Identification and Verification of Quantitative Trait Loci Affecting Milling Yield of Rice

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Experiment and Trait Measurement
2.3. Marker Data and Genetic Maps
2.4. Data Analysis
3. Results
3.1. Phenotypic Performance of the Three RIL Populations
3.2. QTL Detected in the Three RIL Populations
3.3. QTLs Detected in the TI Population
3.4. QTLs Detected in the ZM Population
3.5. QTLs Detected in the XM Population
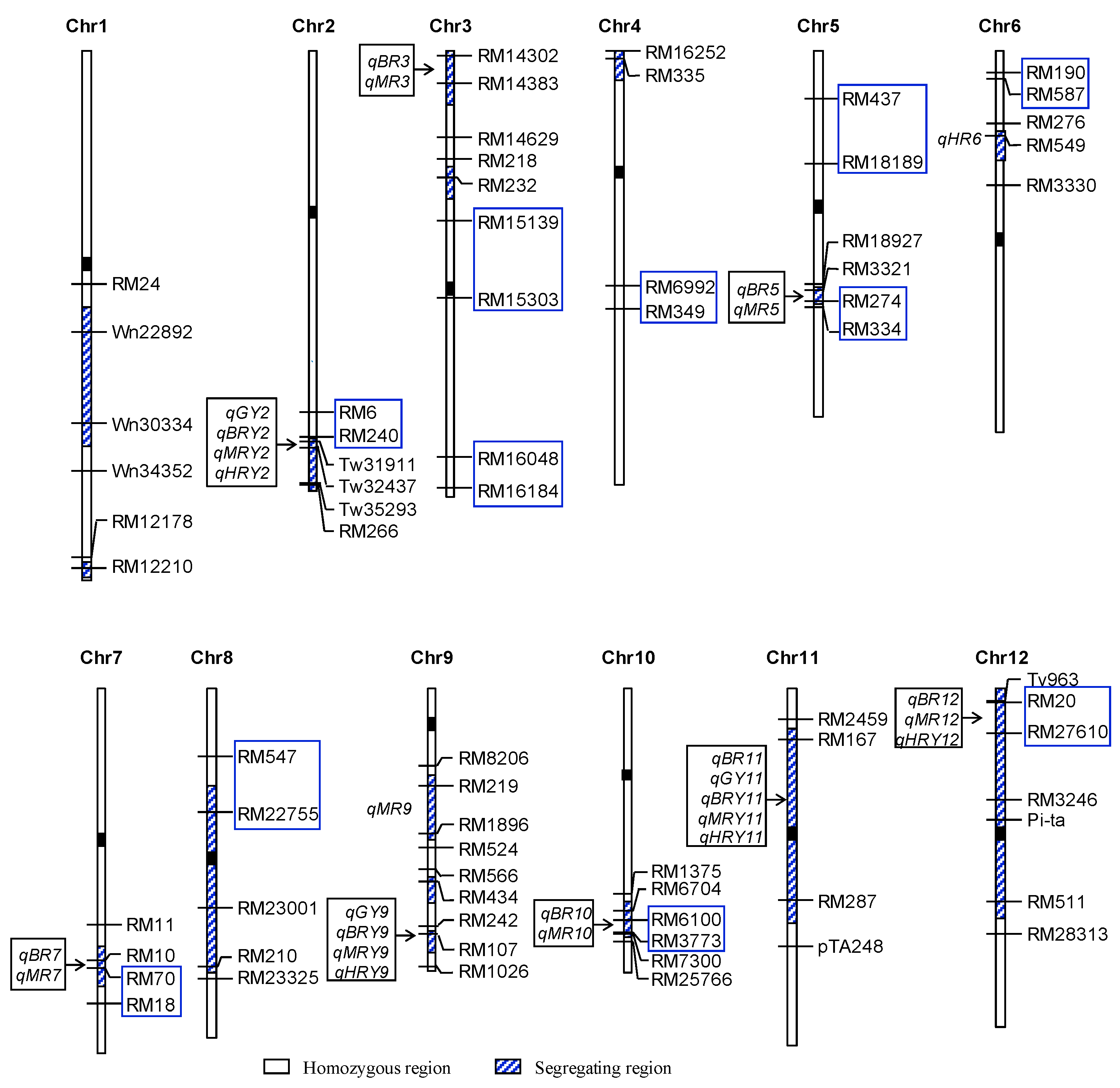
3.6. Validation of Five QTL Regions in an RH-Derived F4:5 Population
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nalley, L.; Tack, J.; Barkley, A.; Jagadish, K.; Brye, K. Quantifying the agronomic and economic performance of hybrid and conventional rice varieties. Agron. J. 2016, 108, 1514–1523. [Google Scholar] [CrossRef] [Green Version]
- Aluko, G.; Martinez, C.; Tohme, J.; Castano, C.; Bergman, C.; Oard, J.H. QTL mapping of grain quality traits from the interspecific cross Oryza sativa x O. glaberrima. Theor. Appl. Genet. 2004, 109, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Kepiro, J.L.; McClung, A.M.; Chen, M.H.; Yeater, K.M.; Fjellstrom, R.G. Mapping QTLs for milling yield and grain characteristics in a tropical japonica long grain cross. J. Cereal Sci. 2008, 48, 477–485. [Google Scholar] [CrossRef]
- Bao, J. Genes and QTLs for rice grain quality improvement. In Rice-Germplasm, Genetics and Improvement; Yan, W., Bao, J., Eds.; Intech: Rijeka, Croatia, 2014; pp. 239–278. [Google Scholar]
- Hu, X.; Shi, Y.-M.; Jia, Q.; Xu, Q.; Wang, Y.; Chen, K.; Sun, Y.; Zhu, L.-H.; Xu, J.-L.; Li, Z.-K. Analyses of QTLs for rice panicle and milling quality traits and their interaction with environment. Acta Agron. Sin. 2011, 37, 1175–1185. [Google Scholar]
- Li, Z.F.; Wan, J.M.; Xia, J.F.; Zhai, H.Q.; Ikehashi, H. Identification of quantitative trait loci underlying milling quality of rice (Oryza sativa) grains. Plant Breed. 2004, 123, 229–234. [Google Scholar] [CrossRef]
- Nelson, J.C.; McClung, A.M.; Fjellstrom, R.G.; Moldenhauer, K.A.K.; Boza, E.; Jodari, F.; Oard, J.H.; Linscombe, S.; Scheffler, B.E.; Yeater, K.M. Mapping QTL main and interaction influences on milling quality in elite US rice germplasm. Theor. Appl. Genet. 2011, 122, 291–309. [Google Scholar] [CrossRef]
- Nelson, J.C.; Jodari, F.; Roughton, A.I.; McKenzie, K.M.; McClung, A.M.; Fjellstrom, R.G.; Scheffler, B.E. QTL mapping for milling quality in elite western U.S. rice germplasm. Crop Sci. 2012, 52, 242–252. [Google Scholar] [CrossRef]
- Tan, Y.F.; Sun, M.; Xing, Y.Z.; Hua, J.P.; Sun, X.L.; Zhang, Q.F.; Corke, H. Mapping quantitative trait loci for milling quality, protein content and color characteristics of rice using a recombinant inbred line population derived from an elite rice hybrid. Theor. Appl. Genet. 2001, 103, 1037–1045. [Google Scholar] [CrossRef]
- Dong, Y.; Tsuzuki, E.; Lin, D.; Kamiunten, H.; Terao, H.; Matsuo, M.; Cheng, S. Molecular genetic mapping of quantitative trait loci for milling quality in rice (Oryza sativa L.). J. Cereal Sci. 2004, 40, 109–114. [Google Scholar] [CrossRef]
- Zheng, T.Q.; Xu, J.L.; Li, Z.K.; Zhai, H.Q.; Wan, J.M. Genomic regions associated with milling quality and grain shape identified in a set of random introgression lines of rice (Oryza sativa L.). Plant Breed. 2007, 126, 158–163. [Google Scholar] [CrossRef]
- Lou, J.; Chen, L.; Yue, G.; Lou, Q.; Mei, H.; Xiong, L.; Luo, L. QTL mapping of grain quality traits in rice. J. Cereal Sci. 2009, 50, 145–151. [Google Scholar] [CrossRef]
- Mei, D.-Y.; Zhu, Y.-J.; Yu, Y.-H.; Fan, Y.-Y.; Huang, D.-R.; Zhuang, J.-Y. Quantitative trait loci for grain chalkiness and endosperm transparency detected in three recombinant inbred line populations of indica rice. J. Integr. Agric. 2013, 12, 1–11. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, J.-Y.; Zhu, Y.-J.; Fan, Y.-Y.; Zhuang, J.-Y. Validation of qGS10, a quantitative trait locus for grain size on the long arm of chromosome 10 in rice (Oryza sativa L.). J. Integr. Agric. 2017, 16, 20–30. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, Y.-J.; Fan, Y.-Y.; Huang, T.-X.; Zhang, J.-F.; Xie, H.-A.; Zhuang, J.-Y. Identification and verification of quantitative trait loci for eating and cooking quality of rice (Oryza sativa). Plant Breed. 2019, 138, 568–576. [Google Scholar] [CrossRef]
- Huang, N.; Angeles, E.R.; Domingo, J.; Magpantay, G.; Singh, S.; Zhang, G.; Kumaravadivel, N.; Bennett, J.; Khush, G.S. Pyramiding of bacterial blight resistance genes in rice: Marker-assisted selection using RFLP and PCR. Theor. Appl. Genet. 1997, 95, 313–320. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- McCouch, S.R.; CGSNL (Committee on Gene Symbolization, Nomenclature and Linkage, Rice Genetic Cooperative). Gene nomenclature system for rice. Rice 2008, 1, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Y.; Wu, Z.L.; Xing, Y.Y.; Zheng, F.G.; Guo, X.L.; Zhang, W.G.; Hong, M.M. Nucleotide sequence of rice waxy gene. Nucleic Acids Res. 1990, 18, 5898. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef]
- Wang, D.-Y.; Zhang, X.-F.; Zhu, Z.-W.; Chen, N.; Min, J.; Yao, Q.; Yan, J.-L.; Liao, X.-Y. Correlation analysis of rice grain quality characteristics. Acta Agron. Sin. 2005, 31, 1086–1091. (In Chinese) [Google Scholar]
- Weng, J.; Gu, S.; Wan, X.; Gao, H.; Guo, T.; Su, N.; Lei, C.; Zhang, X.; Cheng, Z.; Guo, X.; et al. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 2008, 18, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, C.; Xing, Y.; Yun, P.; Luo, L.; Yan, B.; Peng, B.; Xie, W.; Wang, G.; Li, X.; et al. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice. Nat. Genet. 2014, 46, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.H.; Tan, Y.F.; He, Y.Q.; Xu, C.G.; Zhang, Q. Simultaneous improvement for four quality traits of Zhenshan 97, an elite parent of hybrid rice, by molecular marker-assisted selection. Theor. Appl. Genet. 2003, 106, 326–331. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Population | Trait | MR | HR | GY | BRY | MRY | HRY |
|---|---|---|---|---|---|---|---|
| TI | BR | 0.764 ** | −0.157 | 0.229 ** | 0.292 ** | 0.287 ** | 0.107 |
| MR | 0.160 | 0.205 ** | 0.253 ** | 0.287 ** | 0.275 ** | ||
| HR | −0.064 | −0.074 | −0.046 | 0.505 ** | |||
| GY | 0.998 ** | 0.996 ** | 0.816 ** | ||||
| BRY | 0.998 ** | 0.809 ** | |||||
| MRY | 0.826 ** | ||||||
| ZM | BR | 0.815 ** | 0.148 | −0.006 | 0.055 | 0.054 | 0.063 |
| MR | 0.230 ** | 0.063 | 0.112 | 0.136 | 0.157 | ||
| HR | 0.354 ** | 0.362 ** | 0.368 ** | 0.710 ** | |||
| GY | 0.998 ** | 0.997 ** | 0.902 ** | ||||
| BRY | 0.999 ** | 0.905 ** | |||||
| MRY | 0.907 ** | ||||||
| XM | BR | 0.784 ** | 0.233 ** | 0.246 ** | 0.292 ** | 0.296 ** | 0.300 ** |
| MR | 0.363 ** | 0.365 ** | 0.398 ** | 0.425 ** | 0.449 ** | ||
| HR | 0.224 ** | 0.232 ** | 0.241 ** | 0.605 ** | |||
| GY | 0.999 ** | 0.998 ** | 0.900 ** | ||||
| BRY | 0.999 ** | 0.903 ** | |||||
| MRY | 0.907 ** |
| Chr | Interval | QTL | LOD | LOD (A) | LOD (ge) | A | ge | R2 (A) | R2 (ge) |
|---|---|---|---|---|---|---|---|---|---|
| 2 | RM6–RM240 | qGY2 | 8.84 | 8.25 | −1.89 | 10.75 | |||
| qBRY2 | 8.76 | 8.09 | −1.54 | 10.37 | |||||
| qMRY2 | 8.15 | 7.61 | −1.30 | 9.78 | |||||
| qHRY2 | 7.44 | 6.57 | −1.21 | 8.38 | |||||
| 3 | RM15139–RM15303 | qBR3.1 | 26.64 | 25.34 | 0.41 | 14.16 | |||
| qHR3 | 14.07 | 13.17 | −2.51 | 17.81 | |||||
| RM16048–RM16184 | qBR3.2 | 8.02 | 7.46 | 0.21 | 3.75 | ||||
| 4 | RM6992–RM349 | qGY4 | 4.90 | 4.88 | 1.44 | 6.23 | |||
| qBRY4 | 4.59 | 4.58 | 1.14 | 5.64 | |||||
| qMRY4 | 4.53 | 4.52 | 0.99 | 5.69 | |||||
| qHRY4 | 3.90 | 3.85 | 0.89 | 4.55 | |||||
| 5 | RM437–RM18189 | qBR5.1 | 35.75 | 34.08 | −0.51 | 20.20 | |||
| qMR5 | 11.22 | 10.51 | −0.39 | 11.99 | |||||
| qGY5 | 4.18 | 3.52 | −1.30 | 4.52 | |||||
| qBRY5 | 5.66 | 4.81 | −1.25 | 6.02 | |||||
| qMRY5 | 6.17 | 5.06 | −1.13 | 6.48 | |||||
| RM274–RM334 | qBR5.2 | 3.11 | 2.93 | −0.13 | 1.38 | ||||
| 6 | RM190–RM587 | qBR6 | 15.18 | 14.32 | −0.30 | 7.45 | |||
| qMR6 | 6.30 | 5.96 | −0.28 | 6.44 | |||||
| qHR6 | 10.08 | 4.35 | 5.73 | −1.40 | −1.20 | 5.59 | 4.28 | ||
| 7 | RM70–RM18 | qBR7 | 5.82 | 5.54 | 0.19 | 2.75 | |||
| 8 | RM547–RM22755 | qBR8 | 8.00 | 6.82 | −0.20 | 3.31 | |||
| qGY8 | 2.85 | 2.82 | −1.10 | 3.55 | |||||
| qBRY8 | 3.42 | 3.33 | −0.98 | 4.09 | |||||
| qMRY8 | 2.85 | 2.82 | −0.79 | 3.53 | |||||
| 10 | RM6100–RM3773 | qBR10 | 14.93 | 12.44 | −0.28 | 6.60 | |||
| 12 | RM20–RM27610 | qMR12 | 3.48 | 3.36 | 0.21 | 3.50 |
| Chr | Interval | QTL | LOD | LOD (A) | LOD (ge) | A | ge | R2 (A) | R2 (ge) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | RG532–RM5359 | qHR1 | 9.34 | 9.14 | 2.21 | 8.95 | |||
| qGY1.1 | 5.25 | 4.35 | 0.89 | 4.79 | |||||
| qBRY1.1 | 5.53 | 4.56 | 0.74 | 4.93 | |||||
| qMRY1.1 | 5.68 | 4.73 | 0.69 | 5.01 | |||||
| qHRY1.1 | 6.16 | 4.60 | 0.70 | 4.59 | |||||
| RZ730–RG381 | qBR1 | 7.34 | 6.67 | −0.24 | 5.91 | ||||
| qGY1.2 | 6.63 | 6.61 | 1.12 | 7.51 | |||||
| qBRY1.2 | 5.96 | 5.93 | 0.86 | 6.59 | |||||
| qMRY1.2 | 5.58 | 5.58 | 0.76 | 6.02 | |||||
| qHRY1.2 | 4.69 | 4.30 | 0.67 | 4.34 | |||||
| 2 | A5–RM71 | qHR2 | 3.30 | 3.30 | −1.29 | 3.00 | |||
| 3 | RM251–RG393 | qBR3 | 6.00 | 5.77 | −0.22 | 5.02 | |||
| RZ613–RG418A | qGY3 | 4.17 | 3.14 | −0.76 | 3.42 | ||||
| qBRY3 | 4.17 | 3.04 | −0.61 | 3.24 | |||||
| qMRY3 | 4.00 | 2.98 | −0.56 | 3.18 | |||||
| qHRY3 | 5.06 | 4.08 | −0.65 | 4.03 | |||||
| 4 | RZ69–RM3317 | qHRY4 | 5.72 | 5.68 | 0.76 | 5.52 | |||
| RM401–RM3643 | qHR4 | 3.76 | 3.51 | 1.31 | 3.20 | ||||
| 5 | CDO348–RG480 | qHR5 | 4.95 | 4.94 | −1.61 | 4.69 | |||
| qHRY5 | 5.80 | 5.79 | −0.79 | 5.81 | |||||
| 6 | RZ516–RM197 | qHR6 | 5.72 | 4.78 | −1.56 | 4.51 | |||
| qGY6 | 7.97 | 3.30 | 4.67 | −0.79 | 0.80 | 3.72 | 3.72 | ||
| qBRY6 | 7.74 | 3.22 | 4.52 | −0.63 | 0.63 | 3.54 | 3.54 | ||
| qMRY6 | 7.38 | 3.13 | 4.24 | −0.58 | 0.54 | 3.39 | 3.03 | ||
| qHRY6 | 7.41 | 4.15 | 3.27 | −0.66 | −0.64 | 4.11 | 3.89 | ||
| RM276–RZ667 | qMR6 | 3.16 | 3.01 | 0.20 | 3.25 | ||||
| 7 | RG650–RZ395 | qBR7 | 4.08 | 3.50 | 0.18 | 3.25 | |||
| 9 | RG667–RM201 | qMRY9 | 4.13 | 3.59 | 0.60 | 3.77 | |||
| 10 | RZ811–RZ583 | qGY10 | 4.46 | 3.68 | −0.83 | 4.13 | |||
| qBRY10 | 5.21 | 4.34 | −0.73 | 4.78 | |||||
| qMRY10 | 5.17 | 4.45 | −0.68 | 4.82 | |||||
| 11 | RZ816–RM332 | qBR11.1 | 3.12 | 3.10 | 0.16 | 2.63 | |||
| RM187–RM254 | qBR11.2 | 3.20 | 3.18 | −0.16 | 2.73 |
| Chr | Interval | QTL | LOD | LOD (A) | A | R2 (A) |
|---|---|---|---|---|---|---|
| 3 | RM6849–RM14629 | qBR3.1 | 3.97 | 3.58 | 0.21 | 4.57 |
| qMR3.1 | 3.76 | 3.01 | 0.28 | 2.96 | ||
| RZ696–RG445A | qBR3.2 | 3.14 | 2.99 | −0.19 | 3.58 | |
| RZ519–RZ328 | qMR3.2 | 3.35 | 3.26 | −0.28 | 3.10 | |
| RM85–RG418A | qHR3 | 3.39 | 3.12 | −1.61 | 4.28 | |
| 5 | RM13–RM267 | qBR5 | 3.94 | 3.02 | 0.19 | 3.53 |
| RG182–RG413 | qMR5 | 6.86 | 6.86 | 0.43 | 7.05 | |
| RM163–RG470 | qHR5 | 3.78 | 3.28 | −1.62 | 4.58 | |
| qHRY5 | 3.02 | 3.00 | −0.80 | 3.09 | ||
| 6 | RM190–RM204 | qGY6 | 7.14 | 6.64 | −1.74 | 8.46 |
| qBRY6 | 7.14 | 6.65 | −1.43 | 8.25 | ||
| qMRY6 | 7.31 | 6.73 | −1.32 | 8.53 | ||
| qHRY6 | 4.97 | 4.30 | −0.99 | 4.85 | ||
| 10 | RM1859–RM184 | qGY10 | 4.56 | 4.06 | −1.36 | 5.15 |
| qBRY10 | 4.51 | 4.00 | −1.11 | 4.96 | ||
| qMRY10 | 4.70 | 4.17 | −1.04 | 5.28 | ||
| qHRY10 | 6.11 | 5.39 | −1.11 | 6.05 |
| Chr | Interval | QTL | LOD | A | D | R2 (%) |
|---|---|---|---|---|---|---|
| 2 | Tw31911–Tw32437 | qGY2 | 8.93 | 1.87 | 0.41 | 11.60 |
| qBRY2 | 8.88 | 1.52 | 0.29 | 11.59 | ||
| qMRY2 | 9.25 | 1.40 | 0.20 | 12.03 | ||
| qHRY2 | 7.35 | 1.29 | 0.16 | 10.67 | ||
| 3 | RM14302–RM14383 | qBR3 | 2.34 | −0.13 | 0.05 | 2.82 |
| qMR3 | 6.41 | −0.31 | 0.08 | 7.30 | ||
| 5 | RM3321–RM274 | qBR5 | 4.88 | −0.17 | −0.18 | 6.25 |
| qMR5 | 5.92 | −0.26 | −0.33 | 6.54 | ||
| 6 | RM549 | qHR6 | 8.05 | 1.34 | 0.31 | 6.01 |
| 7 | RM10–RM70 | qBR7 | 7.23 | 0.23 | −0.09 | 9.28 |
| qMR7 | 12.89 | 0.44 | −0.15 | 15.51 | ||
| 9 | RM219–RM1896 | qMR9 | 3.06 | 0.20 | 0.12 | 3.34 |
| 9 | RM107 | qGY9 | 2.83 | −0.93 | −0.84 | 3.35 |
| qBRY9 | 2.81 | −0.76 | −0.65 | 3.35 | ||
| qMRY9 | 3.09 | −0.73 | −0.52 | 3.68 | ||
| qHRY9 | 2.04 | −0.62 | −0.28 | 2.67 | ||
| 10 | RM6704–RM7300 | qBR10 | 2.20 | −0.10 | −0.21 | 2.52 |
| qMR10 | 2.92 | −0.18 | −0.24 | 3.29 | ||
| 11 | RM167–RM287 | qBR11 | 2.20 | 0.10 | 0.20 | 2.52 |
| qGY11 | 3.30 | 0.76 | 3.05 | 7.80 | ||
| qBRY11 | 3.48 | 0.63 | 2.52 | 8.00 | ||
| qMRY11 | 3.46 | 0.55 | 2.30 | 8.03 | ||
| qHRY11 | 2.43 | 0.36 | 2.34 | 7.19 | ||
| 12 | Tv963–RM3246 | qBR12 | 2.84 | 0.10 | −0.57 | 9.99 |
| qMR12 | 3.74 | 0.12 | −0.88 | 10.01 | ||
| qHRY12 | 2.04 | 0.62 | −0.32 | 2.73 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zhu, Y.-J.; Zhu, A.-D.; Fan, Y.-Y.; Huang, T.-X.; Zhang, J.-F.; Xie, H.-A.; Zhuang, J.-Y. Identification and Verification of Quantitative Trait Loci Affecting Milling Yield of Rice. Agronomy 2020, 10, 75. https://doi.org/10.3390/agronomy10010075
Zhang H, Zhu Y-J, Zhu A-D, Fan Y-Y, Huang T-X, Zhang J-F, Xie H-A, Zhuang J-Y. Identification and Verification of Quantitative Trait Loci Affecting Milling Yield of Rice. Agronomy. 2020; 10(1):75. https://doi.org/10.3390/agronomy10010075
Chicago/Turabian StyleZhang, Hui, Yu-Jun Zhu, An-Dong Zhu, Ye-Yang Fan, Ting-Xu Huang, Jian-Fu Zhang, Hua-An Xie, and Jie-Yun Zhuang. 2020. "Identification and Verification of Quantitative Trait Loci Affecting Milling Yield of Rice" Agronomy 10, no. 1: 75. https://doi.org/10.3390/agronomy10010075
APA StyleZhang, H., Zhu, Y. -J., Zhu, A. -D., Fan, Y. -Y., Huang, T. -X., Zhang, J. -F., Xie, H. -A., & Zhuang, J. -Y. (2020). Identification and Verification of Quantitative Trait Loci Affecting Milling Yield of Rice. Agronomy, 10(1), 75. https://doi.org/10.3390/agronomy10010075