

Main Characteristics of Processed Grain Starch Products and Physicochemical Features of the Starches from Maize (Zea mays L.) with Different Genotypes

Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Methods
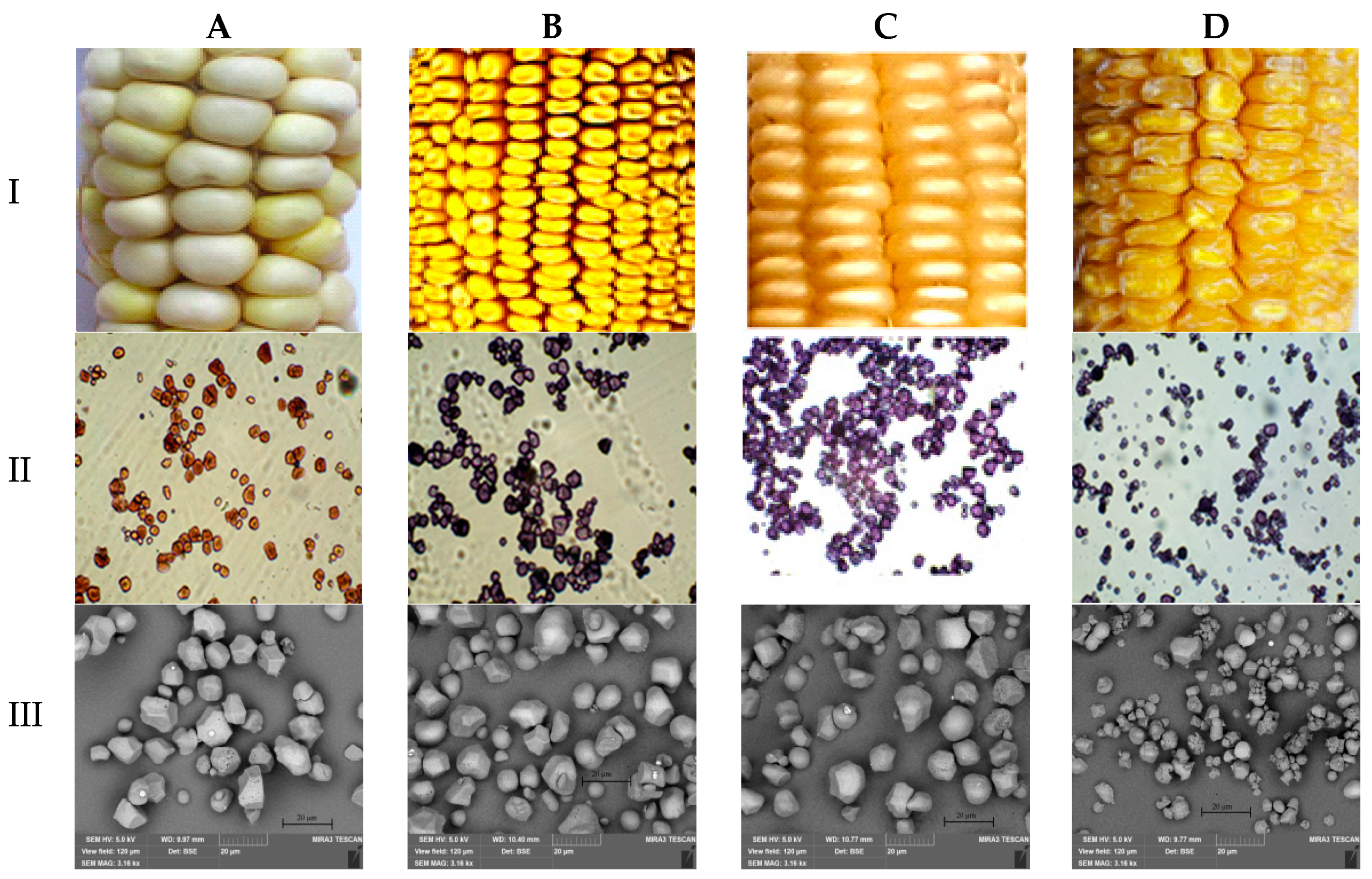
2.2.1. Cytological Analysis
2.2.2. Extraction of Starch and By-Products
2.2.3. Assessment of Grain Characteristics
2.2.4. Measuring the Amylose Content
2.2.5. Light Microscopy
2.2.6. Scanning Electron Microscopy (SEM)
2.2.7. X-ray Diffraction
2.2.8. Differential Scanning Calorimetry
2.2.9. Statistical Analysis
3. Results and Discussion
3.1. Ranking Maize Accessions According to the Type of Starch and Maize Grain Characteristics
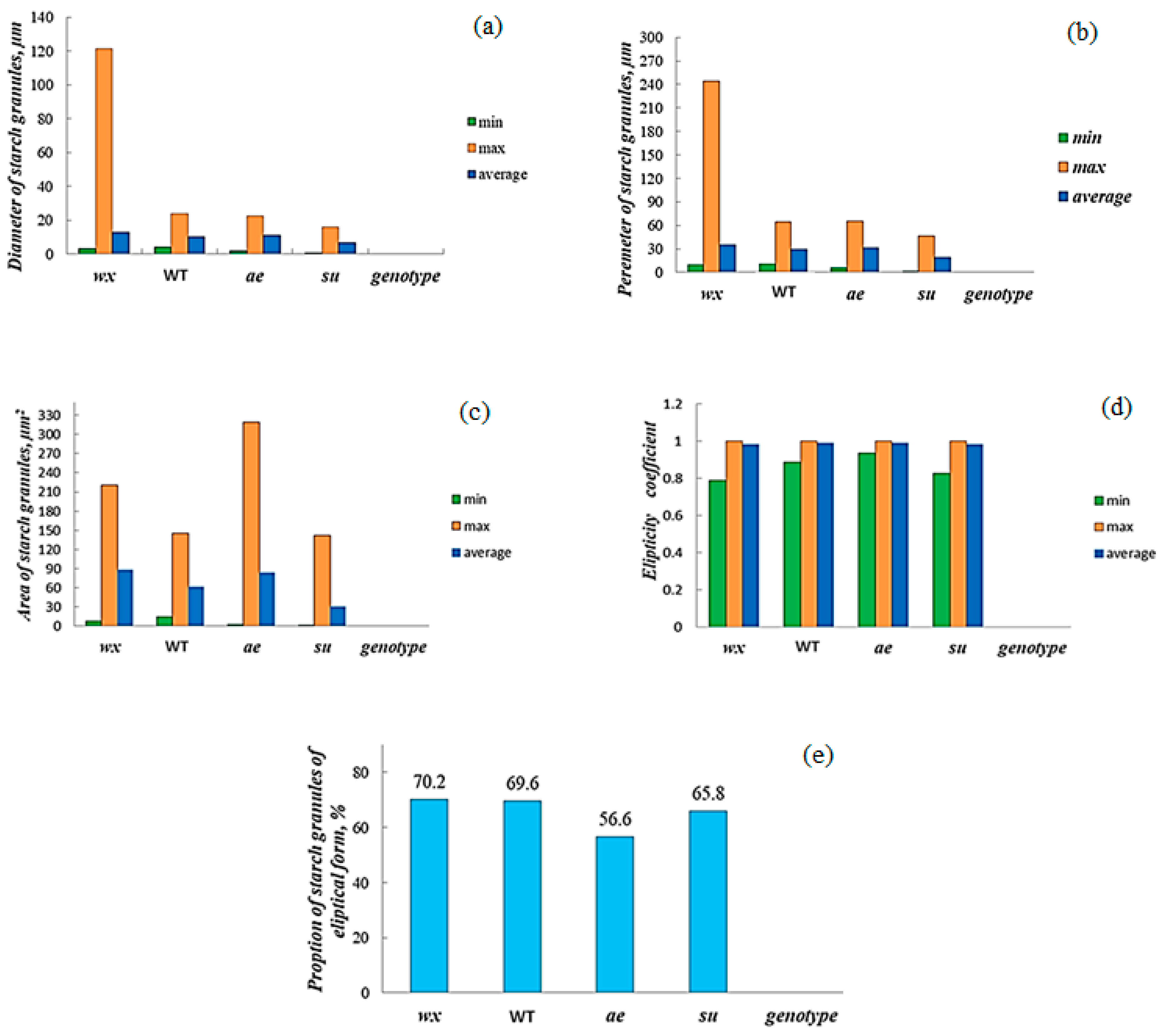
3.2. Analysis and Characteristics of Starch Granule Size
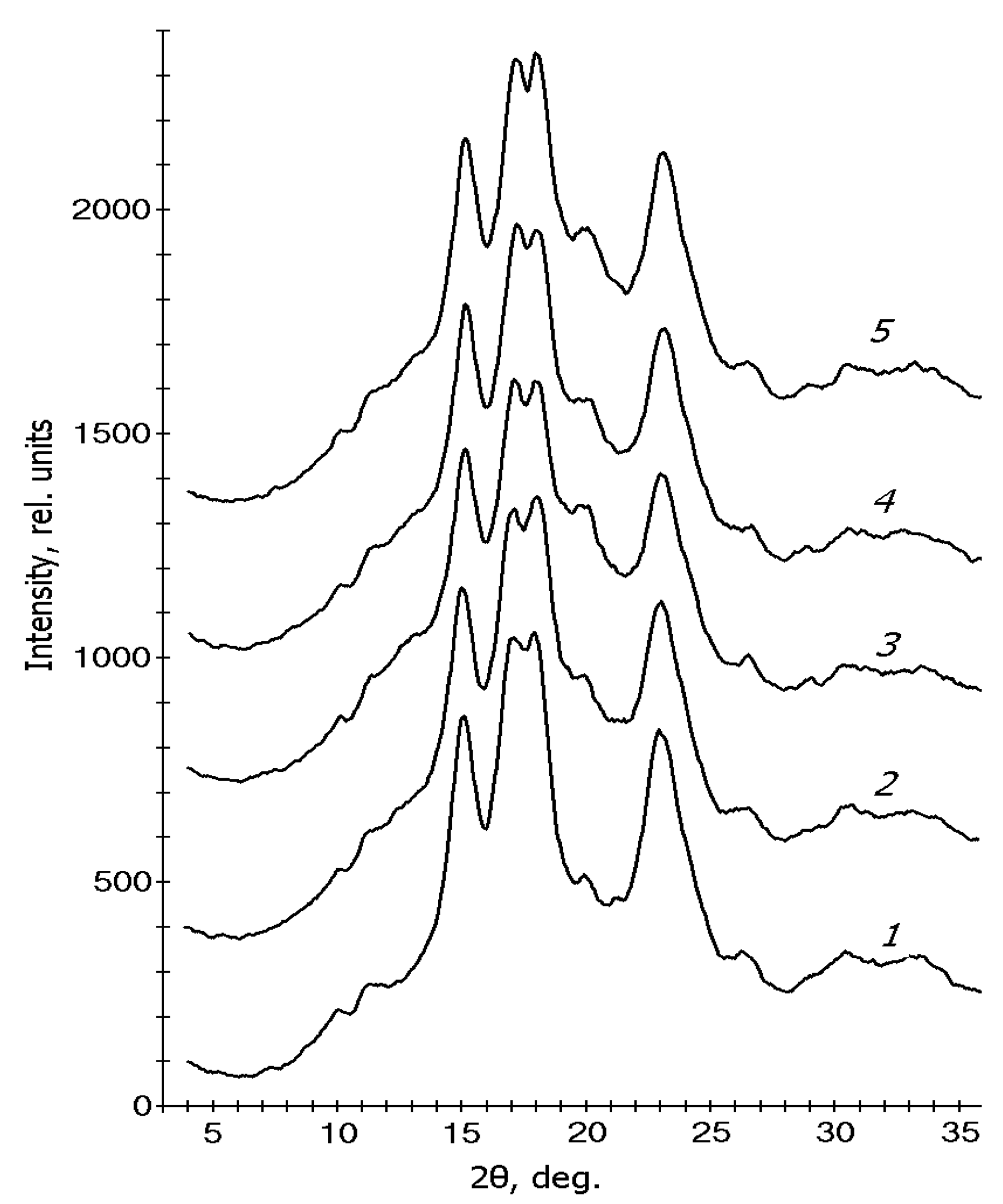
3.3. X-ray Diffraction
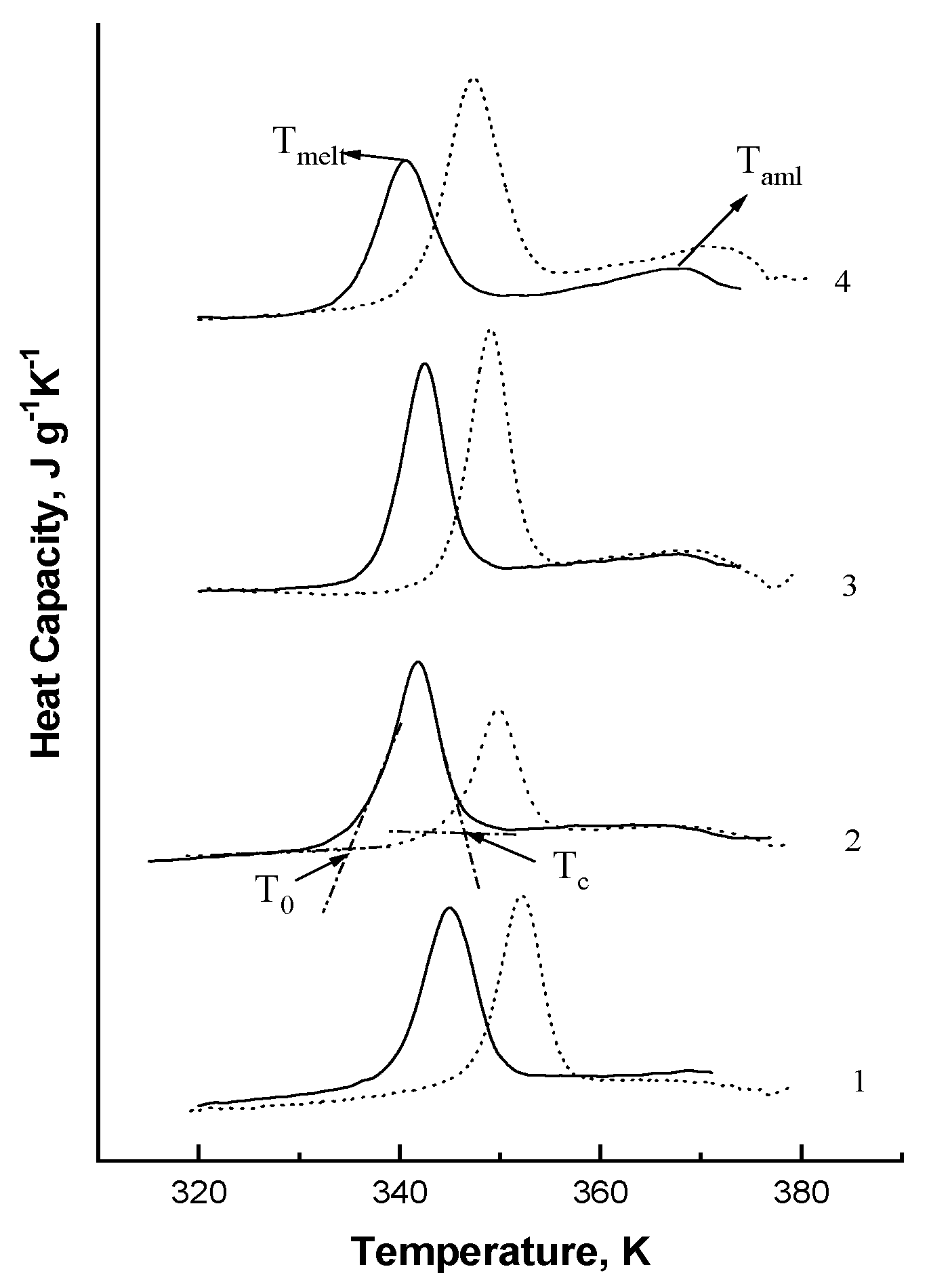
3.4. Thermal Properties of Starches in Different Maize Genotypes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santana, A.L.; Meireles, M.A.A. New starches are the trend for industry applications: A review. Food Public Health 2014, 4, 229–241. [Google Scholar] [CrossRef]
- BeMiller, J.N.; Whistler, R.L. Starch: Chemistry and Technology, 3rd ed.; Academic Press: New York, NY, USA, 2009. [Google Scholar]
- Durrani, C.M.; Donald, A.M. Physical characterization of amylopectin gels. Polym. Gels Netw. 1995, 3, 1–27. [Google Scholar] [CrossRef]
- Zhang, P.; Whistler, R.L.; BeMiller, J.N.; Hamaker, B.R. Banana starch: Production, physicochemical properties, and digestibility—A review. Carbohydr. Polym. 2005, 59, 443–458. [Google Scholar] [CrossRef]
- Hoover, R. Composition, molecular structure, and physicochemical properties of tuber and root starches: A review. Carbohydr. Polym. 2001, 45, 253–267. [Google Scholar] [CrossRef]
- Singh, N.; Singh, J.; Kaur, L.; Sodhi, N.S.; Gill, B.S. Morphological, thermal and rheological properties of starches from different botanical sources. Food Chem. 2003, 81, 219–231. [Google Scholar] [CrossRef]
- Jasim, A.; Brijesh, K.T.; Syed, H.I.; Rao, M.A. (Eds.) Starch-Based Polymeric Materials and Nanocomposites: Chemistry, Processing, and Applications; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar] [CrossRef]
- Guo, B.; Hu, X.; Deng, F.; Wu, J.; Luo, S.; Chen, R.; Liu, C. Supernatant starch fraction of corn starch and its emulsifying ability: Effect of the amylose content. Food Hydrocoll. 2020, 103, 105711. [Google Scholar] [CrossRef]
- Bajaj, R.; Singh, N.; Kaur, A.; Inouchi, N. Structural, morphological, functional and digestibility properties of starches from cereals, tubers and legumes: A comparative study. J. Food Sci. Technol. 2018, 55, 3799–3808. [Google Scholar] [CrossRef]
- Li, H.; Gidley, M.J.; Dhital, S. High-amylose starches to bridge the “Fiber Gap”: Development, structure, and nutritional functionality. Compr. Rev. Food Sci. Food Saf. 2019, 18, 362–379. [Google Scholar] [CrossRef]
- Singh, N.; Inouchi, N.; Nishinari, K. Structural, thermal and viscoelastic characteristics of starches separated from normal, sugary and waxy maize. Food Hydrocoll. 2006, 20, 923–935. [Google Scholar] [CrossRef]
- Li, L.; Jiang, H.; Campbell, M.; Blanco, M.; Jane, J.-L. Characterization of maize amylose-extender (ae) mutant starches. Part I: Relationship between resistant starch contents and molecular structures. Carbohydr. Polym. 2008, 74, 396–404. [Google Scholar] [CrossRef]
- Zhang, R.; Belwal, T.; Li, L.; Lin, X.; Xu, Y.; Luo, Z. Recent advances in polysaccharides stabilized emulsions for encapsulation and delivery of bioactive food ingredients: A review. Carbohydr. Polym. 2020, 242, 116388. [Google Scholar] [CrossRef]
- Xie, F.; Pollet, E.; Halley, P.J.; Av’erous, L. Starch-based nano-biocomposites. Prog. Polym. Sci. 2012, 38, 1590–1628. [Google Scholar] [CrossRef]
- Faisal, M.; Kou, T.; Zhong, Y.; Blennow, A. High amylose-based bio composites: Structures, functions and applications. Polymers 2022, 14, 1235. [Google Scholar] [CrossRef] [PubMed]
- Hung, P.V.; Maeda, T.; Morita, N. Study on physicochemical characteristics of waxy and high-amylose wheat starches in comparison with normal wheat starch. Starch 2007, 59, 125–131. [Google Scholar] [CrossRef]
- Christiansen, C.; Baumann, M.J.; Abu Hachem, M.; Friis, E.; Viksø-Nielsen, A.; Blennow, A.; Svensson, B. Characterization of CBM20 Starch Binding Domains from Three Different Origins. In Proceedings of the Abstract from Plant Biotecj Annual Meeting, København, Denmark, 3 April–3 May 2010. [Google Scholar]
- Liu, J.; Mushegian, A. Three monophyletic superfamilies account for the majority of the known glycosyltransferases. Protein Sci. 2003, 12, 1418–1431. [Google Scholar] [CrossRef] [PubMed]
- Leterrier, M.; Holappa, L.D.; Broglie, K.E.; Beckles, D.M. Cloning, characterisation and comparative analysis of a starch synthase IV gene in wheat: Functional and evolutionary implications. BMC Plant Biol. 2008, 8, 98. [Google Scholar] [CrossRef]
- Shure, M.; Wessler, S.; Fedoroff, N. Molecular identification and isolation of the Waxy locus in maize. Cell 1983, 35, 225–233. [Google Scholar] [CrossRef]
- Craig, J.; Lloyd, J.R.; Tomlinson, K.; Barber, L.; Edwards, A.; Wang, T.L.; Martin, C.; Hedley, C.L.; Smith, A.M. Mutations in the gene encoding starch synthase II profoundly alter amylopectin structure in pea Embryos. Plant Cell 1998, 10, 413–426. [Google Scholar] [CrossRef]
- Mizuno, K.; Kimura, K.; Arai, Y.; Kawasaki, T.; Shimada, H.; Baba, T. Starch branching enzymes from immature rice seeds. J. Biochem. 1992, 112, 643–651. [Google Scholar] [CrossRef]
- Schwall, G.P.; Safford, R.; Westcott, R.J.; Jeffcoat, R.; Tayal, A.; Shi, Y.C.; Gidley, M.J.; Jobling, S.A. Production of very-high-amylose potato starch by inhibition of SBE A and B. Nat. Biotechnol. 2000, 18, 551–554. [Google Scholar] [CrossRef]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef]
- Sidebottom, C.; Kirkland, M.; Strongitharm, B.; Jeffcoat, R. Characterization of the difference of starch branching enzyme activities in normal and low-amylopectin maize during kernel development. J. Cereal Sci. 1998, 27, 279–287. [Google Scholar] [CrossRef]
- Boyer, C.D.; Damewood, P.A.; Matters, G.L. Effect of gene dosage at high amylose loci on the properties of the amylopectin fractions of the starches. Starch 1980, 32, 217–222. [Google Scholar] [CrossRef]
- Takeda, C.; Takeda, Y.; Hizukuri, S. Structure of the amylopectin fraction of amylomaize. Carbohydr. Res. 1993, 246, 273–281. [Google Scholar] [CrossRef]
- James, M.G.; Robertson, D.S.; Myers, A.M. Characterization of the maize gene sugary1, a determinant of starch composition in kernels. Plant Cell 1995, 7, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Myers, A.M.; Morell, M.K.; James, M.G.; Ball, S.G. Recent progress toward understanding biosintesis of the amylopectin crystal. Plant Physiol. 2000, 122, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Dong, S.T.; Gao, R.Q.; Li, Y.Q. Comparison of starch synthesis and related enzyme activities in developing grains among different types of maize. J. Plant Physiol. Mol. Biol. 2007, 33, 25–32. [Google Scholar]
- Khatefov, E.B.; Khoreva, V.I.; Kerv, Y.A.; Shelenga, T.V.; Sidorova, V.V.; Demurin, Y.N.; Goldstein, V.G. Comparative analysis of the chemical composition and size of starch granules in grain between diploid and tetraploid sweetcorn cultivars. Proc. Appl. Bot. Genet. Breed. 2021, 182, 53–62. [Google Scholar] [CrossRef]
- Romeis, B. Mikroskopische Technik (17. Neubearbeitete und Erweiterte Auflage, Herausgegeben von P. BÖCK); Urban und Schwarzenberg: Munich, Germany, 1989. [Google Scholar] [CrossRef]
- Adkins, G.K.; Greenwood, C.T. The isolation of cereal starches in the laboratory. Starch 1966, 7, 213–218. [Google Scholar] [CrossRef]
- McGrance, C.J.; Cornell, H.J.; Rix, C.J. A simple and rapid colorimetric method for the determination of amylose in starch products. Starch 1998, 50, 158–163. [Google Scholar] [CrossRef]
- Lille, M.; Autio, K. Microstructure of high-pressure vs. atmospheric frozen starch gels. Innov. Food Sci. Emerg. Technol. 2007, 8, 117–126. [Google Scholar] [CrossRef]
- Wasserman, L.A.; Papakhin, A.A.; Borodina, Z.M.; Krivandin, A.V.; Sergeev, A.I.; Tarasov, V.F. Some physico-chemical and thermodynamic characteristics of maize starches hydrolyzed by glucoamylase. Carbohydr. Polym. 2019, 212, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, L.A.; Papakhin, A.A.; Krivandin, A.V.; Filatova, A.G.; Borodina, Z.M.; Plashchina, I.G. Changes in the structure, thermodynamic, and functional properties of maize starch during mechanical processing. Russ. J. Phys. Chem. B 2022, 16, 141–147. [Google Scholar] [CrossRef]
- Andreev, N.R.; Kalistratova, E.N.; Wasserman, L.A.; Yuryev, V.P. The influence of heating rate and annealing on the melting thermodynamic parameters of some cereal starches in excess water. Starch 1999, 50, 422–429. [Google Scholar] [CrossRef]
- Bocharnikova, I.; Wasserman, L.A.; Krivandin, A.V.; Fornal, J.; Baszczak, W.; Chernykh, V.Y.; Schiraldi, A.; Yuryev, V.P. Structure and thermodynamic melting parameters of wheat starches with different amylose content. J. Therm. Anal. Calorim. 2003, 74, 681–695. [Google Scholar] [CrossRef]
- Privalov, P.L.; Potekhin, S.A. Scanning microcalorimetry in studying temperature-induced changes in proteins. In Methods in Enzymology; Colowick, N.P., Kaplan, N.P.C.H., Hirs, W., Timasheff, S.N., Eds.; Volume 131, Enzyme Structure. Part I; Elsevier: Amsterdam, The Netherlands, 1986; pp. 4–51. [Google Scholar] [CrossRef]
- Matveev, Y.I.; van Soest, J.J.G.; Nieman, C.; Wasserman, L.A.; Protserov, V.A.; Ezernitskaja, M.; Yuryev, V.P. The relationship between thermodynamic and structural properties of low and high amylose maize starches. Carbohydr. Polym. 2001, 44, 151–160. [Google Scholar] [CrossRef]
- Imberty, A.; Chanzy, H.; Perez, S.; Buleon, A.; Tran, V. The double-helical nature of the crystalline part of A-starch. J. Mol. Biol. 1988, 201, 365–378. [Google Scholar] [CrossRef]
- Singh, J.; Singh, N. Studies on the morphological and rheological properties of granular cold-water soluble corn and potato starches. Food Hydrocoll. 2003, 17, 63–72. [Google Scholar] [CrossRef]
- Bershtein, V.A.; Egorov, V.M. Differential scanning calorimetry of polymers—Physics, chemistry, analysis, technology. In Ellis Horwood Series in Polymer Science and Technology; Kemp, T.J., Ed.; Ellis Horwood Ltd.: New York, NY, USA; London, UK; Toronto, ON, Canada; Sydney, Australia; Tokyo, Japan; Singapore, 1994. [Google Scholar]
- Pérez, S.; Bertoft, E. The molecular structures of starch components and their contribution to the architecture of starch granules: A comprehensive review. Starch 2010, 62, 389–420. [Google Scholar] [CrossRef]
- Peng, X.; Yao, Y. Small-granule starches from sweet corn and cow cockle: Physical properties and amylopectin branching pattaern. Food Hydrocoll. 2018, 74, 349–357. [Google Scholar] [CrossRef]
- Chen, P.; Yua, L.; Chen, L.; Li, X. Morphology and microstructure of maize starches with different amylose/amylopectin content. Starch 2006, 58, 611–615. [Google Scholar] [CrossRef]
- Buléon, A.; Colonna, P.; Planchot, V.; Ball, S. Starch granules: Structure and biosynthesis. Int. J. Biol. Macromol. 1998, 23, 85–112. [Google Scholar] [CrossRef]
- Bogracheva, T.Y.; Morris, V.J.; Ring, S.G.; Hedley, C.L. The granular structure of C-type starch and its role in gelatinisation. Biopolym. 1998, 45, 323–332. [Google Scholar] [CrossRef]
- Biliaderis, C.G. Structures and phase transitions in food systems. Food Technol. 1992, 46, 98–109. [Google Scholar]
- Jenkins, P.J.; Donald, A.M. The influence of amylose on starch granule structure. Int. J. Biol. Macromol. 1995, 17, 315–321. [Google Scholar] [CrossRef]
- Kozlov, S.S.; Krivandin, A.V.; Shatalova, O.V.; Noda, T.; Bertoft, E.; Fornal, J.; Yuryev, V.P. Structure of starches extracted from near-isogenic wheat lines. Part II. Molecular organization of amylopectin clusters. J. Therm. Anal. Calorim. 2007, 87, 575–584. [Google Scholar] [CrossRef]
- Koroteeva, D.A.; Kiseleva, V.I.; Krivandin, A.V.; Shatalova, O.V.; Błaszczak, W.; Bertoft, E.; Piyachomkwan, K.; Yuryev, V.P. Structural and thermodynamic properties of rice starches with different genetic background. Part 2. Defectiveness of different supramolecular structures in starch granules. Int. J. Biol. Macromol. 2007, 41, 534–547. [Google Scholar] [CrossRef]
- Zobel, H.F. Starch crystal transformations and their industrial importance. Starch 1988, 40, 1–7. [Google Scholar] [CrossRef]
- Wunderlich, B. Macromolecular Physics, Volume 2, Crystal Nucleation, Growth, Annealing; Academic Press: New York, NY, USA, 1977. [Google Scholar] [CrossRef]
- Whittam, M.A.; Noel, T.R.; Ring, S.G. Melting behaviour of A- and B-type starches. Int. J. Biol. Macromol. 1990, 12, 359–362. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Ploidy, n | Genotype | Dry Matter Mass Fraction in Grain, % | Starch Content in Grain (S1), % | Mass Fraction of Ash Content in Grain, % | Amylose Content in Starch, % | Starch Yield after Maize Grain Processing under Laboratory Conditions (S2), % of Grain DM | Starch Extraction Ratio (S2/S1·100), % |
|---|---|---|---|---|---|---|---|---|
| Mais agestano | 2 | wx | 92.2 ± 0.3 | 70.5 ± 0.1 | 1.7 ± 0.2 | 0.0 ± 0.5 | 58.6 ± 0.7 | 83.1 |
| Mestnaya | 2 | wx | 92.1 ± 0.4 | 70.8 ± 0.5 | 1.7 ±0.2 | 0.0 ± 0.5 | 59.9 ± 1.4 | 84.6 |
| Populyatsiya MRPP22 | 4 | WT | 91.9 ± 0.4 | 73.7 ± 0.3 | 1.7 ± 0.1 | 15.0 ± 0.7 | 62.9 ± 0.9 | 85.3 |
| Luch | 2 | WT | 92.1 ± 0.2 | 75.3 ± 0.4 | 1.3 ± 0.1 | 17.5 ± 1.0 | 64.7 ± 0.7 | 85.9 |
| Kabardinskaya belaya zubovidnaya | 2 | WT | 91.4 ± 0.3 | 73.0 ± 0.1 | 1.5 ± 0.1 | 21.0 ± 0.9 | 59.9 ± 0.6 | 82.1 |
| Otbornyy 150SV | 2 | WT | 92.1 ± 0.2 | 70.9 ± 0.1 | 1.4 ± 0.1 | 26.4 ± 0.9 | 56.8 ± 0.4 | 80.1 |
| Gornaya chechenskaya | 2 | “ae” | 92.0 ± 0.3 | 71.3 ± 0.3 | 1.5 ±0.2 | 32.0 ± 0.9 | 62.0 ± 0.8 | 86.9 |
| White Flint | 2 | “ae” | 91.2 ± 0.5 | 69.3 ± 0.3 | 1.5 ± 0.1 | 38. 0 ± 0.1 | 62.6 ± 0.7 | 90.3 |
| Baksanskaya sakharnaya | 4 | su | 93.3 ± 0.3 | 59.3 ± 0.2 | 1.7 ± 0.1 | 25.5 ± 1.1 | 20.8 ± 1.4 | 35.1 |
| Alina | 2 | su | 92.8 ± 0.4 | 65.9 ± 0.6 | 1.4 ±0.2 | 26.4 ± 1.2 | 10.5 ± 2.3 | 15.9 |
| Ranyaya Lacomka | 2 | su | 93.3 ± 0.2 | 58.8 ± 0.4 | 1.5 ±0.1 | 27.7± 0.8 | 17.8 ± 2.1 | 30.3 |
| Cultivar | Genotype | Amylose Content, % | C, % |
|---|---|---|---|
| Mais agestano | wx | 0 | 41 |
| Luch | WT | 17.5 | 29 |
| Gornaya chechenskaya | “ae” | 32.0 | 30 |
| White Flint | “ae” | 38.0 | 30 |
| Baksanskaya sakharnaya | su | 25.5 | 26 |
| Cultivar | Genotype/ Ploidy, n | Tm, K | ΔT = Tc ࢤ T0 | ΔHm, kJ/mol | ΔHvH, kJ/mol | ν, Anhydroglucose Residue | Lcrl, nm | Taml, K | ΔHaml, kJ/mol |
|---|---|---|---|---|---|---|---|---|---|
| Mais agestano | wx/2 | 345.8 ± 0.1 | 9.7 | 3.7 ± 0.2 | 48.6 ± 1.5 | 13.4 ± 0.7 | 4.7 ± 0.2 | - | - |
| Mestnaya | wx/2 | 345.2 ± 0.1 | 10.0 | 2.7 ± 0.2 | 38.1 ± 1.5 | 14.2 ± 0.6 | 5.0 ± 0.2 | - | - |
| Populyatsiya MRPP22 | WT/4 | 342.1 ± 0.1 | 10.7 | 2.4 ± 0.1 | 38.2 ± 0.1 | 16.0 ± 0.5 | 5.6 ± 0.2 | 366.5 ± 0.1 | 0.2 ± 0.05 |
| Luch | WT/2 | 343.7 ± 0.1 | 8.4 | 1.9 ± 0.1 | 34.9 ± 1.8 | 17.9 ± 0.1 | 6.3 ± 0.0 | 365.6 ± 0.0 | 0.30 ± 0.05 |
| Kabardinskaya belaya zubovidnaya | WT/2 | 343.1 ± 0.0 | 8.6 | 1.9 ± 0.3 | 36.2 ± 4.0 | 20.0 ± 0.8 | 7.0 ± 0.3 | 365.7 ± 0.1 | 0.10 ± 0.0 |
| Otbornyy 150SV | WT/2 | 343.2 ± 0.1 | 7.3 | 2.1 ± 0.3 | 42.5 ± 2.4 | 21.0 ± 3.0 | 7.4 ± 0.7 | 366.1 ± 0.0 | 0.30 ± 0.05 |
| Gornaya chechenskaya | “ae”/2 | 343.5 ± 0.0 | 10.2 | 2.8 ± 0.3 | 43.2 ± 2.5 | 15.5 ± 0.8 | 5.5 ± 0.2 | 366.0 ± 0.0 | 0.30 ± 0.03 |
| White Flint | “ae”/2 | 342.9 ± 0.0 | 10.4 | 2.7 ± 0.1 | 42.1 ± 1.2 | 16.0 ± 0.4 | 5.6 ± 0.1 | 365.8 ± 0.0 | 0.40 ± 0.0 |
| Baksanskaya sakharnaya | su/4 | 340.8 ± 0.1 | 12.3 | 2.4 ± 0.1 | 35.1 ± 0.4 | 14.7 ± 0.8 | 5.2 ± 0.2 | 367.3 ± 0.0 | 0.7 ± 0.1 |
| Alina | su/2 | 342.1 ± 0.1 | 12.1 | 1.6 ± 0.1 | 27.9 ± 1.2 | 16.9 ± 0.4 | 6.0 ± 0.1 | 367.5 ± 0.1 | 0.3 ± 0.05 |
| Ranyaya Lacomka | su/2 | 341.8 ± 0.1 | 14.5 | 1.8 ± 0.1 | 28.5 ± 0.8 | 15.8 ± 0.4 | 5.5 ± 0.2 | 366.6 ± 0.1 | 0.5 ± 0.1 |
| Mean value | 365.9 ± 0.7 | 0.4 ± 0.1 |
| Cultivar | Genotype/ Ploidy, n | si × 107 J/cm2 |
|---|---|---|
| Mais agestano | wx/2 | 0.08 |
| Mestnaya | wx/2 | 0.13 |
| Populyatsiya MRPP22 | WT/4 | 0.17 |
| Luch | WT/2 | 0.23 |
| Kabardinskaya belaya zubovidnaya | WT/2 | 0.26 |
| Otbornyy 150SV | WT/2 | 0.26 |
| Gornaya chechenskaya | “ae”/2 | 0.15 |
| White Flint | “ae”/2 | 0.16 |
| Baksanskaya sakharnaya | su/4 | 0.16 |
| Alina | su/2 | 0.23 |
| Ranyaya Lacomka | su/2 | 0.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatefov, E.B.; Goldstein, V.G.; Krivandin, A.V.; Wasserman, L.A. Main Characteristics of Processed Grain Starch Products and Physicochemical Features of the Starches from Maize (Zea mays L.) with Different Genotypes. Polymers 2023, 15, 1976. https://doi.org/10.3390/polym15081976
Khatefov EB, Goldstein VG, Krivandin AV, Wasserman LA. Main Characteristics of Processed Grain Starch Products and Physicochemical Features of the Starches from Maize (Zea mays L.) with Different Genotypes. Polymers. 2023; 15(8):1976. https://doi.org/10.3390/polym15081976
Chicago/Turabian StyleKhatefov, Eduard B., Vladimir G. Goldstein, Alexey V. Krivandin, and Lyubov A. Wasserman. 2023. "Main Characteristics of Processed Grain Starch Products and Physicochemical Features of the Starches from Maize (Zea mays L.) with Different Genotypes" Polymers 15, no. 8: 1976. https://doi.org/10.3390/polym15081976
APA StyleKhatefov, E. B., Goldstein, V. G., Krivandin, A. V., & Wasserman, L. A. (2023). Main Characteristics of Processed Grain Starch Products and Physicochemical Features of the Starches from Maize (Zea mays L.) with Different Genotypes. Polymers, 15(8), 1976. https://doi.org/10.3390/polym15081976