A Comparative Study of Field Nematode Communities over a Decade of Cotton Production in Australia


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Soil Characteristics and Nematode Sampling
2.2. Soil Analysis
2.3. Root Tissue Analysis
2.4. Community Comparisons and Statistical Analysis
3. Results
3.1. Soil Sample and Total Nematode Comparisons
3.2. Plant Parasitic Nematode Populations
3.3. Nematode Community Assemblages
3.4. Root Tissue Observations
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Skevas, T.; Lansink, A.; Stefanou, S.E. Designing the Emerging Eu Pesticide Policy: A Literature Review. NJAS-Wagening. J. Life Sci. 2013, 64, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Bielza, P.; Denholm, I.; Sterk, G.; Leadbeater, A.; Leonard, P.; Jørgensen, L.N. Declaration of Ljubljana #8211; the Impact of a Declining European Pesticide Portfolio on Resistance Management. Outlooks Pest Manag. 2008, 19, 246–248. [Google Scholar]
- Erickson, B. Pesticides: Bayer Cropscience, Epa Agree to Phase out Use of Aldicarb. Chem. Eng. News 2010, 88. [Google Scholar] [CrossRef]
- Pesticides, A.; Authority, V.M. The Nra Review of Aldicarb. In Existing Chemical Review Program; National Registration Authority for Agricultural and Veterinary Chemicals: Canberra, Australia, 2001. [Google Scholar]
- Cox, C. Aldicarb. J. Pestic. Reform 1992, 12, 31–35. [Google Scholar]
- Robinson, A.F. Nematode Management in Cotton. In Integrated Management and Biocontrol of Vegetable and Grain Crops Nematodes; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 149–182. [Google Scholar]
- Bauer, B.; Smith, L.; Scheikowski, L.; Lehane, J.; Cobon, J.; O’Neill, W. Reniform Nematode Surveys in Central Queensland Cotton. In Proceedings of the 17th Australian Cotton Conference, Gold Coast, Australian, 5 August 2014. [Google Scholar]
- Farrell, T. Cotton Pest Management Guide 2007–2008; NSW Department of Primary Industries: Orange, Australian, 2007.
- CSD. New Cotton Seed Treatment Reduces in-Furrow Woes. Seeds Thought 2011, 10, 19. [Google Scholar]
- Koppenhöfer, M.A.; Fuzy, E.M. Early Timing and New Combinations to Increase the Efficacy of Neonicotinoid–Entomopathogenic Nematode (Rhabditida: Heterorhabditidae) Combinations against White Grubs (Coleoptera: Scarabaeidae). Pest Manag. Sci. 2008, 64, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Tahseen, Q.; Shamim Jairajpuri, M.; Ahmad, I. Nematicidal Impact on the Reproductive Biology of Mesorhabditis Cranganorensis. Afro-Asian J. Nematol. 1996, 6, 184–187. [Google Scholar]
- Meher, C.H.; Gajbhiye, V.T.; Singh, G.; Kamra, A.; Chawla, G. Persistence and Nematicidal Efficacy of Carbosulfan, Cadusafos, Phorate, and Triazophos in Soil and Uptake by Chickpea and Tomato Crops under Tropical Conditions. J. Agric. Food Chem. 2010, 58, 1815–1822. [Google Scholar] [CrossRef]
- Wada, S.; Toyota, K.; Takada, A. Effects of the Nematicide Imicyafos on Soil Nematode Community Structure and Damage to Radish Caused by Pratylenchus Penetrans. J. Nematol. 2011, 43, 1–6. [Google Scholar]
- Khan, R.M.; Zaidi, B.; Haque, Z. Nematicides Control Rice Root-Knot, Caused by Meloidogyne Graminicola. Phytopathol. Mediterr. 2012, 2012, 298–306. [Google Scholar]
- Soltani, A.; Louati, H.; Hanachi, A.; Salem, F.B.; Essid, N.; Aissa, P.; Mahmoudi, E.; Beyrem, H. Impacts of Permethrin Contamination on Nematode Density and Diversity: A Microcosm Study on Benthic Meiofauna from a Mediterranean Coastal Lagoon. Biologia 2012, 67, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Hara, A.H.; Kaya, H.K. Effects of Selected Insecticides and Nematicides on the in Vitro Development of the Entomogenous Nematode Neoaplectana Carpocapsae. J. Nematol. 1982, 14, 486–491. [Google Scholar] [PubMed]
- Sabino, P.H.; Sales, F.S.; Guevara, E.J.; Moino, A., Jr.; Filgueiras, C.C. Compatibility of Entomopathogenic Nematodes (Nematoda: Rhabditida) with Insecticides Used in the Tomato Crop. Nematoda 2014, 1. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Shono, T.; Yamanaka, S.; Tanabe, H. Effects of Insecticides on the Entomopathogenic Nematode Steinernema Carpocapsae Weiser. Appl. Entomol. Zool. 1994, 29, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Cox, C. Cyfluthrin. J. Pestic. Reform 1994, 14, 28–34. [Google Scholar]
- Allen, S.; Smith, L.; Scheikowski, L.; Gambley, C.; Sharman, M.; Maas, S.; Kirkby, K.; Lonergan, P. Common Diseases of Cotton. In Cotton Pest Management Guide 2014–15; Susan, M., Ed.; Greenmount Press: East Toowoomba, Australia, 2014; pp. 118–121. [Google Scholar]
- Blessitt, A.J.; Stetina, S.R.; Wallace, T.P.; Smith, P.T.; Sciumbato, G.L. Cotton (Gossypium Hirsutum) Cultivars Exhibiting Tolerance to the Reniform Nematode (Rotylenchulus Reniformis). Int. J. Agron. 2012. [Google Scholar] [CrossRef] [Green Version]
- Knox, G.O.G.; Anderson, C.M.T.; Nehl, D.B.; Gupta, V.V.S.R. Observation of Tylenchorhynchus Ewingi in Association with Cotton Soils in Australia. Plant Dis.Notes 2006, 1, 47–48. [Google Scholar] [CrossRef] [Green Version]
- Knox, O.; Anderson, C.; Vadakattu, G.; Seymour, N. Tiny Worms: Nematodes in Australian Cotton. Aust. Cottongrow. 2007, 28, 10–13. [Google Scholar]
- Lue, P.L.; Lewis, C.C.; Melchor, V.E. The Effect of Aldicarb on Nematode Population and Its Persistence in Carrots, Soil and Hydroponic Solution. J. Environ. Sci. Health B 1984, 19, 343–354. [Google Scholar] [CrossRef]
- Sturz, A.; Kimpinski, J. Effects of Fosthiazate and Aldicarb on Populations of Plant-Growth-Promoting Bacteria, Root-Lesion Nematodes and Bacteria-Feeding Nematodes in the Root Zone of Potatoes. Plant Pathol. 1999, 48, 26–32. [Google Scholar] [CrossRef]
- Knox, G.O.G.; Anderson, C.M.T.; Allen, S.J.; Nehl, D.B. Helicotylenchus Dihystera in Australian Cotton Roots. Australas. Plant Pathol. 2006, 35, 287–288. [Google Scholar] [CrossRef]
- Stirling, R.G.; Lodge, G.M. A Survey of Australian Temperate Pastures in Summer and Winter Rainfall Zones: Soil Nematodes, Chemical, and Biochemical Properties. Soil Res. 2005, 43, 887–904. [Google Scholar] [CrossRef]
- Byrd, W.B.; Kirkpatrick, T.L.; Barker, K.R. An Improved Technique for Clearing and Staining Plant Tissues for Detection of Nematodes. J. Nematol. 1983, 15, 142–143. [Google Scholar]
- Yeates, G.W. Nematodes as Soil Indicators: Functional and Biodiversity Aspects. Biol. Fertil. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G.M. Ninja: An Automated Calculation System for Nematode-Based Biological Monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Bongers, T. The Maturity Index: An Ecological Measure of Environmental Disturbance Based on Nematode Species Composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A Framework for Soil Food Web Diagnostics: Extension of the Nematode Faunal Analysis Concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Knox, G.O.G.; Gupta, V.V.S.R.; Lardner, R. Field Evaluation of the Effects of Cotton Variety and Gm Status on Rhizosphere Microbial Diversity and Function in Australian Soils. Soil Res. 2014, 52, 203–215. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T. Nematode Diversity in Agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 113–135. [Google Scholar] [CrossRef]
- Coppedge, R.J.; Bull, D.L.; Ridgway, R.L. Movement and Persistence of Aldicarb in Certain Soils. Arch. Environ. Contam. Toxicol. 1977, 5, 129–141. [Google Scholar] [CrossRef]
- Brmez, M.; Ivezic, M.; Raspudic, E.; Tripar, V.; Balicevic, R. Nematode Communities as Bioindicators of Antropogenic Influence in Agroecosystems. Cereal Res. Commun. 2007, 35, 297–300. [Google Scholar] [CrossRef]
- Čerevková, A.; Miklisová, D.; Cagáň, Ľ. Effects of Experimental Insecticide Applications and Season on Soil Nematode Communities in a Maize Field. Crop Prot. 2017, 92, 1–15. [Google Scholar] [CrossRef]
- Farrell, T.; Hulugalle, N.; Gett, V. Healthier Cotton Soils through High Input Cereal Rotations. In Proceedings of the 14th Australian Cotton Conference, Broadbeach, Australia, 12–14 August 2008. [Google Scholar]
- Bell, M.; Seymour, N.; Stirling, G.R.; Stirling, A.M.; van Zwieten, L.; Vancov, T.; Sutton, G.; Moody, P. Impacts of Management on Soil Biota in Vertosols Supporting the Broadacre Grains Industry in Northern Australia. Aust. J. Soil Res. 2006, 44, 433–451. [Google Scholar] [CrossRef] [Green Version]
- McSorley, R. Overview of Organic Amendments for Management of Plant-Parasitic Nematodes, with Case Studies from Florida. J. Nematol. 2011, 43, 69–81. [Google Scholar] [PubMed]
- Braunack, M.V. Cotton Farming Systems in Australia: Factors Contributing to Changed Yield and Fibre Quality. Crop Pasture Sci. 2013, 64, 834–844. [Google Scholar] [CrossRef]
- Yeates, W.G.; Wardle, D.A.; Watson, R.N. Responses of Soil Nematode Populations, Community Structure, Diversity and Temporal Variability to Agricultural Intensification over a Seven-Year Period. Soil Biol. Biochem. 1999, 31, 1721–1733. [Google Scholar] [CrossRef]
- Ferris, V.R.; Bernard, R.L. Crop Rotation Effects on Population Densities of Ectoparasitic Nematodes. J. Nematol. 1971, 3, 119–122. [Google Scholar]
- McLeod, R.; Reay, F.; Smith, J. Plant Nematodes of Australia Listed by Plant and by Genus: Compiled 1994; Francoise, R., Smyth, J., Eds.; NSW Agriculture: Orange, Australia, 1994.
- Villenave, C.; Saj, S.; Pablo, A.; Sall, S.; Djigal, D.; Chotte, J.; Bonzi, M. Influence of Long-Term Organic and Mineral Fertilization on Soil Nematofauna When Growing Sorghum Bicolor in Burkina Faso. Biol. Fertil. Soils 2010, 46, 659–670. [Google Scholar] [CrossRef]
- Zahid, I.M.; Gurr, G.M.; Hodda, M.; Nikandrow, A.; Fulkerson, W.J. Orientation, Reproduction and Effect of Spiral Nematode (Helicotylenchus Dihystera) on Growth of White Clover (Cv. Haifa). Australas. Plant Pathol. 2002, 31, 55–56. [Google Scholar] [CrossRef]
- Taylor, P.S.; Vanstone, V.A.; Ware, A.H.; McKay, A.C.; Szot, D.; Russ, M.H. Measuring Yield Loss in Cereals Caused by Root Lesion Nematodes (Pratylenchus Neglectus and P. Thornei) with and without Nematicide. Aust. J. Agric. Res. 1999, 50, 617–627. [Google Scholar] [CrossRef]
- Thompson, P.J.; Owen, K.J.; Stirling, G.R.; Bell, M.J. Root-Lesion Nematodes (Pratylenchus Thornei and P. Neglectus): A Review of Recent Progress in Managing a Significant Pest of Grain Crops in Northern Australia. Australas. Plant Pathol. 2008, 37, 235–242. [Google Scholar] [CrossRef]
- Small, R.W. A Review of the Prey of Predatory Soil Nematodes. Pedobiologia 1987, 30, 179–206. [Google Scholar]
- Yeates, W.G.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding Habits in Soil Nematode Families and Genera—An Outline for Soil Ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Ferris, H.; Griffiths, B.S.; Porazinska, D.L.; Powers, T.O.; Wang, K.; Tenuta, M. Reflections on Plant and Soil Nematode Ecology: Past, Present and Future. J. Nematol. 2012, 44, 115–126. [Google Scholar] [PubMed]


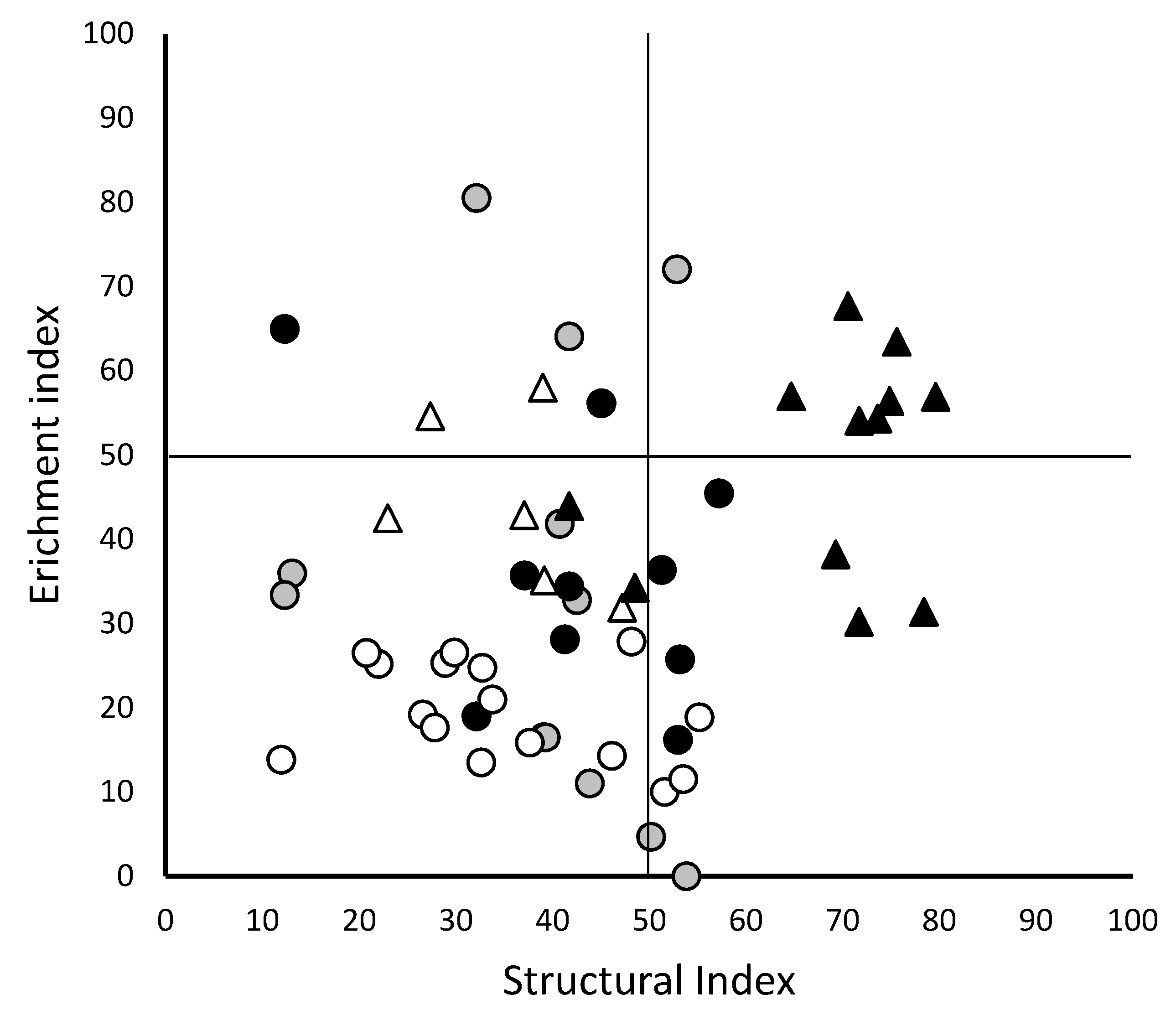
 ) and 2015 (●) and field B sampled in 2005 (
) and 2015 (●) and field B sampled in 2005 (  ) and 2015 (
) and 2015 (  ). The Enrichment index parallels with the nutrient enrichment whilst the structural index correlates with the maturity of the ecosystem.
) and 2015 (●) and field B sampled in 2005 ( ) and 2015 ( ). The Enrichment index parallels with the nutrient enrichment whilst the structural index correlates with the maturity of the ecosystem.
). The Enrichment index parallels with the nutrient enrichment whilst the structural index correlates with the maturity of the ecosystem.
) and 2015 (●) and field B sampled in 2005 ( ) and 2015 ( ). The Enrichment index parallels with the nutrient enrichment whilst the structural index correlates with the maturity of the ecosystem.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field | Year | Nematodes Per 200 mL | Per g/Soil | % Plant Parasitic | % Stunt | % Lesion | % Spiral | % Dagger |
|---|---|---|---|---|---|---|---|---|
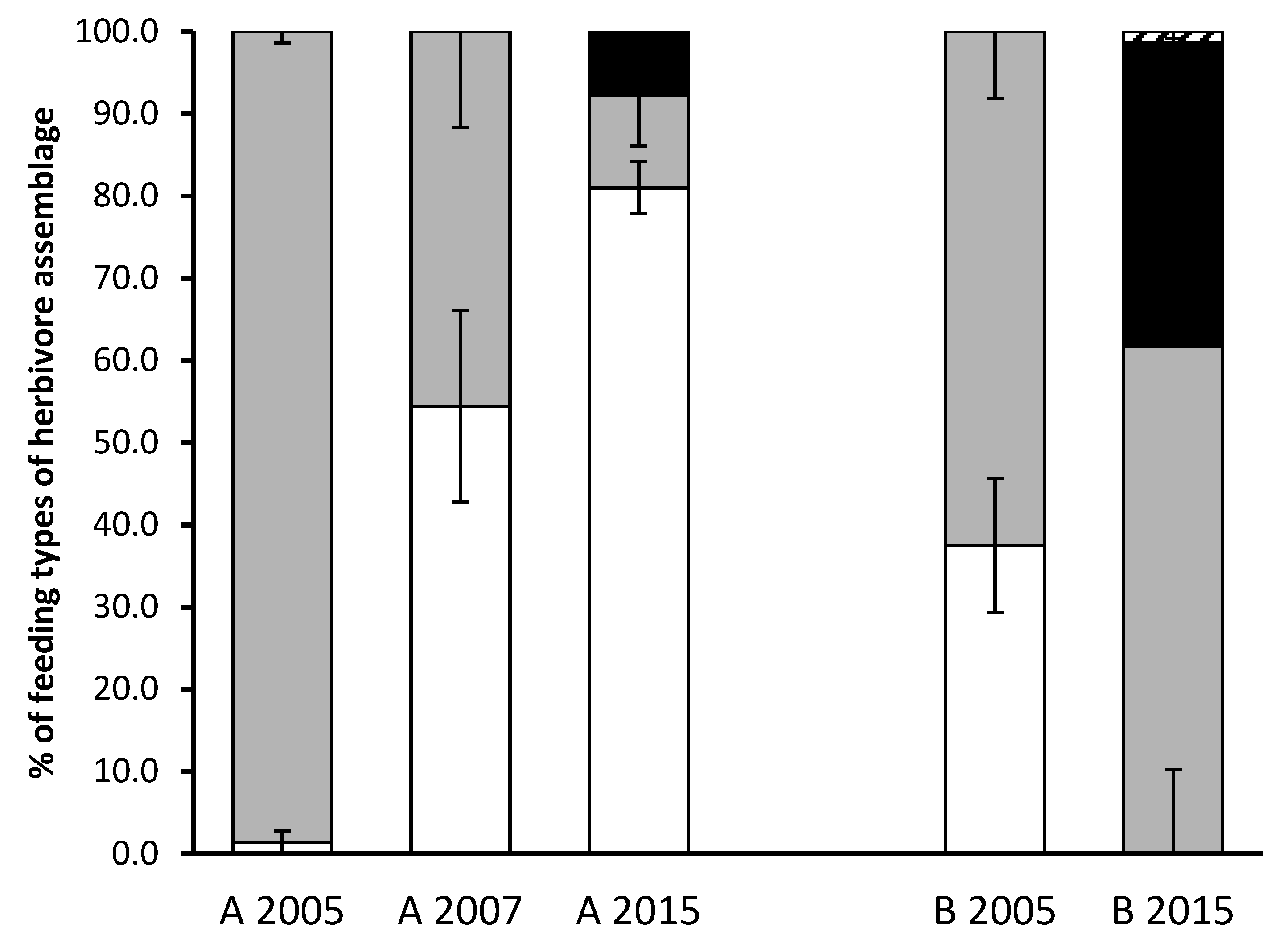
| A | 2005 | 1064 | 8.5 | 2.9 | 92.4 A | 1.3 A | nd | nd |
| 2007 | 791 | 6.3 | 1.0 | 41.8 B | 49.9 B | nd | nd | |
| 2015 | 1319 | 10.2 | 1.6 | 11.3 B | 81 B | 7.7 | nd | |
| ns | ns | ns | p < 0.01 | p < 0.001 | ns | ns | ||
| B | 2005 | 1229 | 9.8 | 1.1 | 100.0 | nd | nd | nd |
| 2015 | 1687 | 11.8 | 7.7 | 61.7 | nd | 36.8 | 1.4 | |
| ns | ns | p < 0.001 | p < 0.001 | p < 0.01* | ns |
| Index Name | Field A 2005 | Field A 2007 | Field A 2015 | Field B 2005 | Field B 2015 | ANOVA p Value | |
|---|---|---|---|---|---|---|---|
| Maturity Index | mean | 2.3 | 2.2 | 2.2 | 2.1 | 2.4 | <0.001 |
| SD | 0.2 | 0.2 | 0.1 | 0.1 | 0.1 | ||
| Plant Parasitic Index | mean | 2.4 | 2.6 | 2.1 | 2.3 | 3.2 | <0.001 |
| SD | 0.3 | 0.3 | 0.1 | 0.4 | 0.5 | ||
| Enrichment Index | mean | 35.6 | 26.1 | 37.4 | 44.3 | 49.2 | 0.001 |
| SD | 6.2 | 17.6 | 14.3 | 10.6 | 12.9 | ||
| Structure Index | mean | 51.3 | 38.5 | 47.5 | 35.5 | 68.4 | <0.001 |
| SD | 15.2 | 13.6 | 7.6 | 8.8 | 11.7 | ||
| Nematode channel ratio | mean | 0.7 | 0.9 | 0.9 | 0.7 | 0.9 | <0.001 |
| SD | 0.1 | 0.1 | 0.0 | 0.1 | 0.1 | ||
| Herbivore footprint | mean | 1.5 | 1.5 | 3.2 | 2.0 | 21.0 | <0.001 |
| SD | 0.6 | 0.6 | 1.4 | 0.8 | 10.5 | ||
| Fungivore footprint | mean | 3.2 | 1.3 | 0.7 | 2.2 | 0.8 | <0.001 |
| SD | 1.5 | 0.7 | 0.5 | 0.8 | 0.5 | ||
| Bacterivore footprint | mean | 14.4 | 22.5 | 24.2 | 24.5 | 15.2 | 0.009 |
| SD | 3.8 | 13.4 | 5.3 | 10.6 | 3.4 | ||
| Predator footprint | mean | 1.6 | 0.5 | 0.7 | 0.0 | 0.7 | 0.09 |
| SD | 1.9 | 1.0 | 0.9 | 0.0 | 0.9 | ||
| Omnivore footprint | mean | 11.8 | 7.7 | 5.3 | 6.6 | 5.7 | 0.044 |
| SD | 8.6 | 3.8 | 2.4 | 5.8 | 2.8 | ||
| Total number | mean | 132.6 | 118.8 | 120.3 | 119.2 | 123.8 | 0.045 |
| (nematode/200 mL) | SD | 11.6 | 15.7 | 9.0 | 15.0 | 11.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knox, O.; Backhouse, D.; Gupta, V. A Comparative Study of Field Nematode Communities over a Decade of Cotton Production in Australia. Agronomy 2020, 10, 123. https://doi.org/10.3390/agronomy10010123
Knox O, Backhouse D, Gupta V. A Comparative Study of Field Nematode Communities over a Decade of Cotton Production in Australia. Agronomy. 2020; 10(1):123. https://doi.org/10.3390/agronomy10010123
Chicago/Turabian StyleKnox, Oliver, David Backhouse, and Vadakattu Gupta. 2020. "A Comparative Study of Field Nematode Communities over a Decade of Cotton Production in Australia" Agronomy 10, no. 1: 123. https://doi.org/10.3390/agronomy10010123
APA StyleKnox, O., Backhouse, D., & Gupta, V. (2020). A Comparative Study of Field Nematode Communities over a Decade of Cotton Production in Australia. Agronomy, 10(1), 123. https://doi.org/10.3390/agronomy10010123