
A Novel Heterogalactan from Antrodia camphorata and Anti-Angiogenic Activity of Its Sulfated Derivative

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. General Methods
2.3. Isolation and Purification of Polysaccharide from A. Camphorata
2.4. Purity and Molecular Weight Determination
2.5. Monosaccharide Composition Analysis
2.6. Glycosyl Linkage Analysis
2.7. NMR Analysis
2.8. Preparation of Sulfated Derivatives
2.9. Cell Lines and Culture Conditions
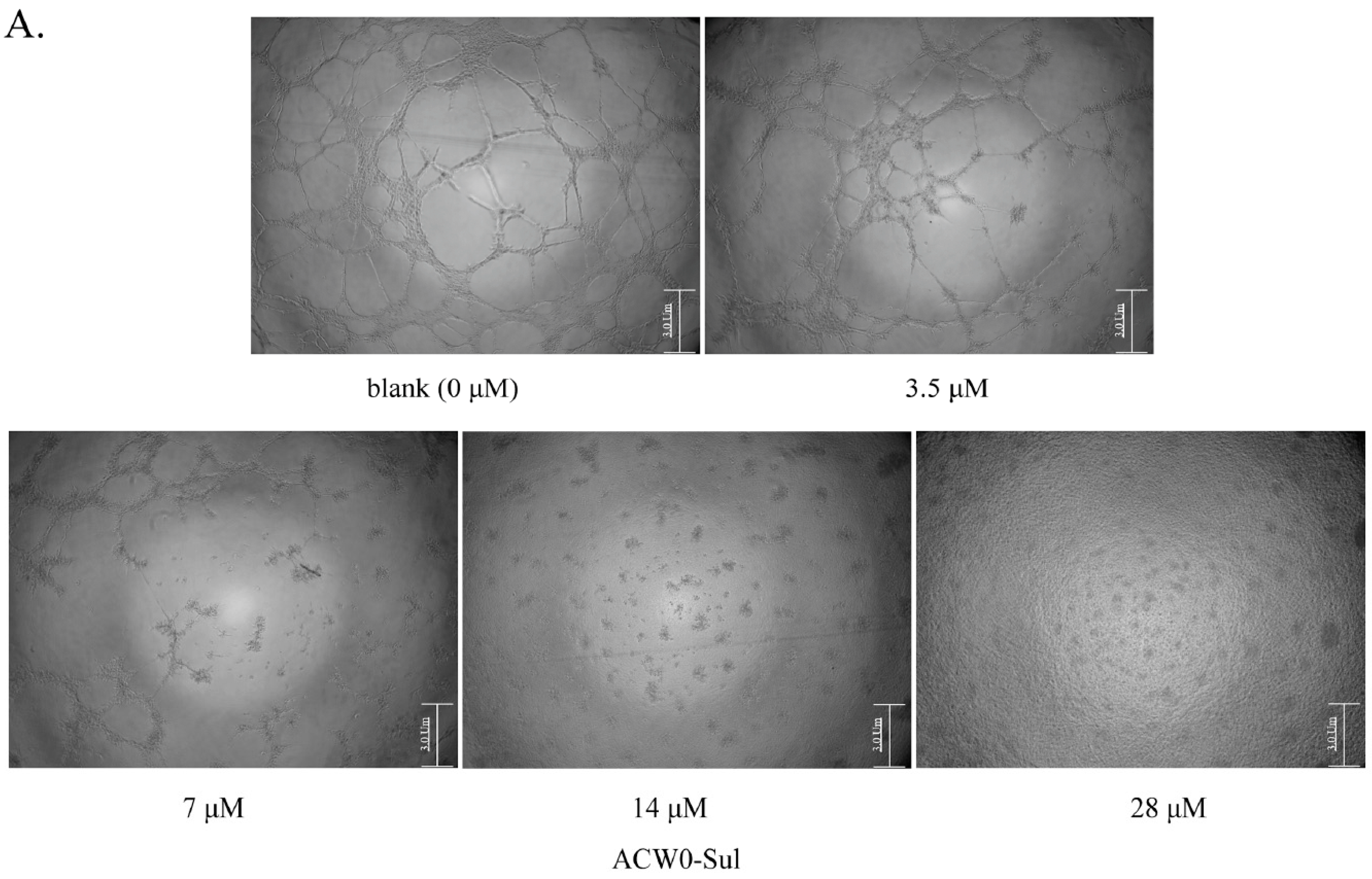
2.10. Tube Formation on Matrigel
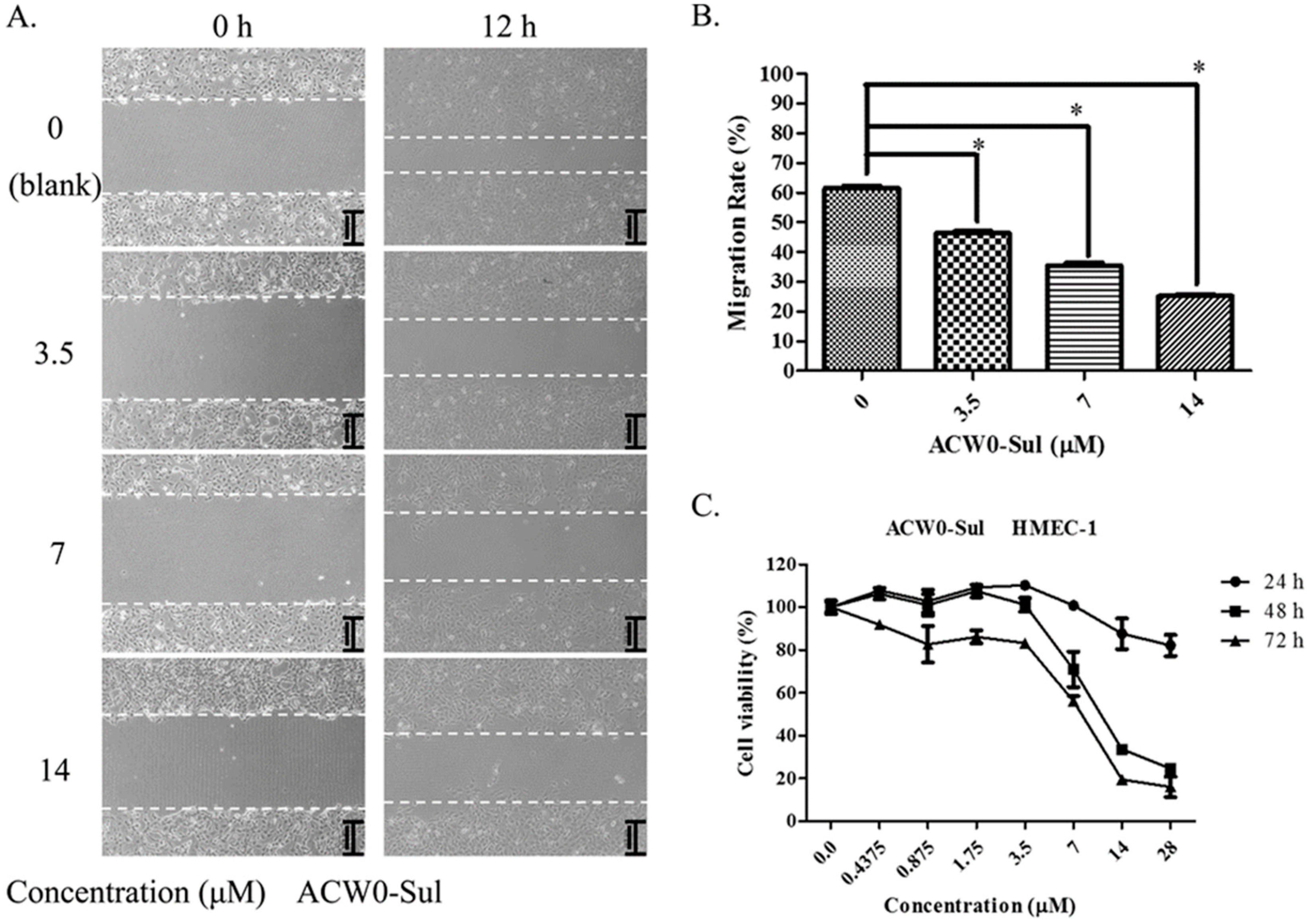
2.11. Wound Healing Assay of HMEC-1 Cells
2.12. Cell Proliferation (MTT) Assay
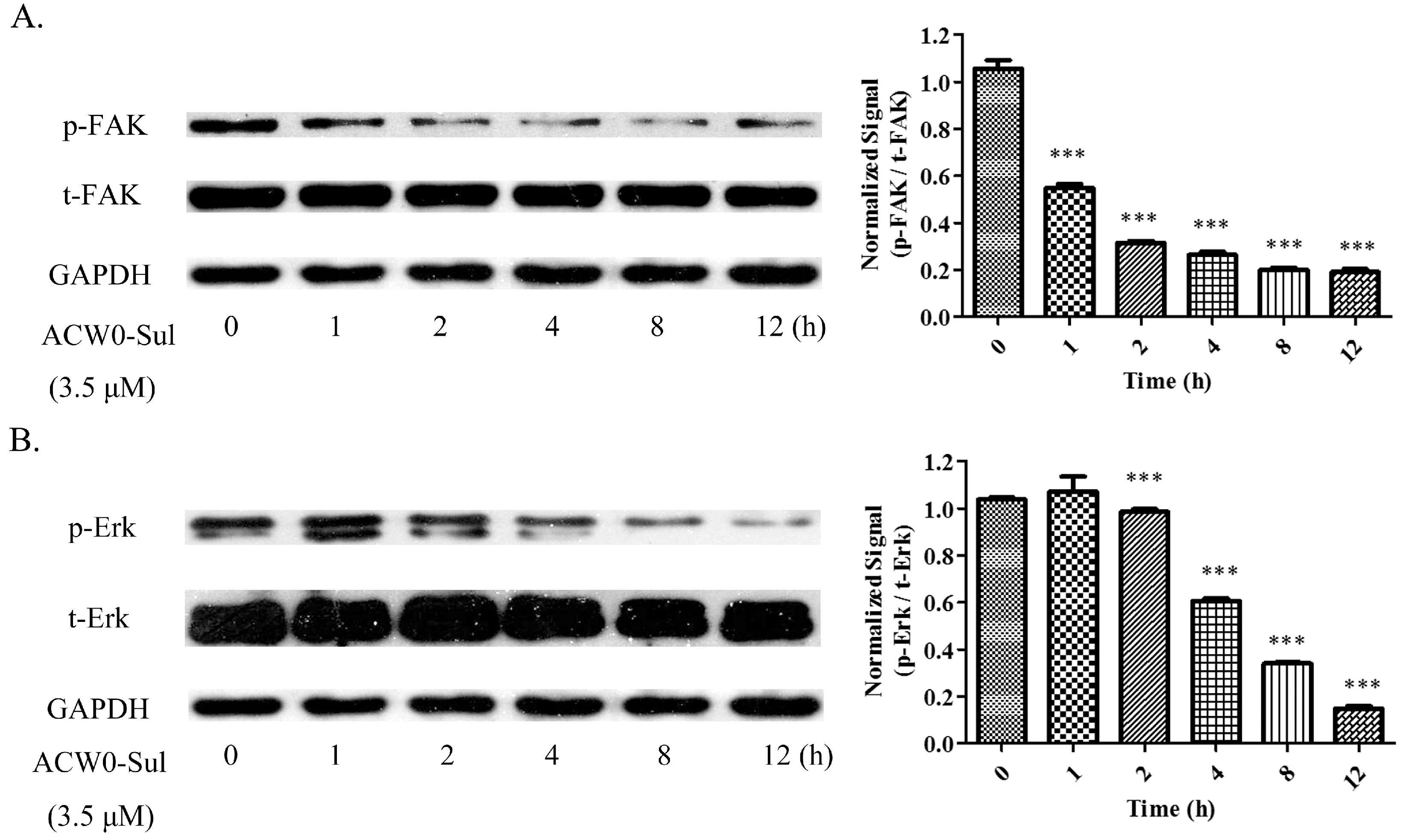
2.13. Western Blotting
2.14. Statistical Analysis
3. Results
3.1. Isolation and Purification of Polysaccharide
3.2. Composition and Linkage Analysis
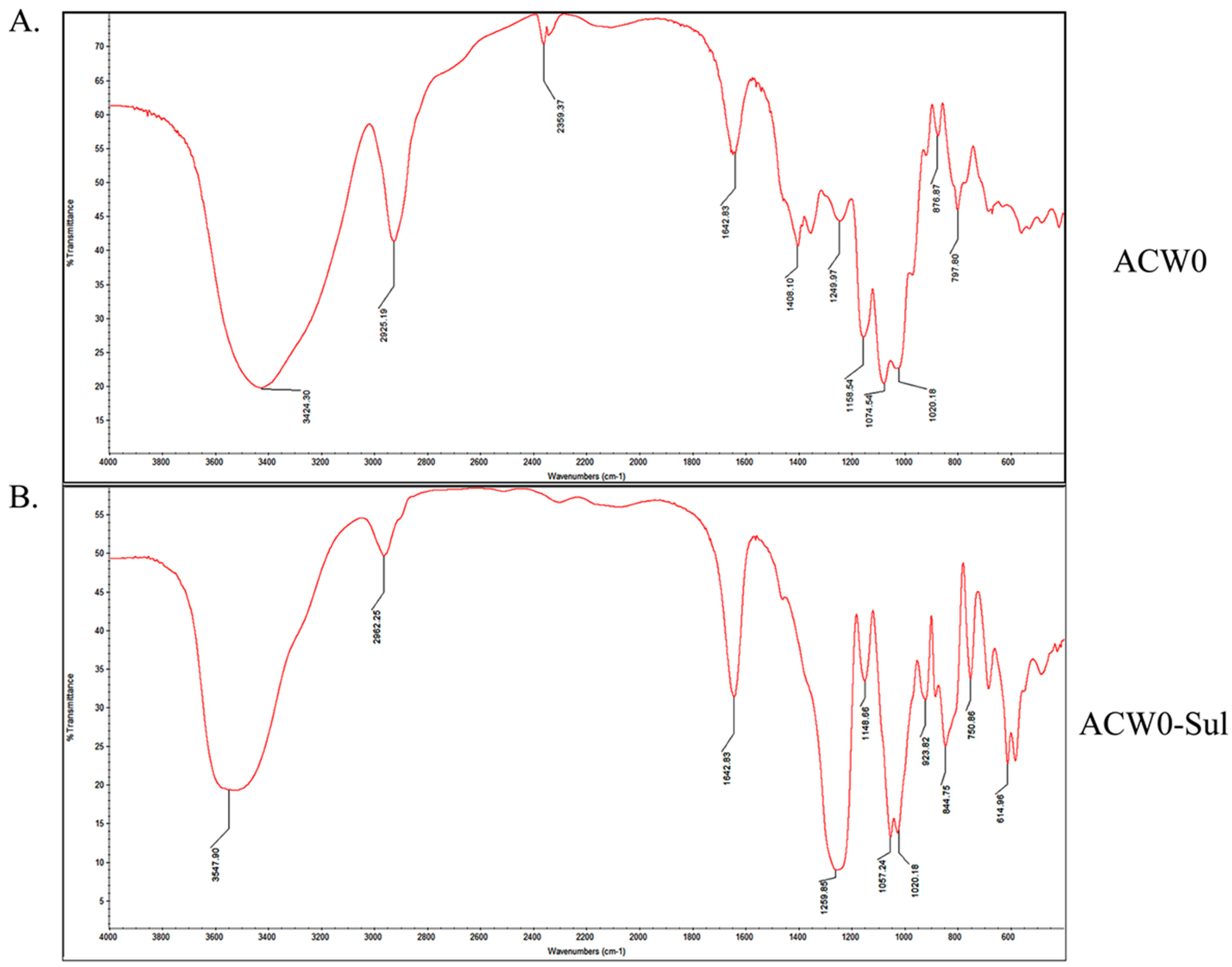
3.3. IR and Specific Rotation Analysis
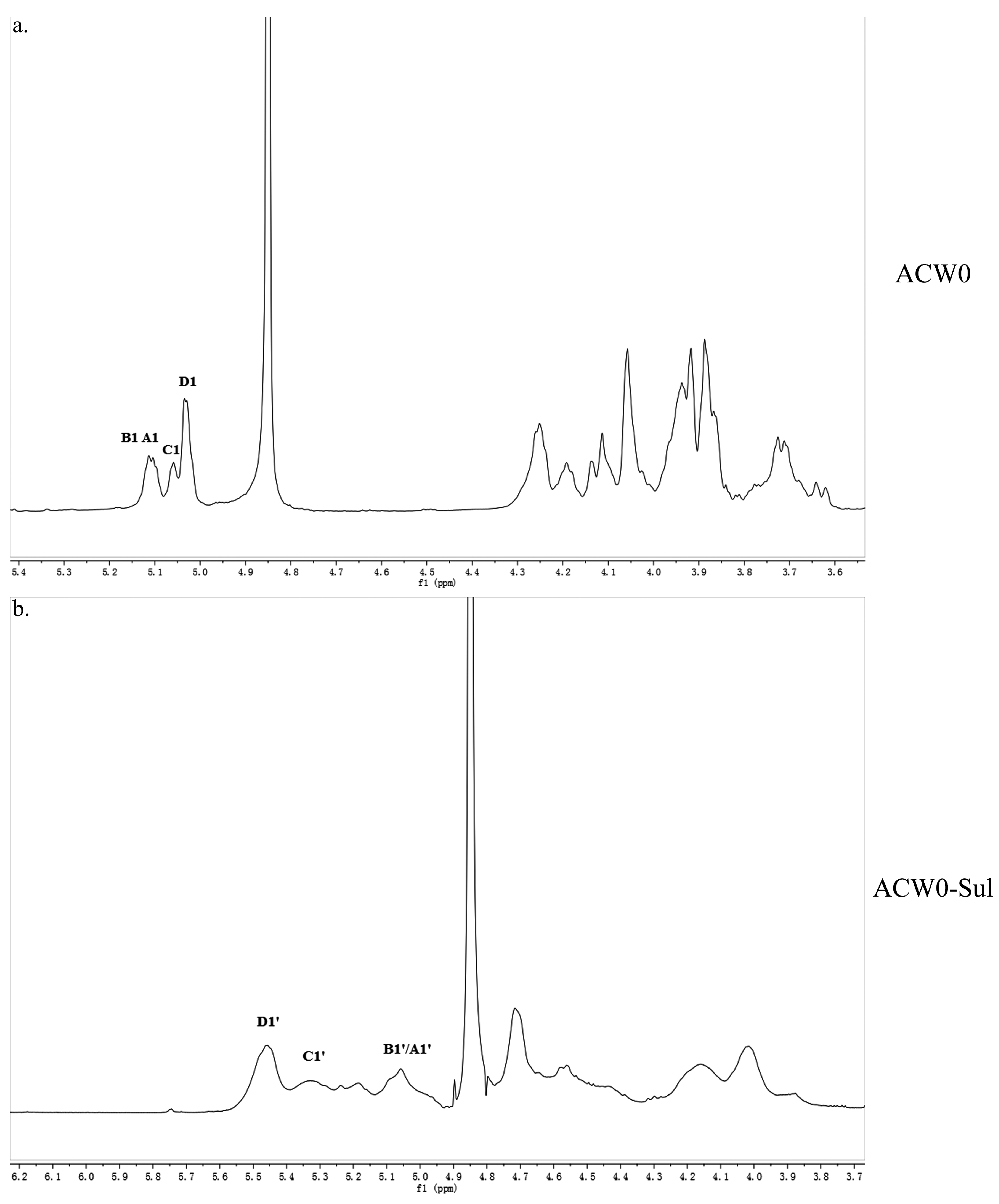
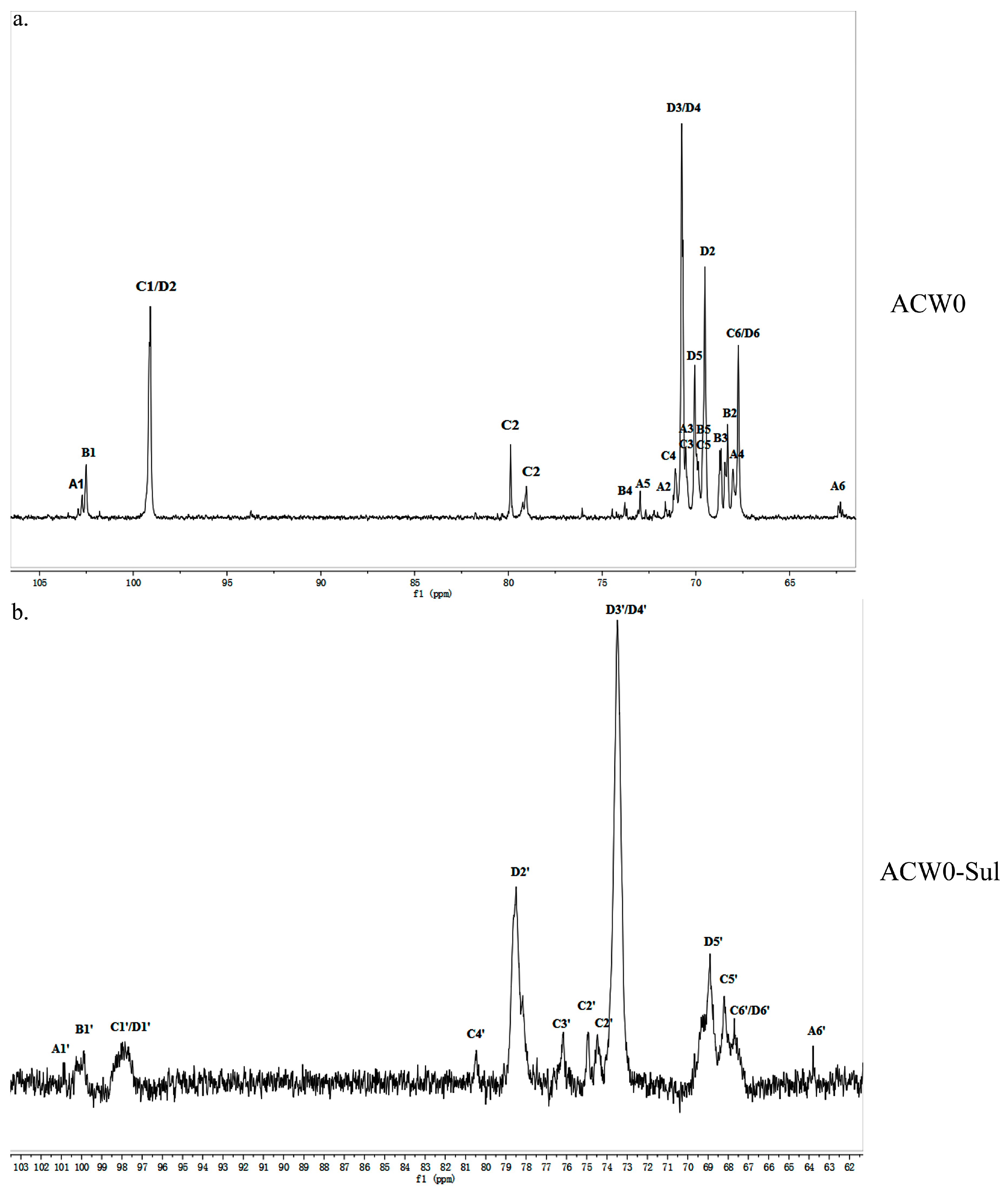
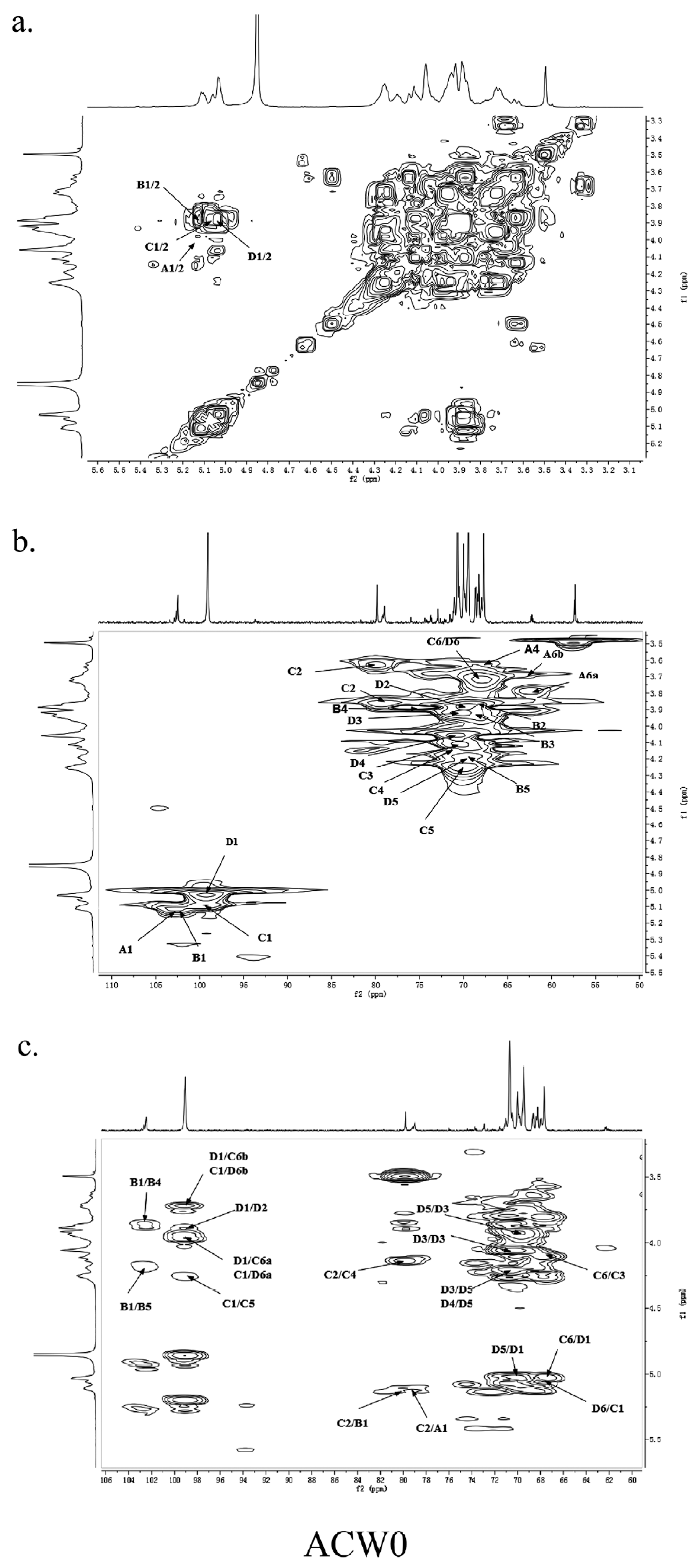
3.4. 1D and 2D NMR Analysis
3.5. Preparation and Analysis of Sulfated Derivative of ACW0
3.6. ACW0-Sul Inhibits HMEC-1 Tube Formation, Migration and Proliferation
3.7. ACW0-Sul Inhibits the Phosphorylation of Erk and FAK
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cuvillier, O. The therapeutic potential of HIF-2 antagonism in renal cell carcinoma. Transl. Androl. Urol. 2017, 6, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Watabe, K. The roles of microglia/macrophages in tumor progression of brain cancer and metastatic disease. Front. Biosci. 2017, 22, 1805–1829. [Google Scholar] [CrossRef]
- Escudero, C.A. Pro-angiogenic Role of Insulin: From Physiology to Pathology. Front. Physiol. 2017, 8, 204. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, L.; Yao, J.; Shi, Y.; Li, P.; Ding, K. An arabinogalactan from flowers of Panax notoginseng inhibits angiogenesis by BMP2/Smad/Id1 signaling. Carbohydr. Polym. 2015, 121, 328–335. [Google Scholar] [CrossRef]
- Qiu, H.; Yang, B.; Pei, Z.C.; Zhang, Z.; Ding, K. WSS25 inhibits growth of xenografted hepatocellular cancer cells in nude mice by disrupting angiogenesis via blocking bone morphogenetic protein (BMP)/Smad/Id1 signaling. J. Biol. Chem. 2010, 285, 32638–32646. [Google Scholar] [CrossRef]
- Chen, H.; Cong, Q.; Du, Z.; Liao, W.; Zhang, L.; Yao, Y.; Ding, K. Sulfated fucoidan FP08S2 inhibits lung cancer cell growth in vivo by disrupting angiogenesis via targeting VEGFR2/VEGF and blocking VEGFR2/Erk/VEGF signaling. Cancer Lett. 2016, 382, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, H.; Zhang, L.; Qin, Y.; Cong, Q.; Wang, P.; Ding, K. A fucoidan from Nemacystus decipiens disrupts angiogenesis through targeting bone morphogenetic protein 4. Carbohydr. Polym. 2016, 144, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Yang, Y.; Hu, X.; Zhang, Z. Effect of ultrasonic extraction conditions on antioxidative and immunomodulatory activities of a Ganoderma lucidum polysaccharide originated from fermented soybean curd residue. Food Chem. 2014, 155, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-C.; Song, T.-Y.; Shih, P.-H.; Yen, G.-C. Antioxidant properties of water-soluble polysaccharides from Antrodia cinnamomea in submerged culture. Food Chem. 2007, 104, 1115–1122. [Google Scholar] [CrossRef]
- Pan, C.; Wang, Y.; Qiu, M.K.; Wang, S.Q.; Liu, Y.B.; Quan, Z.W.; Ou, J.M. Knockdown of HMGB1 inhibits cell proliferation and induces apoptosis in hemangioma via downregulation of AKT pathway. J. Biol. Regul. Homeost. Agents 2017, 31, 41–49. [Google Scholar] [PubMed]
- Cheng, J.J.; Huang, N.K.; Chang, T.T.; Wang, D.L.; Lu, M.K. Study for anti-angiogenic activities of polysaccharides isolated from Antrodia cinnamomea in endothelial cells. Life Sci. 2005, 76, 3029–3042. [Google Scholar] [CrossRef]
- Yang, H.L.; Chen, C.S.; Chang, W.H.; Lu, F.J.; Lai, Y.C.; Chen, C.C.; Hseu, T.H.; Kuo, C.T.; Hseu, Y.C. Growth inhibition and induction of apoptosis in MCF-7 breast cancer cells by Antrodia camphorata. Cancer Lett. 2006, 231, 215–227. [Google Scholar] [CrossRef]
- Hseu, Y.C.; Wu, F.Y.; Wu, J.J.; Chen, J.Y.; Chang, W.H.; Lu, F.J.; Lai, Y.C.; Yang, H.L. Anti-inflammatory potential of Antrodia Camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-kappaB pathway. Int. Immunopharmacol. 2005, 5, 1914–1925. [Google Scholar] [CrossRef]
- Cho, S.M.; Yun, B.S.; Yoo, I.D.; Koshino, H. Structure of fomitellan A, a mannofucogalactan from the fruiting bodies of Fomitella fraxinea. Bioorg. Med. Chem. Lett. 2011, 21, 204–206. [Google Scholar] [CrossRef]
- Cheng, J.-J.; Lu, M.-K.; Lin, C.-Y.; Chang, C.-C. Characterization and functional elucidation of a fucosylated 1,6-α-d-mannogalactan polysaccharide from Antrodia cinnamomea. Carbohydr. Polym. 2011, 83, 545–553. [Google Scholar] [CrossRef]
- Fraser, R.N.; Karacsonyi, S.; Lindberg, B.; Levstek, I.; Eržen, V.; Blinc, R.; Paušak, S.; Ehrenberg, L.; Dumanović, J. Polysaccharides Elaborated by Polyporus pinicola (Fr). Acta Chem. Scand. 1967, 21, 1783–1789. [Google Scholar] [CrossRef]
- Alquini, G.; Carbonero, E.R.; Rosado, F.R.; Cosentino, C.; Iacomini, M. Polysaccharides from the fruit bodies of the basidiomycete Laetiporus sulphureus (Bull.: Fr.) Murr. FEMS Microbiol. Lett. 2004, 230, 47–52. [Google Scholar] [CrossRef]
- Fan, J.; Zhang, J.; Tang, Q.; Liu, Y.; Zhang, A.; Pan, Y. Structural elucidation of a neutral fucogalactan from the mycelium of Coprinus comatus. Carbohydr. Res. 2006, 341, 1130–1134. [Google Scholar] [CrossRef]
- Huang, X.; Wang, D.; Hu, Y.; Lu, Y.; Guo, Z.; Kong, X.; Sun, J. Effect of sulfated astragalus polysaccharide on cellular infectivity of infectious bursal disease virus. Int. J. Biol. Macromol. 2008, 42, 166–171. [Google Scholar] [CrossRef]
- Li, X.L.; Xiao, J.J.; Zha, X.Q.; Pan, L.H.; Asghar, M.N.; Luo, J.P. Structural identification and sulfated modification of an antiglycation Dendrobium huoshanense polysaccharide. Carbohydr. Polym. 2014, 106, 247–254. [Google Scholar] [CrossRef]
- Morimoto, M.; Takatori, M.; Hayashi, T.; Mori, D.; Takashima, O.; Yoshida, S.; Sato, K.; Kawamoto, H.; Tamura, J.; Izawa, H.; et al. Depolymerization of sulfated polysaccharides under hydrothermal conditions. Carbohydr. Res. 2014, 384, 56–60. [Google Scholar] [CrossRef]
- Wang, J.; Hu, Y.; Wang, D.; Liu, J.; Zhang, J.; Abula, S.; Zhao, B.; Ruan, S. Sulfated modification can enhance the immune-enhancing activity of lycium barbarum polysaccharides. Cell. Immunol. 2010, 263, 219–223. [Google Scholar] [CrossRef]
- Cong, Q.; Shang, M.; Dong, Q.; Liao, W.; Xiao, F.; Ding, K. Structure and activities of a novel heteroxylan from Cassia obtusifolia seeds and its sulfated derivative. Carbohydr. Res. 2014, 393, 43–50. [Google Scholar] [CrossRef]
- Tabatabai, M.A. A Rapid Method for Determination of Sulfate in Water Samples. Environ. Lett. 1974, 7, 237–243. [Google Scholar] [CrossRef]
- Maiese, K. Warming up to New Possibilities with the Capsaicin Receptor TRPV1: Mtor, AMPK, and Erythropoietin. Curr. Neurovasc. Res. 2017. [Google Scholar] [CrossRef]
- Chen, K.; Yan, M.; Li, Y.; Dong, Z.; Huang, D.; Li, J.; Wei, M. Intermedin1-53 enhances angiogenesis and attenuates adverse remodeling following myocardial infarction by activating AMP-activated protein kinase. Mol. Med. Rep. 2017, 15, 1497–1506. [Google Scholar] [CrossRef]
- Efferth, T. From ancient herb to modern drug: From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. Semin. Cancer Biol. 2017. [Google Scholar] [CrossRef]
- Lu, X.; Guo, S.; Cheng, Y.; Kim, J.H.; Feng, Y.; Feng, Y. Stimulation of ovarian follicle growth after AMPK inhibition. Reproduction 2017, 153, 683–694. [Google Scholar] [CrossRef]
- Inoue, K.; Kawamoto, K.; Nakajima, H.; Kohno, M.; Kadoya, S.; Mizuno, D. Structure-antitumor activity relationship of a d-manno-d-glucan from Microellobosporia grisea: Effect of periodate modification on antitumor activity. Carbohydr. Res. 1983, 115, 199–208. [Google Scholar] [CrossRef]
- Stelzl, T.; Geillinger-Kastle, K.E.; Stolz, J.; Daniel, H. Glycans in the intestinal peptide transporter PEPT1 contribute to function and protect from proteolysis. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G580–G591. [Google Scholar] [CrossRef]
- Haworth, W.S.; Swenson, V.J. Method and Device for Monitoring Platelet Function. U.S. Patent 20060269978A1, 30 November 2006. [Google Scholar]
- Haines, A.H. Synthesis of 6,6′-ether linked disaccharides from d-allose, d-galactose, d-glucose and d-mannose; evidence on the structure of coyolosa. Org. Biomol. Chem. 2004, 2, 2352–2358. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Bao, W.; Yang, J.; Zhang, T.; Sun, D.; Liang, Y.; Li, S.; Wang, Y.; Feng, X.; Hao, H.; et al. Focal Adhesion Kinase Directly Interacts with TSC2 through its FAT Domain and Regulates Cell Proliferation in Cashmere Goat Fetal Fibroblasts. DNA Cell Biol. 2016, 35, 480–488. [Google Scholar] [CrossRef]
- Song, B.Q.; Li, H.P.; Yuan, J.W.; Yang, X.; Wu, G.Y. Synthesis and Characterization of Bagasse Xylan Chlorosulfonate from the Chlorosulfonic Acid/Pyridine Method. Adv. Mater. Res. 2014, 1049–1050, 133–136. [Google Scholar] [CrossRef]
- Du, L.; Niu, H.; Zhang, X.; Xiao, D. Preparation of Glucan Sulfates with Different Degree of Substitution and Their Immunoprophylaxis Potentials in Escherichia coli Induced Mice Peritonitis. Int. Conf. Bioinform. Biomed. Eng. 2010, 1–4. [Google Scholar]
- Wei, D.; Wei, Y.; Cheng, W.; Zhang, L. Sulfated modification, characterization and antitumor activities of Radix hedysari polysaccharide. Int. J. Biol. Macromol. 2012, 51, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Vasil’eva, N.Y.; Levdansky, A.V.; Kuznetsov, B.N.; Skvortsova, G.P.; Kazachenko, A.S.; Djakovitch, L.; Pinel, C. Sulfation of arabinogalactan by sulfamic acid in dioxane. Russ. J. Bioorg. Chem. 2015, 41, 725–731. [Google Scholar] [CrossRef]
- Wang, L.; Huang, H.; Wei, Y.; Li, X.; Chen, Z. Characterization and anti-tumor activities of sulfated polysaccharide SRBPS2a obtained from defatted rice bran. Int. J. Biol. Macromol. 2009, 45, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Lechertier, T.; Hodivala-Dilke, K. Focal adhesion kinase and tumour angiogenesis. J. Pathol. 2012, 226, 404–412. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
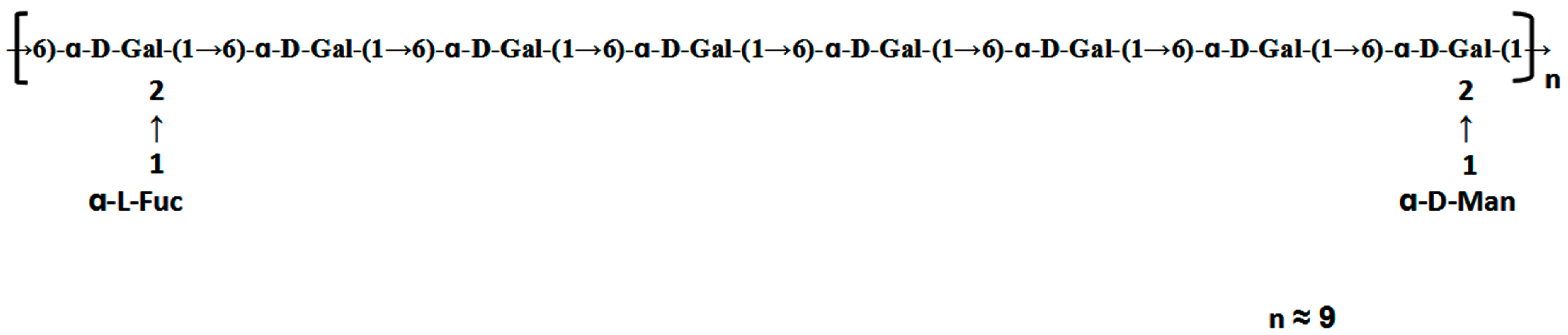{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methylated Sugars | Type of Linkages | Molar Ratios (%) | Mass Fragments (m/z) |
|---|---|---|---|
| 2,3,4,6-Me4-Man | T-Manp | 8.90 | 43,87,101,117,129,145,161,205 |
| 2,3,4,6-Me4-Fuc | T-Fucp | 5.38 | 43,89,101,117,131,161,175 |
| 3,4-Me2-Gal | 1,6-Galp | 66.92 | 43,99,101,117,129,161,189,233 |
| 2,3,4-Me3-Gal | 1,2,6-Galp | 18.80 | 43,87,99,129,189,233 |
| Residues | 1 | 2 | 3 | 4 | 5 | 6 | |
|---|---|---|---|---|---|---|---|
| A α-d-T-Manp | H | 5.11 | 4.02 | 3.68 | 3.63 | 3.89 | 3.78/3.68 |
| C | 102.73 | 71.55 | 70.47 | 67.93 | 72.98 | 62.22 | |
| B α-l-T-Fucp | H | 5.12 | 3.86 | 3.92 | 3.89 | 4.20 | 1.29 |
| C | 102.45 | 68.35 | 68.68 | 73.78 | 69.87 | 16.99 | |
| C α-d-1,2,6-Galp | H | 5.06 | 3.86/3.63 | 4.11 | 4.14 | 4.25 | 3.94/3.73 |
| C | 99.10 | 79.08/79.80 | 70.48 | 70.48 | 69.88 | 67.76 | |
| D α-d-1,6-Galp | H | 5.04 | 3.89 | 3.92 | 3.92 | 4.19 | 3.94/3.73 |
| C | 99.10 | 69.50 | 70.75 | 70.75 | 70.07 | 67.76 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Ding, Y.; Ye, M.; Zhu, T.; Tian, D.; Ding, K. A Novel Heterogalactan from Antrodia camphorata and Anti-Angiogenic Activity of Its Sulfated Derivative. Polymers 2017, 9, 228. https://doi.org/10.3390/polym9060228
Liu Y, Ding Y, Ye M, Zhu T, Tian D, Ding K. A Novel Heterogalactan from Antrodia camphorata and Anti-Angiogenic Activity of Its Sulfated Derivative. Polymers. 2017; 9(6):228. https://doi.org/10.3390/polym9060228
Chicago/Turabian StyleLiu, Yanqiu, Yaqi Ding, Min Ye, Tao Zhu, Danbi Tian, and Kan Ding. 2017. "A Novel Heterogalactan from Antrodia camphorata and Anti-Angiogenic Activity of Its Sulfated Derivative" Polymers 9, no. 6: 228. https://doi.org/10.3390/polym9060228
APA StyleLiu, Y., Ding, Y., Ye, M., Zhu, T., Tian, D., & Ding, K. (2017). A Novel Heterogalactan from Antrodia camphorata and Anti-Angiogenic Activity of Its Sulfated Derivative. Polymers, 9(6), 228. https://doi.org/10.3390/polym9060228