.M.; Mahmoud, A.E.; Nooman, M.U.; Mahmoud, H.A.; El-Torky, A.E.M.M.; Keshta, A.T. Production of antioxidant xylooligosaccharides from lignocellulosic materials using Bacillus amyloliquifaciens NRRL B-14393 xylanase. J. App. Pharm. Sci. 2016, 6, 030–036. [Google Scholar] [CrossRef] [Green Version]
- Christakopoulos, P.; Katapodis, P.; Kalogeris, E.; Kekos, D.; Macris, B.J.; Stamatis, H.; Skaltsa, H. Antimicrobial activity of acidic xylo-oligosaccharides produced by family 10 and 11 endoxylanases. Int. J. Biol. Macromol. 2003, 31, 171–175. [Google Scholar] [CrossRef]
- Kallel, F.; Driss, D.; Chaabouni, S.E.; Ghorbel, R. Biological Activities of xylooligosaccharides generated from garlic straw xylan by purified xylanase from Bacillus mojavensis UEB-FK. Appl. Biochem. Biotechnol. 2015, 175, 950–964. [Google Scholar] [CrossRef]
- Moura, P.; Barata, R.; Carvalheiro, F.; Gírio, F.; Loureiro-Dias, M.; Esteves, M.P. In vitro fermentation of xylo-oligosaccharides from corn cobs autohydrolysis by Bifidobacterium and Lactobacillus strains. LWT Food Sci. Technol. 2007, 40, 963–972. [Google Scholar] [CrossRef]
- Mäkeläinen, H.; Saarinen, M.; Stowell, J.; Rautonen, N.; Ouwehand, A.C. Xylo-oligosaccharides and lactitol promote the growth of Bifidobacterium lactis and Lactobacillus species in pure cultures. Benef. Microbes. 2010, 1, 139–148. [Google Scholar] [CrossRef]
- Driss, D.; Zouari-Ellouzi, S.; Chaari, F.; Kallel, F.; Ghazala, I.; Bouaziz, F.; Ghorbel, R.; Chaabouni, S. Production and in vitro evaluation of xylooligosaccharides generated from corncobs using immobilized Penicillium occitanis xylanase. J. Mol. Catal. B Enzym. 2014, 102, 146–153. [Google Scholar] [CrossRef]
- Mon, W.P.; Boonlue, S.; Mongkolthanaruk, W. Investigation of Streptomyces for reduction of commercial silk dyes. Korean J. Microbiol. 2020, 56, 206–213. [Google Scholar]
- Velkova, Z.Y.; Kirova, G.K.; Stoytcheva, M.S.; Gochev, V.K. Biosorption of Congo Red and Methylene Blue by pretreated waste Streptomyces fradiae biomass—Equilibrium, kinetic and thermodynamic studies. J. Serb. Chem. Soc. 2018, 83, 107–120. [Google Scholar] [CrossRef]
- Wang, X.S.; Chen, J.P. Biosorption of Congo Red from Aqueous Solution using Wheat Bran and Rice Bran: Batch Studies. Sep. Sci. Technol. 2009, 44, 1452–1466. [Google Scholar] [CrossRef]
- Ciçek, F.; Ozer, D.; Ozer, A.; Ozer, A. Low cost removal of reactive dyes using wheat bran. J. Hazard Mater. 2007, 146, 408–416. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain name | Carbon and Nitrogen Sources | Culture Conditions | MW of Xylanase(s) | Ref. |
|---|---|---|---|---|
| S. thermocarboxydus TKU045 | 2% WBP, and 1.50% KNO3 | 50/250 mL, pH 9.0, 37 °C, 125 rpm, 48 h | >180, 36, 29, and 27 kDa | this study |
| S. thermocarboxydus HY-5 | 43.962 Da | [31] | ||
| S. thermocarboxydus MW8 | 1% soytone, 0.50% birchwood xylan | pH 7.0, 40 °C, 4 d | 52 kDa | [32] |
| Streptomyces sp. SWU10 | 1% ground rice straw, and 0.20% NaNO3 | 1/3 L, 37 °C, 200 rpm, 3 d | 31 and 44 kDa | [33] |
| S. matensis DW67 | 1.50% corncob xylan, 0.40% yeast extract, and 0.80% tryptone | 50/250 mL, 30 °C, 150 rpm, and 5 d | 21.2 kDa | [34] |
| Streptomyces sp. B6 | 0.20% beechwood xylan, and 0.02% peptone | pH 7.2, 5–7 d | 48 and 33 kDa | [35] |
| Streptomyces sp. B-12-2 | 1% oat spelt xylan | 40.5, 36.2, 36.2, 26.4, and 23.8 kDa | [36] | |
| S. actuosus A-151 | 5% rice bran | 45, 30, 26, and 20 kDa | [37] | |
| S. chartreusis L1105 | 2.50% corncob xylan, 0.50% yeast extract, and 1% tryptone | 50/250 mL, pH 6, 40 °C, 140 rpm, and 7 d | 34.2 kDa | [38] |
| Streptomyces sp. RCK-2010 | 2.50% wheat bran, 1.20% (N2 equivalent) beef extract, and 0.20% (N2 equivalent) peptone | pH 8.0, 40 °C, 200 rpm, 48 h | [60] | |
| Streptomyces sp. CS428 | wheat bran | 37 kDa | [61] | |
| Streptomyces spp. SKK1-8 | 1% yeast extract, 10.30% sucrose, and 0.50% birchwood Xylan | 30 °C, 140 rpm, and 10 d | 15.21, 16.8, and 13.8 kDa | [57] |
| S. olivaceoviridis E-86 | 1.50% corncob xylan, and 1.50% Tryptone | 100/500 mL, pH 6.0, 30 °C, 140 rpm, and 5 d | 23 and 47 kDa | [58] |
| S. cyaneus SN32 | 3% wheat bran, and 1% peptone | 250 mL flask, 50 mL, pH 9.0, 42 °C, 200 rpm, 48 h | 20.5 kDa | [62] |
| Streptomyces sp. S38 | 1% oat-spelleds xylan | 24.5, 37.5, and 38 kDa | [59] | |
| Streptomyces sp. P12-137 | 1% wheat bran, 1% KNO3, and 0.50% xylose | 50/250 mL, pH 7.2, 28 °C, and 120 h | [68] | |
| S. thermocoprophilus TC13W | 1% pretreated oil palm empty fruit bunch, and 0.50% yeast extract | pH 6.5, 150 rpm, 40 °C, and 120 h | [63] | |
| S. variabilis MAB3 | 2% birchwood xylan | pH 8.2, 46.5 °C, and 68 h | 50 kDa | [67] |
| S. thermovulgaris TISTR1948 | 2.70% rice straw, and 0.56% yeast extract | 50/250 mL, pH 7.09, 50.01 °C, and 4 d | 46.2 kDa | [66,69] |
| S. rameus L2001 | 2.50% corncob xylan, and 0.50% yeast extract | 50/250 mL, 140 rpm, 40 °C, pH 6, 7 d | 21.1 kDa | [65] |
| S. griseorubens LH-3 | 3% bagasse semi-cellulose, and 3% yeast extract | 50/250 mL, pH 8, 37 °C, 160 rpm, and 96 h | 45.5 kDa | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, T.N.; Doan, C.T.; Wang, S.-L. Conversion of Wheat Bran to Xylanases and Dye Adsorbent by Streptomyces thermocarboxydus. Polymers 2021, 13, 287. https://doi.org/10.3390/polym13020287
Tran TN, Doan CT, Wang S-L. Conversion of Wheat Bran to Xylanases and Dye Adsorbent by Streptomyces thermocarboxydus. Polymers. 2021; 13(2):287. https://doi.org/10.3390/polym13020287
Chicago/Turabian StyleTran, Thi Ngoc, Chien Thang Doan, and San-Lang Wang. 2021. "Conversion of Wheat Bran to Xylanases and Dye Adsorbent by Streptomyces thermocarboxydus" Polymers 13, no. 2: 287. https://doi.org/10.3390/polym13020287
APA StyleTran, T. N., Doan, C. T., & Wang, S. -L. (2021). Conversion of Wheat Bran to Xylanases and Dye Adsorbent by Streptomyces thermocarboxydus. Polymers, 13(2), 287. https://doi.org/10.3390/polym13020287