
Synthetic Biomimetic Polymethacrylates: Promising Platform for the Design of Anti-Cyanobacterial and Anti-Algal Agents

 , ,
, ,  , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Synthesis of Cationic Amphiphilic Poly(Methacrylate) Derivatives (SBPs)
2.2. Cyanobacterial and Algal Cultures
2.3. Growth Inhibition Assay
2.4. Algicidal/Cyanocidal Activity Assay
2.5. Data Analysis and Statistics
3. Results
3.1. Design and Characterization of Studied Biomimetic Polymethacrylates (SBPs)
3.2. Growth-Inhibitory Activity
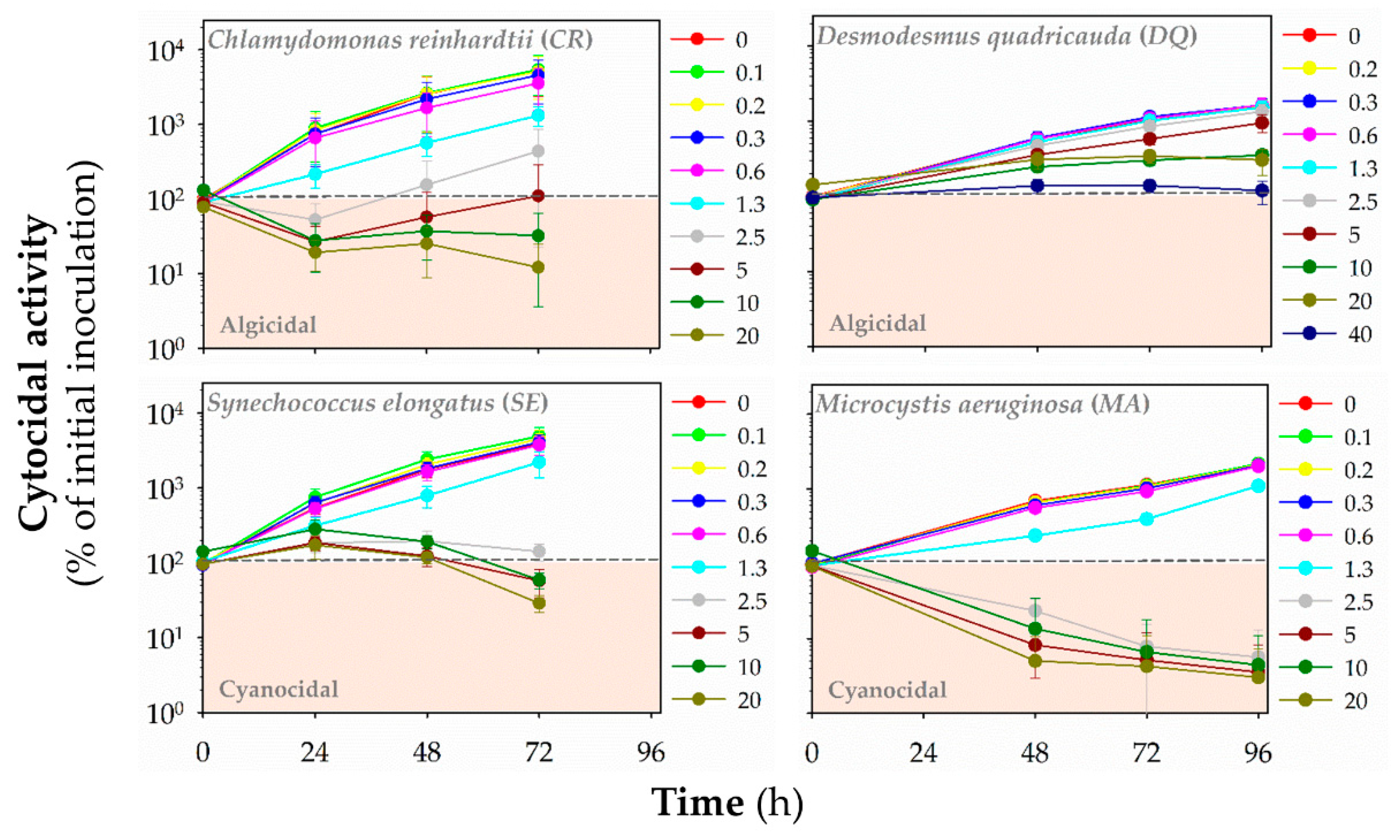
3.3. Algicidal/Cyanocidal Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vardaka, E.; Kormas, K.A. Advancing knowledge on cyanobacterial blooms in freshwaters. Water 2020, 12, 2583. [Google Scholar] [CrossRef]
- World Health Organization. Algae and cyanobacteria in fresh water. In Guidelines for Safe Recreational Water Environments; World Health Organization: Geneva, Switzerland, 2003; pp. 136–154. [Google Scholar]
- Di Pippo, F.; Di Gregorio, L.; Congestri, R.; Tandoi, V.; Rossetti, S. Biofilm growth and control in cooling water industrial systems. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauer, T.; Čapek, P.; Böhmová, P. Main photoautotrophic components of biofilms in natural draft cooling towers. Folia Microbiol. 2016, 61, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, M.H. Removal of cyanobacterial and algal cells from water by ultrasonic waves—A review. J. Mol. Liq. 2016, 222, 1109–1114. [Google Scholar] [CrossRef]
- Sengco, M.R.; Anderson, D.M. Controlling harmful algal blooms through clay flocculation. J. Eukaryot. Microbiol. 2004, 51, 169–172. [Google Scholar] [CrossRef]
- Anderson, D.M. Approaches to monitoring, control and management of harmful algal blooms (HABs). Ocean Coast. Manag. 2009, 52, 342. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, D.P.; Salmaso, N.; Paerl, H.W. Mitigating harmful cyanobacterial blooms: Strategies for control of nitrogen and phosphorus loads. Aquat. Ecol. 2016, 50, 351–366. [Google Scholar] [CrossRef]
- Matthijs, H.C.P.; Jančula, D.; Visser, P.M.; Maršálek, B. Existing and emerging cyanocidal compounds: New perspectives for cyanobacterial bloom mitigation. Aquat. Ecol. 2016, 50, 443–460. [Google Scholar] [CrossRef] [Green Version]
- Westrick, J.A.; Szlag, D.C.; Southwell, B.J.; Sinclair, J. A review of cyanobacteria and cyanotoxins removal/inactivation in drinking water treatment. Anal. Bioanal. Chem. 2010, 397, 1705–1714. [Google Scholar] [CrossRef]
- Jančula, D.; Maršálek, B. Critical review of actually available chemical compounds for prevention and management of cyanobacterial blooms. Chemosphere 2011, 85, 1415–1422. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Xu, Y.; Fusetani, N. Natural products as antifouling compounds: Recent progress and future perspectives. Biofouling 2010, 26, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Ergene, C.; Yasuhara, K.; Palermo, E.F. Biomimetic antimicrobial polymers: Recent advances in molecular design. Polym. Chem. 2018, 9, 2407–2427. [Google Scholar] [CrossRef] [Green Version]
- Kamaruzzaman, N.F.; Tan, L.P.; Hamdan, R.H.; Choong, S.S.; Wong, W.K.; Gibson, A.J.; Chivu, A.; Pina, M.d.F. Antimicrobial polymers: The potential replacement of existing antibiotics? Int. J. Mol. Sci. 2019, 20, 2747. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.R.E.; Fonseca, A.C.; Mendonça, P.V.; Branco, R.; Serra, A.C.; Morais, P.V.; Coelho, J.F.J. Recent developments in antimicrobial polymers: A review. Materials 2016, 9, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Palermo, E.F.; Yasuhara, K.; Caputo, G.A.; Kuroda, K. Molecular design, structures, and activity of antimicrobial peptide-mimetic polymers. Macromol. Biosci. 2013, 13, 1285–1299. [Google Scholar] [CrossRef] [Green Version]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Sovadinova, I.; Palermo, E.F.; Urban, M.; Mpiga, P.; Caputo, G.A.; Kuroda, K. Activity and mechanism of antimicrobial peptide-mimetic amphiphilic polymethacrylate derivatives. Polymers 2011, 3, 1512–1532. [Google Scholar] [CrossRef]
- Sovadinova, I.; Palermo, E.F.; Huang, R.; Thoma, L.M.; Kuroda, K. Mechanism of polymer-induced hemolysis: Nanosized pore formation and osmotic lysis. Biomacromolecules 2011, 12, 260–268. [Google Scholar] [CrossRef]
- Nadres, E.T.; Takahashi, H.; Kuroda, K. Radical-medicated end-group transformation of amphiphilic methacrylate random copolymers for modulation of antimicrobial and hemolytic activities. J. Polym. Sci. Part A Polym. Chem. 2017, 55, 304–312. [Google Scholar] [CrossRef]
- Mikula, P.; Mlnarikova, M.; Takahashi, H.; Babica, P.; Kuroda, K.; Blaha, L.; Sovadinova, I. Branched poly(ethylene imine)s as anti-algal and anti-cyanobacterial agents with selective flocculation behavior to cyanobacteria over algae. Macromol. Biosci. 2018, 18, e1800187. [Google Scholar] [CrossRef]
- Staub, R. Research on physiology of nutrients of the planktonic cyanobacterium Oscillatoria rubescens. Schweiz. Z. Hydrol. 1961, 23, 83–198. [Google Scholar]
- Bold, H.C. The morphology of Chlamydomonas chlamydogama, Sp. Nov. Bull. Torrey Bot. Club 1949, 76, 101–108. [Google Scholar] [CrossRef]
- Pouneva, I. Evaluation of algal culture viability and physiological state by fluorescent microscopic methods. Bulg. J. Plant Physiol. 1997, 23, 67–76. [Google Scholar]
- Schulze, K.; Lopez, D.A.; Tillich, U.M.; Frohme, M. A simple viability analysis for unicellular cyanobacteria using a new autofluorescence assay, automated microscopy, and ImageJ. BMC Biotechnol. 2011, 11, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetsche, E.-M.; Meysman, F.J.R. Dead or alive? Viability assessment of micro- and mesoplankton. J. Plankton Res. 2012, 34, 493. [Google Scholar] [CrossRef]
- Palermo, E.F.; Vemparala, S.; Kuroda, K. Cationic spacer arm design strategy for control of antimicrobial activity and conformation of amphiphilic methacrylate random copolymers. Biomacromolecules 2012, 13, 1632–1641. [Google Scholar] [CrossRef]
- Sabatini, V.; Cattò, C.; Cappelletti, G.; Cappitelli, F.; Antenucci, S.; Farina, H.; Ortenzi, M.A.; Camazzola, S.; Di Silvestro, G. Protective features, durability and biodegration study of acrylic and methacrylic fluorinated polymer coatings for marble protection. Prog. Org. Coat. 2018, 114, 47–57. [Google Scholar] [CrossRef]
- Preece, E.P.; Hardy, F.J.; Moore, B.C.; Bryan, M. A review of microcystin detections in Estuarine and Marine waters: Environmental implications and human health risk. Harmful Algae 2017, 61, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Arora, M.; Sahoo, D. Green Algae BT. In The Algae World; Sahoo, D., Seckbach, J., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 91–120. ISBN 978-94-017-7321-8. [Google Scholar]
- Janssen, C.R.; Vangheluwe, M.; Van Sprang, P. A brief review and critical evaluation of the status of microbiotests BT. In New Microbiotests for Routine Toxicity Screening and Biomonitoring; Persoone, G., Janssen, C., De Coen, W., Eds.; Springer: Boston, MA, USA, 2000; pp. 27–37. ISBN 978-1-4615-4289-6. [Google Scholar]
- Kenawy, E.-R.; Worley, S.D.; Broughton, R. The chemistry and applications of antimicrobial polymers: A state-of-the-art review. Biomacromolecules 2007, 8, 1359–1384. [Google Scholar] [CrossRef]
- Palermo, E.F.; Sovadinova, I.; Kuroda, K. Structural determinants of antimicrobial activity and biocompatibility in membrane-disrupting methacrylamide random copolymers. Biomacromolecules 2009, 10, 3098–3107. [Google Scholar] [CrossRef]
- Taha, S.T.; Han, H.; Chang, S.-R.; Sovadinova, I.; Kuroda, K.; Langford, R.M.; Clarkson, B.H. Nano/micro fluorhydroxyapatite crystal pastes in the treatment of dentin hypersensitivity: An in vitro study. Clin. Oral Investig. 2015, 19, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-C.; Moon, J.C.; Kim, N.-H.; Kim, E.-J.; Jeong, J.-E.; Nelson, A.D.L.; Jo, B.-H.; Jang, M.-K.; Lee, J.R. Algicidal effect of hybrid peptides as potential inhibitors of harmful algal blooms. Biotechnol. Lett. 2016, 38, 847–854. [Google Scholar] [CrossRef]
- Park, S.-C.; Lee, J.-K.; Kim, S.W.; Park, Y. Selective algicidal action of peptides against harmful algal bloom species. PLoS ONE 2011, 6, e26733. [Google Scholar] [CrossRef] [Green Version]
- Schafer, H.; Hettler, H.; Fritsche, U.; Pitzen, G.; Roderer, G.; Wenzel, A. Biotests using unicellular algae and ciliates for predicting long-term effects of toxicants. Ecotoxicol. Environ. Saf. 1994, 27, 64–81. [Google Scholar] [CrossRef] [PubMed]
- US Environmental Protection Agency. OPP Pesticide Ecotoxicity Database; US Environmental Protection Agency: Washington, DC, USA, 2016.
- Wyatt, N.B.; Gloe, L.M.; Brady, P.V.; Hewson, J.C.; Grillet, A.M.; Hankins, M.G.; Pohl, P.I. Critical conditions for ferric chloride-induced flocculation of freshwater algae. Biotechnol. Bioeng. 2012, 109, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, Y.; Tao, Y.; Zhang, Y.; Li, A.; Li, T.; Sang, M.; Zhang, C. Freshwater microalgae harvested via flocculation induced by pH decrease. Biotechnol. Biofuels 2013, 6, 98. [Google Scholar] [CrossRef] [Green Version]
- Hoiczyk, E.; Hansel, A. Cyanobacterial cell walls: News from an unusual prokaryotic envelope. J. Bacteriol. 2000, 182, 1191–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantt, E. Supramolecular membrane organization. In The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006; p. 121. [Google Scholar]
- Haranahalli, K.; Tong, S.; Ojima, I. Recent advances in the discovery and development of antibacterial agents targeting the cell-division protein FtsZ. Bioorg. Med. Chem. 2016, 24, 6354–6369. [Google Scholar] [CrossRef] [Green Version]
- Klibanov, A.M. Permanently microbicidal materials coatings. J. Mater. Chem. 2007, 17, 2479–2482. [Google Scholar] [CrossRef]
- Tashiro, T. Antibacterial and bacterium adsorbing macromolecules. Macromol. Mater. Eng. 2001, 286, 63–87. [Google Scholar] [CrossRef]
- Gonzalo, S.; Rodea-Palomares, I.; Leganes, F.; Garcia-Calvo, E.; Rosal, R.; Fernandez-Pinas, F. First evidences of PAMAM dendrimer internalization in microorganisms of environmental relevance: A linkage with toxicity and oxidative stress. Nanotoxicology 2015, 9, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Petit, A.-N.; Debenest, T.; Eullaffroy, P.; Gagne, F. Effects of a cationic PAMAM dendrimer on photosynthesis and ROS production of Chlamydomonas reinhardtii. Nanotoxicology 2012, 6, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Petit, A.-N.; Eullaffroy, P.; Debenest, T.; Gagne, F. Toxicity of PAMAM dendrimers to Chlamydomonas reinhardtii. Aquat. Toxicol. 2010, 100, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, D. Geometric models for calculating cell biovolume and surface area for phytoplankton. J. Plankton Res. 2003, 25, 1331–1346. [Google Scholar] [CrossRef] [Green Version]
- Umysova, D.; Vitova, M.; Douskova, I.; Bisova, K.; Hlavova, M.; Cizkova, M.; Machat, J.; Doucha, J.; Zachleder, V. Bioaccumulation and toxicity of selenium compounds in the green alga Scenedesmus quadricauda. BMC Plant Biol. 2009, 9, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domozych, D.S.; Ciancia, M.; Fangel, J.U.; Mikkelsen, M.D.; Ulvskov, P.; Willats, W.G.T. The cell walls of green algae: A journey through evolution and diversity. Front. Plant Sci. 2012, 3, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voigt, J.; Woestemeyer, J.; Frank, R. The chaotrope-soluble glycoprotein GP2 is a precursor of the insoluble glycoprotein framework of the Chlamydomonas cell wall. J. Biol. Chem. 2007, 282, 30381–30392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiore, M.F.; Trevors, J.T. Cell composition and metal tolerance in cyanobacteria. Biometals 1994, 7, 83–103. [Google Scholar] [CrossRef]
- Jürgens, U.J.; Martin, C.; Weckesser, J. Cell wall constituents of Microcystis sp. PCC 7806. FEMS Microbiol. Lett. 1989, 65, 47–51. [Google Scholar] [CrossRef]
- Dai, H.; Huang, Y.; Huang, H. Eco-friendly polyvinyl alcohol/carboxymethyl cellulose hydrogels reinforced with graphene oxide and bentonite for enhanced adsorption of methylene blue. Carbohydr. Polym. 2018, 185, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Sharma, K.; Tiwary, K.P.; Sen, G. Polymethacrylic acid grafted psyllium (Psy-g-PMA): A novel material for waste water treatment. Appl. Water Sci. 2013, 3, 285–291. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SBP | End Group | HG 1 (mol. %) | DP 2 1H NMR | Mn3 (g/mol) 1H NMR | Mn4 (g/mol) GPC (boc-protected polymer) | Đ 5 GPC | |
|---|---|---|---|---|---|---|---|
| α | ω | (boc-protected polymer) | |||||
| HP1 | AIBN | Phenyl thio-ester | 0 | 15.2 | 3900 | 2400 | 1.15 |
| HP2 | AIBN | Cyanoisobutyl | 0 | n.d. | n.d. | 2700 | 1.15 |
| HP3 | Cationic | Phenyl thio-ester | 0 | 15.7 | 3600 | 2400 | 1.11 |
| CP1 | AIBN | Phenyl thio-ester | 47.5 | 16.8 | 3500 | 2100 | 1.12 |
| CP2 | AIBN | Phenyl thio-ester | 27.3 | 16.2 | 3900 | 2100 | 1.14 |
| CP3 | AIBN | Phenyl thio-ester | 32.8 | 14.1 | 3400 | 2100 | 1.17 |
| CP4 | Cationic | Phenyl thio-ester | 27.6 | 10.5 | 2900 | 2000 | 1.11 |
| CP5 | Cationic | Cationic | 26.8 | n.d. | n.d. | 2000 | 1.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikula, P.; Mlnaříková, M.; Nadres, E.T.; Takahashi, H.; Babica, P.; Kuroda, K.; Bláha, L.; Sovadinová, I. Synthetic Biomimetic Polymethacrylates: Promising Platform for the Design of Anti-Cyanobacterial and Anti-Algal Agents. Polymers 2021, 13, 1025. https://doi.org/10.3390/polym13071025
Mikula P, Mlnaříková M, Nadres ET, Takahashi H, Babica P, Kuroda K, Bláha L, Sovadinová I. Synthetic Biomimetic Polymethacrylates: Promising Platform for the Design of Anti-Cyanobacterial and Anti-Algal Agents. Polymers. 2021; 13(7):1025. https://doi.org/10.3390/polym13071025
Chicago/Turabian StyleMikula, Přemysl, Marie Mlnaříková, Enrico T. Nadres, Haruko Takahashi, Pavel Babica, Kenichi Kuroda, Luděk Bláha, and Iva Sovadinová. 2021. "Synthetic Biomimetic Polymethacrylates: Promising Platform for the Design of Anti-Cyanobacterial and Anti-Algal Agents" Polymers 13, no. 7: 1025. https://doi.org/10.3390/polym13071025
APA StyleMikula, P., Mlnaříková, M., Nadres, E. T., Takahashi, H., Babica, P., Kuroda, K., Bláha, L., & Sovadinová, I. (2021). Synthetic Biomimetic Polymethacrylates: Promising Platform for the Design of Anti-Cyanobacterial and Anti-Algal Agents. Polymers, 13(7), 1025. https://doi.org/10.3390/polym13071025