
Polyhydroxyalkanoate Production from Fruit and Vegetable Waste Processing



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Agricultural Residues, and Preparation, Treatment, and Analysis
2.2. Culture Media and Experimental Set-Up
2.3. PHA Analysis
2.4. Analytic Methodologies
2.5. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition and Sugar Extraction from Fruit and Vegetable Wastes
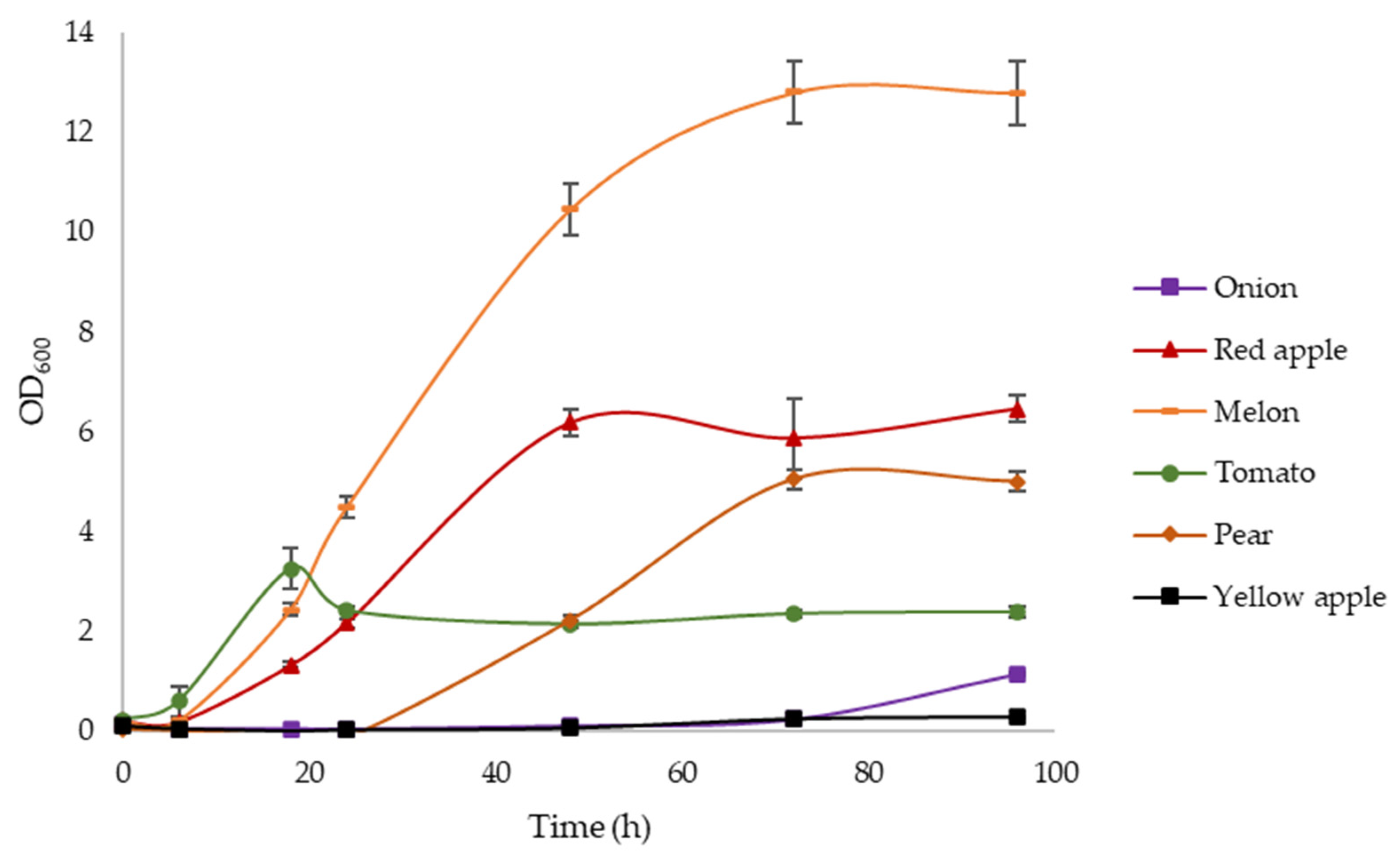
3.2. Initial Screening of C. necator DSM 545 and H. pseudoflava DSM 1034 Growth with Different Agricultural Wastes
3.3. PHA Production from Selected Agricultural Wastes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asmelash, E.; Boshell, F.; Castellanos, G. Reaching Zero with Renewables; International Renewable Energy Agency: Abu Dhabi, United Arab Emirates, 2020; Volume 216, Available online: www.irena.org/publications (accessed on 3 December 2022).
- Circular Economy Network 3° Rapporto Sull’economia Circolare in Italia. 2021. Available online: https://circulareconomynetwork.it/rapporto-2021/ (accessed on 3 December 2022).
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological Degradation of Plastics: A Comprehensive Review. Biotechnol. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.B.S.; Malafaia, C.B. Perspectives on the Production, Structural Characteristics and Potential Applications of Bioplastics Derived from Polyhydroxyalkanoates. Int. J. Biol. Macromol. 2018, 107, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, J.M.; Cardona, C.A.; Higuita, J.C. Use of Residual Banana for Polyhydroxybutyrate (PHB) Production: Case of Study in an Integrated Biorefinery. Waste Manag. 2014, 34, 2634–2640. [Google Scholar] [CrossRef] [PubMed]
- Obruca, S.; Sedlacek, P.; Slaninova, E.; Fritz, I.; Daffert, C.; Meixner, K.; Sedrlova, Z.; Koller, M. Novel Unexpected Functions of PHA Granules. Appl. Microbiol. Biotechnol. 2020, 104, 4795–4810. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Perez, S.; Serrano, A.; Pantión, A.A.; Alonso-Fariñas, B. Challenges of Scaling-up PHA Production from Waste Streams. A Review. J. Environ. Manage. 2018, 205, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Yadav, B.; Pandey, A.; Kumar, L.R.; Tyagi, R.D. Bioconversion of Waste (Water)/Residues to Bioplastics—A Circular Bioeconomy Approach. Bioresour. Technol. 2020, 298, 122584. [Google Scholar] [CrossRef]
- Favaro, L.; Basaglia, M.; Casella, S. Improving Polyhydroxyalkanoate Production from Inexpensive Carbon Sources by Genetic Approaches: A Review. Biofuels, Bioprod. Biorefining 2019, 13, 208–227. [Google Scholar] [CrossRef] [Green Version]
- Samorì, C.; Basaglia, M.; Casella, S.; Favaro, L.; Galletti, P.; Giorgini, L.; Marchi, D.; Mazzocchetti, L.; Torri, C.; Tagliavini, E. Dimethyl Carbonate and Switchable Anionic Surfactants: Two Effective Tools for the Extraction of Polyhydroxyalkanoates from Microbial Biomass. Green Chem. 2015, 17, 1047–1056. [Google Scholar] [CrossRef]
- Jõgi, K.; Bhat, R. Valorization of Food Processing Wastes and By-Products for Bioplastic Production. Sustain. Chem. Pharm. 2020, 18, 100326. [Google Scholar] [CrossRef]
- European Bioplastics; Nova-Institute Bioplastics. Market Development Update 2020. Eur. Bioplastics Org. 2020, 2021, 2020–2021. [Google Scholar]
- Koller, M.; Bona, R.; Braunegg, G.; Hermann, C.; Horvat, P.; Kroutil, M.; Martinz, J.; Neto, J.; Pereira, L.; Varila, P. Articles from ISBP 2004. Time 2008, 6, 561–565. [Google Scholar]
- Esparza, I.; Jiménez-Moreno, N.; Bimbela, F.; Ancín-Azpilicueta, C.; Gandía, L.M. Fruit and Vegetable Waste Management: Conventional and Emerging Approaches. J. Environ. Manage. 2020, 265, 110510. [Google Scholar] [CrossRef] [PubMed]
- Santagata, R.; Ripa, M.; Genovese, A.; Ulgiati, S. Food Waste Recovery Pathways: Challenges and Opportunities for an Emerging Bio-Based Circular Economy. A Systematic Review and an Assessment. J. Clean. Prod. 2021, 286, 125490. [Google Scholar] [CrossRef]
- Bomfim, A.S.C.D.; Oliveira, D.M.D.; Voorwald, H.J.C.; Benini, K.C.C.D.C.; Dumont, M.J.; Rodrigue, D. Valorization of Spent Coffee Grounds as Precursors for Biopolymers and Composite Production. Polymers 2022, 14, 437. [Google Scholar] [CrossRef]
- Ravindran, R.; Jaiswal, A.K. Exploitation of food industry waste for high-value products. Trends Biotechnol. 2016, 34, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Correa-Galeote, D.; Argiz, L.; Val del Rio, A.; Mosquera-Corral, A.; Juarez-Jimenez, B.; Gonzalez-Lopez, J.; Rodelas, B. Dynamics of PHA-Accumulating Bacterial Communities Fed with Lipid-Rich Liquid Effluents from Fish-Canning Industries. Polymers 2022, 14, 1396. [Google Scholar] [CrossRef]
- Marciniak, P.; Możejko-Ciesielska, J. What is new in the field of industrial wastes conversion into polyhydroxyalkanoates by bacteria? Polymers 2021, 13, 1731. [Google Scholar] [CrossRef]
- Gupte, A.P.; Basaglia, M.; Casella, S.; Favaro, L. Rice waste streams as a promising source of biofuels: Feedstocks, biotechnologies and future perspectives. Renew. Sust. Energy Rev. 2022, 167, 112673. [Google Scholar] [CrossRef]
- Iglesias-Iglesias, R.; Portela-Grandío, A.; Treu, L.; Campanaro, S.; Kennes, C.; Veiga, M.C. Co-digestion of cheese whey with sewage sludge for caproic acid production: Role of microbiome and polyhydroxyalkanoates potential production. Biores. Technol. 2021, 337, 125388. [Google Scholar] [CrossRef]
- Abbondanzi, F.; Biscaro, G.; Carvalho, G.; Favaro, L.; Lemos, P.; Paglione, M.; Torri, C. Fast method for the determination of short-chain-length polyhydroxyalkanoates (scl-PHAs) in bacterial samples by In Vial-Thermolysis (IVT). New Biotechnol. 2017, 39, 29–35. [Google Scholar] [CrossRef]
- Alibardi, L.; Green, K.; Favaro, L.; Vale, P.; Soares, A.; Cartmell, E.; Fernández, Y.B. Performance and stability of sewage sludge digestion under CO2 enrichment: A pilot study. Biores. Technol. 2017, 245, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakeney, M. Global Food Losses and Food Waste. 2011. Available online: http://www.fao.org/docrep/014/mb060e/mb060e00.pdf (accessed on 3 December 2022).
- Dornburg, V.; Lewandowski, I.; Patel, M. Comparing the Land Requirements, Energy Savings, and Greenhouse Gas Emissions Reduction of Biobased Polymers and Bioenergy: An Analysis and System Extension of Life-Cycle Assessment Studies. J. Ind. Ecol. 2003, 7, 93–116. [Google Scholar] [CrossRef]
- Blakeney, M. Food loss and waste and food security. In Food Loss Food Waste; Chapter 1; Edward Elgar Publishing: Cheltenham, UK, 2019; Volume 1, pp. 1–26. [Google Scholar] [CrossRef] [Green Version]
- The Food Loss and Waste Accounting and Reporting Standard. 2016. Available online: https://www.wbcsd.org/Programs/Food-and-Nature/Food-Land-Use/Climate-Smart-Agriculture/Resources/Food-Loss-and-Waste-Accounting-and-Reporting-Standard%0Afile:///C (accessed on 3 December 2022).
- Koller, M.; Maršálek, L.; de Sousa Dias, M.M.; Braunegg, G. Producing Microbial Polyhydroxyalkanoate (PHA) Biopolyesters in a Sustainable Manner. N. Biotechnol. 2017, 37, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, K.; Dumont, M.J.; Del Rio, L.F.; Orsat, V. Producing PHAs in the Bioeconomy—Towards a Sustainable Bioplastic. Sustain. Prod. Consum. 2017, 9, 58–70. [Google Scholar] [CrossRef]
- Tsang, Y.F.; Kumar, V.; Samadar, P.; Yang, Y.; Lee, J.; Ok, Y.S.; Song, H.; Kim, K.H.; Kwon, E.E.; Jeon, Y.J. Production of Bioplastic through Food Waste Valorization. Environ. Int. 2019, 127, 625–644. [Google Scholar] [CrossRef]
- Anagnostopoulou, C.; Kontogiannopoulos, K.N.; Gaspari, M.; Morlino, M.S.; Assimopoulou, A.N.; Kougias, P.G. Valorization of household food wastes to lactic acid production: A response surface methodology approach to optimize fermentation process. Chemosphere 2022, 296, 133871. [Google Scholar] [CrossRef] [PubMed]
- Haas, C.; Steinwandter, V.; De Apodaca, E.D.; Madurga, B.M.; Smerilli, M.; Dietrich, T.; Neureiter, M. Production of PHB from Chicory Roots—Comparison of Three Cupriavidus necator Strains. Chem. Biochem. Eng. Q. 2015, 29, 99–112. [Google Scholar] [CrossRef]
- Haas, R.; Jin, B.; Zepf, F.T. Production of Poly(3-Hydroxybutyrate) from Waste Potato Starch. Biosci. Biotechnol. Biochem. 2008, 72, 253–256. [Google Scholar] [CrossRef]
- Brojanigo, S.; Parro, E.; Cazzorla, T.; Favaro, L.; Basaglia, M.; Casella, S. Conversion of Starchy Waste Streams into Polyhydroxyalkanoates Using Cupriavidus necator DSM 545. Polymers 2020, 12, 1496. [Google Scholar] [CrossRef] [PubMed]
- Brojanigo, S.; Gronchi, N.; Cazzorla, T.; Wong, T.S.; Basaglia, M.; Favaro, L.; Casella, S. Engineering Cupriavidus necator DSM 545 for the One-Step Conversion of Starchy Waste into Polyhydroxyalkanoates. Bioresour. Technol. 2022, 347, 126383. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, M.G.; Povolo, S.; Favaro, L.; Fontana, F.; Basaglia, M.; Casella, S. Engineering Delftia acidovorans DSM39 to Produce Polyhydroxyalkanoates from Slaughterhouse Waste. Int. J. Biol. Macromol. 2014, 71, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Favaro, L.; Basaglia, M.; Rodriguez, J.E.G.; Morelli, A.; Ibraheem, O.; Pizzocchero, V.; Casella, S. Bacterial production of PHAs from lipid-rich by-products. App. Food Biotechnol. 2019, 6, 45–52. [Google Scholar]
- Obruca, S.; Marova, I.; Snajdar, O.; Mravcova, L.; Svoboda, Z. Production of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) by Cupriavidus necator from Waste Rapeseed Oil Using Propanol as a Precursor of 3-Hydroxyvalerate. Biotechnol. Lett. 2010, 32, 1925–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Follonier, S.; Goyder, M.S.; Silvestri, A.C.; Crelier, S.; Kalman, F.; Riesen, R.; Zinn, M. Fruit Pomace and Waste Frying Oil as Sustainable Resources for the Bioproduction of Medium-Chain-Length Polyhydroxyalkanoates. Int. J. Biol. Macromol. 2014, 71, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Basaglia, M.; D’ambra, M.; Piubello, G.; Zanconato, V.; Favaro, L.; Casella, S. Agro-food Residues and Bioethanol Potential: A Study for a Specific Area. Processes 2021, 9, 344. [Google Scholar] [CrossRef]
- Rodríguez Gamero, J.E.; Favaro, L.; Pizzocchero, V.; Lomolino, G.; Basaglia, M.; Casella, S. Nuclease Expression in Efficient Polyhydroxyalkanoates-Producing Bacteria Could Yield Cost Reduction during Downstream Processing. Bioresour. Technol. 2018, 261, 176–181. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Torri, C.; Cordiani, H.; Samorì, C.; Favaro, L.; Fabbri, D. Fast procedure for the analysis of poly(hydroxyalkanoates) in bacterial cells by online pyrolysis/gas-Chromatography with flame ionization detector. J. Chromatogr. A 2014, 1359, 230–236. [Google Scholar] [CrossRef]
- Braunegg, G.; Sonnleitner, B.; Lafferty, R.M. A Rapid Gas Chromatographic Method for the Determination of Poly-β-Hydroxybutyric Acid in Microbial Biomass. Eur. J. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- Gronchi, N.; Favaro, L.; Cagnin, L.; Brojanigo, S.; Pizzocchero, V.; Basaglia, M.; Casella, S. Novel yeast strains for the efficient saccharification and fermentation of starchy by-products to bioethanol. Energies 2019, 12, 714. [Google Scholar] [CrossRef] [Green Version]
- Kirk, P.L. Kjeldahl Method for Total Nitrogen. Anal. Chem. 1950, 22, 354–358. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On Tyrosine and Tryptophane Determinations in Proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar] [CrossRef]
- Persic, M.; Mikulic-Petkovsek, M.; Slatnar, A.; Veberic, R. Chemical Composition of Apple Fruit, Juice and Pomace and the Correlation between Phenolic Content, Enzymatic Activity and Browning. LWT Food Sci. Technol. 2017, 82, 23–31. [Google Scholar] [CrossRef]
- Ali, S.; Masud, T.; Abbasi, K.S. Physico-Chemical Characteristics of Apricot (Prunus Armeniaca L.) Grown in Northern Areas of Pakistan. Sci. Hortic. 2011, 130, 386–392. [Google Scholar] [CrossRef]
- Saidani, F.; Giménez, R.; Aubert, C.; Chalot, G.; Betrán, J.A.; Gogorcena, Y. Phenolic, Sugar and Acid Profiles and the Antioxidant Composition in the Peel and Pulp of Peach Fruits. J. Food Compos. Anal. 2017, 62, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.A.; Ntatsi, G.; Fernandes; Barros, L.; Barreira, J.C.M.; Ferreira, I.C.F.R.; Antoniadis, V. Long-Term Storage Effect on Chemical Composition, Nutritional Value and Quality of Greek Onion Landrace “Vatikiotiko”. Food Chem. 2016, 201, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Rosa-Martínez, E.; García-Martínez, M.D.; Adalid-Martínez, A.M.; Pereira-Dias, L.; Casanova, C.; Soler, E.; Figàs, M.R.; Raigón, M.D.; Plazas, M.; Soler, S.; et al. Fruit Composition Profile of Pepper, Tomato and Eggplant Varieties Grown under Uniform Conditions. Food Res. Int. 2021, 147, 110531. [Google Scholar] [CrossRef]
- Wilson, E.A.; Demmig-Adams, B. Antioxidant, Anti-Inflammatory, and Antimicrobial Properties of Garlic and Onions. Nutr. Food Sci. 2007, 37, 178–183. [Google Scholar] [CrossRef]
- Turco, R.; Santagata, G.; Corrado, I.; Pezzella, C.; Di Serio, M. In Vivo and Post-Synthesis Strategies to Enhance the Properties of PHB-Based Materials: A Review. Front. Bioeng. Biotechnol. 2021, 8, 1–31. [Google Scholar] [CrossRef]
- Amini, M.; Yousefi-Massumabad, H.; Younesi, H.; Abyar, H.; Bahramifar, N. Production of the Polyhydroxyalkanoate Biopolymer by Cupriavidus necator Using Beer Brewery Wastewater Containing Maltose as a Primary Carbon Source. J. Environ. Chem. Eng. 2020, 8, 103588. [Google Scholar] [CrossRef]
- Low, T.J.; Mohammad, S.; Sudesh, K.; Baidurah, S. Utilization of Banana (Musa Sp.) Fronds Extract as an Alternative Carbon Source for Poly(3-Hydroxybutyrate) Production by Cupriavidus necator H16. Biocatal. Agric. Biotechnol. 2021, 34, 102048. [Google Scholar] [CrossRef]
- Sukruansuwan, V.; Napathorn, S.C. Use of Agro-Industrial Residue from the Canned Pineapple Industry for Polyhydroxybutyrate Production by Cupriavidus necator Strain A-04. Biotechnol. Biofuels 2018, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, N.; Sivakumar, N. Production of Polyhydroxybutyrate from Wheat Bran Hydrolysate Using Ralstonia eutropha through Microbial Fermentation. J. Biotechnol. 2016, 237, 13–17. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| (% Dry Mass) | ||||||
|---|---|---|---|---|---|---|
| Composition | Dry Mass (%) | Cellulose | Hemicellulose | Lignin | Starch | Protein |
| Cucumber | 5.70 | 16.30 | 5.60 | 1.80 | 0.30 | 18.20 |
| Nectarine | 20.80 | 17.60 | 9.20 | 12.60 | - | 7.20 |
| Onion | 11.10 | 6.20 | 2.50 | 0.40 | 0.30 | 10.90 |
| Plum | 14.50 | 15.70 | 7.30 | 10.20 | 0.00 | 6.40 |
| Melon | 6.70 | 18.30 | 3.50 | 5.60 | - | 14.80 |
| Apricot pomace * | 17.0 | 16.00 | 7.10 | 10.10 | - | 7.20 |
| Yellow apple * | 11.10 | 6.00 | 3.10 | 3.80 | - | 2.90 |
| Pear * | 30.30 | 30.20 | 25.60 | 20.40 | - | 3.10 |
| Tomato * | 21.80 | 16.40 | 10.50 | 31.00 | 0.70 | 16.70 |
| Red apple * | 12.60 | 5.40 | 2.90 | 2.80 | - | 2.90 |
| Waste | Glucose | Fructose | Cellobiose | Total Sugars | Total Nitrogen | Total Phosphorus | Total Polyphenols |
|---|---|---|---|---|---|---|---|
| Cucumber | 1.63 | 6.26 | nd | 7.89 | 0.45 | 0.20 | 1.07 |
| Nectarine | 5.63 | 6.48 | nd | 12.11 | 0.77 | 0.31 | 0.40 |
| Onion | 12.35 | 10.72 | nd | 23.07 | 0.15 | 0.06 | 0.97 |
| Plum | 11.86 | 8.24 | 0.41 | 20.51 | 0.58 | 0.16 | 0.57 |
| Melon | 3.06 | 7.14 | nd | 10.20 | 0.09 | 0.03 | 0.45 |
| Apricot pomace * | 5.30 | 3.49 | nd | 8.79 | 0.15 | 0.07 | 0.45 |
| Yellow apple * | 10.29 | 19.40 | 1.52 | 31.21 | 0.17 | 0.18 | 0.43 |
| Pear * | 0.17 | 0.66 | nd | 0.83 | 0.20 | 0.06 | 0.17 |
| Tomato * | 0.67 | 0.93 | nd | 1.60 | 0.01 | 0.02 | 0.20 |
| Red apple * | 10.80 | 22.14 | 3.60 | 36.54 | 0.07 | 0.02 | 0.29 |
| Sterilization | Culture Medium | CDM (g/L) | 3HB (% CDM) | 3HB (g/L) | 3HV (% CDM) | 3HV (g/L) | |
|---|---|---|---|---|---|---|---|
| DSMZ-81 | 0.7 ± 0.22 | 0.3 ± 0.6 | - | - | - | ||
| Tomato | Filtration | DSMZ-81 (no NH4Cl) | 1.3 ± 0.03 | 34.6 ± 2.5 | 0.4 ± 0.1 | - | - |
| Water | 0.1 ± 0.00 | - | - | - | - | ||
| DSMZ-81 | 1.2 ± 0.02 | 1.3 ± 0.7 | 0 | - | - | ||
| Pear | Filtration | DSMZ-81 (no NH4Cl) | 0.8 ± 0.04 | 54.5 ± 11.3 | 0.4 ± 0.2 | - | - |
| Water | 0.1 ± 0.01 | - | - | - | - | ||
| DMSZ-81 | 10.9 ± 0.1 | 67.9 ± 0.6 | 7.4 ± 0.01 | - | - | ||
| Red apple | Filtration | DSMZ-81 (no NH4Cl) | 6.2 ± 0.2 | 79.1 ± 0.9 | 4.9 ± 0.03 | - | - |
| Water | 4.9 ± 0.2 | 79.7 ± 0.1 | 3.9 ± 0.04 | - | - | ||
| DMSZ-81 | 6.5 ± 0.5 | 21.6 ± 1.0 | 1.4 ± 0.1 | - | - | ||
| Red apple | Autoclaving | DSMZ-81 (no NH4Cl) | 6.7 ± 0.04 | 34.5 ± 2.1 | 2.3 ± 0.1 | - | - |
| Water | 5.9 ± 0.5 | 30.8 ± 2.4 | 1.8 ± 0.1 | - | - | ||
| DMSZ-81 | 9.6 ± 0.5 | 20.3 ± 2.3 | 1.94 ± 0.1 | - | - | ||
| Melon | Filtration | DSMZ-81 (no NH4Cl) | 4.2 ± 0.4 | 34.0 ± 6.7 | 1.43 ± 0.2 | - | - |
| Water | 0.3 ± 0.1 | - | - | - | - | ||
| DMSZ-81 | 5.9 ± 0.5 | 32.6 ± 6.1 | 1.93 ± 0.2 | 3.1 ± 0.2 | 0.18 ± 0.1 | ||
| Melon | Autoclaving | DSMZ-81 (no NH4Cl) | 5.8 ± 0.4 | 73.8 ± 10.1 | 4.26 ± 0.1 | 0.7 ± 1.1 | 0.04 ± 1.7 |
| Water | 5.1 ± 0.3 | 74. 9 ± 6.6 | 3.84 ± 0.1 | 4.6 ± 0.3 | 0.23 ± 0.1 |
| Food Waste | Microorganism | Biomass (g/L) | PHB Content (%) | 3HV (%) | Reference |
|---|---|---|---|---|---|
| Treated brewery wastewater | C. necator DSM 545 | 7.9 | 26.5 | 11.5 | [55] |
| Banana frond extract | C. necator H16 | 3.6 | 37.4 | - | [56] |
| Crude aqueous extract of pineapple waste | C. necator A-04 | 13.6 | 60.1 | - | [57] |
| Wheat bran hydrolysate | C. necator H16 (mutant NCIMB 11599) | 24.43 | 62.5 | - | [58] |
| Melon extract | C. necator DSM 545 | 5.1 | 74.9 | 4.6 | This study |
| Red apple extract | C. necator DSM 545 | 10.9 | 67.9 | - | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, P.; Basaglia, M.; Casella, S.; Favaro, L. Polyhydroxyalkanoate Production from Fruit and Vegetable Waste Processing. Polymers 2022, 14, 5529. https://doi.org/10.3390/polym14245529
Costa P, Basaglia M, Casella S, Favaro L. Polyhydroxyalkanoate Production from Fruit and Vegetable Waste Processing. Polymers. 2022; 14(24):5529. https://doi.org/10.3390/polym14245529
Chicago/Turabian StyleCosta, Paolo, Marina Basaglia, Sergio Casella, and Lorenzo Favaro. 2022. "Polyhydroxyalkanoate Production from Fruit and Vegetable Waste Processing" Polymers 14, no. 24: 5529. https://doi.org/10.3390/polym14245529
APA StyleCosta, P., Basaglia, M., Casella, S., & Favaro, L. (2022). Polyhydroxyalkanoate Production from Fruit and Vegetable Waste Processing. Polymers, 14(24), 5529. https://doi.org/10.3390/polym14245529