1. Introduction
Reduction in rainfall is amongst the major climate change manifestation phenomena, and will have a significant impact on water resource availability in the grassland [
1,
2]. One of the major challenges for ecologists today is assessing mechanisms that determine ecosystems recovery and response as well as ecological changes that come as a result of climatic changes and frequent extreme climate [
3]. However, the response of the rangelands to the rainfall reduction is highly variable and ranges from minimal level impacts [
4,
5,
6] to prolonged change in ecosystem function structure and recovery [
7,
8,
9,
10]. The impact of reduced rainfall and its extremes has a potential to alter the ecosystem function depending on its influence on botanical composition, abundance of the species impacted [
11] and might even lead to soil erosion due to loss of vegetation cover.
Moreover, increase in precipitation variability is generally related to an increase in rainfall extremes and the frequencies of drought events [
12]. The availability of water is among the most critical factors that regulate the biological activities in arid and semi-arid ecosystems [
13]. Precipitation dictates to a large extent the availability of soil moisture and species abundance in arid and semi-arid ecosystems [
14,
15]. Harvesting by grazing animals is a common occurrence in the grassland ecosystem [
16]. The intensity of harvesting and resting period can impact the ability of certain species to compete and survive under reduced rainfall condition. These disturbances could exacerbate the negative impacts of environmental stress on plant production [
17].
Some studies have shown that resistance strategies have been developed by certain plant species to maintain their reproductive capabilities and stability [
18]. Some plants have tolerance strategies to resist combinations of defoliation and environmental stress [
19]. Indigenous grass species are often the most dominant plants because of their superior adaptation to stress and extreme conditions such as water stress and extensive grazing and thus preserve stability and productivity of the rangeland in semi-arid environments [
20]. These species are categorised grossly into those species that are abundant in good rangelands but decrease in abundance when the rangeland is undergrazed or overgrazed (decreasers), those that are abundant when the rangeland is overgrazed (increaser II) and those that are abundant when the rangeland is undergrazed (increaser I) [
21].
The increase in abundance of unpalatable functional groups, such as some increaser grass species and forbs, at the expense of palatable and taller grass species as a result of overgrazing and extreme weather events such as reduced rainfall are often used as an indication of land degradation [
22]. However, increases in these unpalatable functional groups may not be completely unnecessary to the functioning of the rangeland as they still provide soil cover [
23]. For example, forbs are high quality food for feeding of browsers in South African rangelands [
24]. However, the response of forbs to common drivers of vegetation dynamics such as grazing and rainfall variability remain under studied [
23].
Therefore, it is very important to know how grasslands are going to respond to decreasing rainfall that is expected in Southern African region and how the extent of reduction in rainfall and grazing by animals will affect the ecosystem species dynamics and biomass productivity of the system and state transitions. This can be done through monitoring the aboveground biomass yield, in particular by focusing on the dynamics of grass and forbs composition and abundance under future anticipated rainfall reduction climate scenarios in South Africa [
13]. The objective of this study was to investigate the impact of simulated rainfall reduction/deficit and resting periods on seasonal change in herbaceous species vegetation structure and aboveground biomass production of natural grassland in the semi-arid areas of South Africa.
4. Discussion
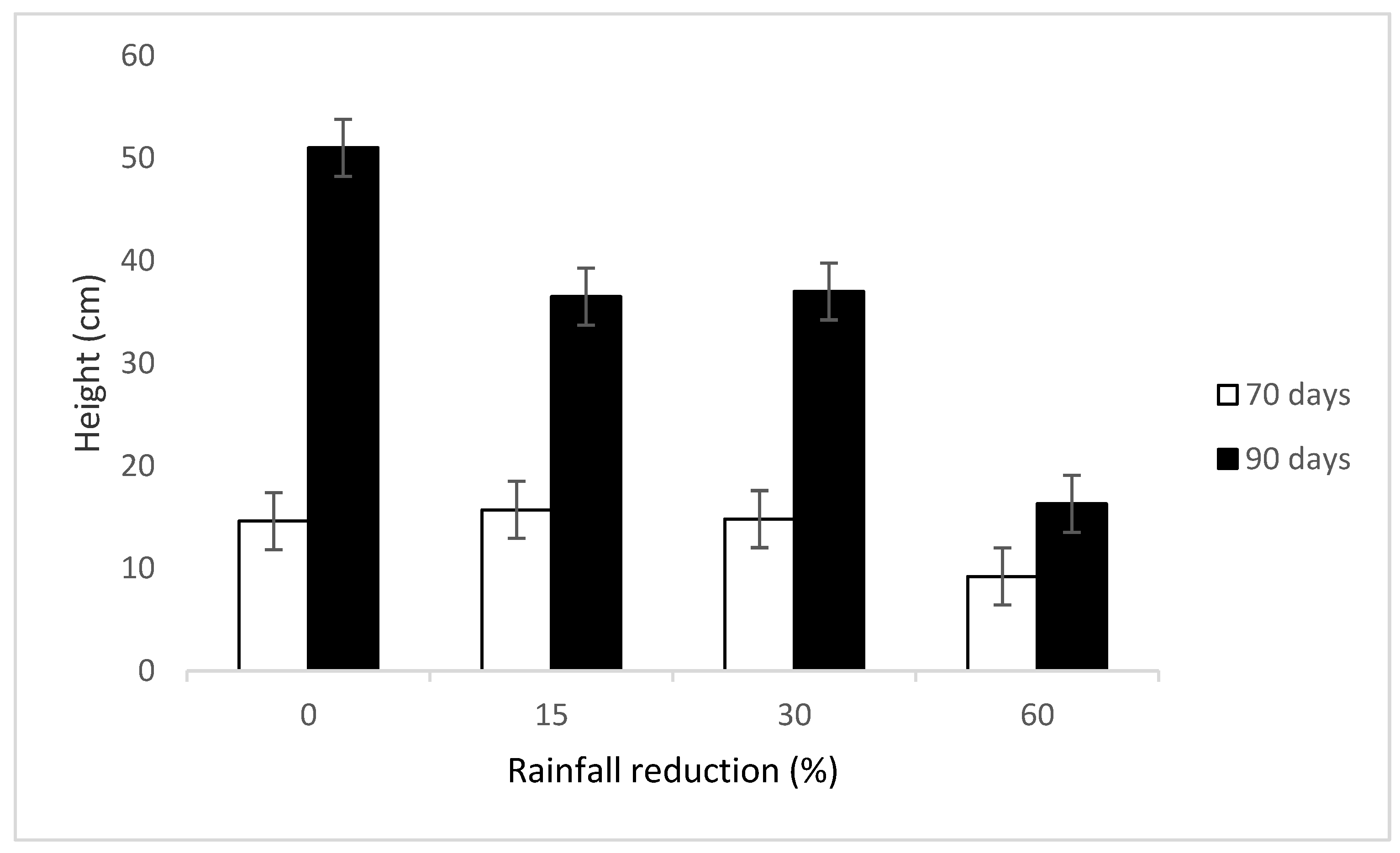
Our results showed that the vegetation height coincides with the vegetation biomass yield, which suggests that there is a positive relationship between vegetation height and the biomass production. The vegetation was taller at lower rainfall reduction treatments (0% and 15%) than the higher rainfall reduction treatments (30% and 60%). The heights were also affected by the seasons as the season with higher rainfall had taller vegetation (
Table 3). Vegetation height and biomass are very much influenced by available resources such as nutrients and water [
31]. These results concur with those of [
32,
33], who reported that vegetation growth mainly depends upon summer rainfalls, as the rainfall is generally lower in other seasons due to skewed distribution, particularly in summer rainfall areas.
Moreover, resting period affected vegetation height in different ways in different seasons. In spring, the vegetation was taller at the 90 days resting period rather than the 70 days one, while the opposite was true in autumn in both years. This is due to the fact that in spring the extra 20 days for the 90 days resting period gave this resting period extra time to grow. On the other hand, the 70 days resting period had a taller vegetation than the 90 days resting period in autumn in both years. This could be due to the fact that the 70 days resting period received rainfall just after it was cut and it was cut while getting summer rainfall and before the 90 days resting period was cut in summer. This gave an advantage to the 70 days resting period as the rainfall starts to decline in autumn and some annual plants begin to die. Height also correlated well with biomass in both years. Resting for 90 days in spring improved grass biomass production while resting for 70 days improved grass biomass yield in autumn in both years. The higher biomass yield in the 70 days resting period in autumn is attributed to the 20 day delay for 90 days cutting in December as the rainfalls are beginning to decrease while the 70 days resting period gets the summer rainfall (
Figure 1).
Grass harvest sampling every 70 days and 90 days over three seasons over a period of two years was able to show seasonal differences in terms of biomass production. This was also reported by [
33,
34]. In this study, shorter resting periods improved grass biomass production in autumn while in spring longer resting periods improved grass biomass production. The 0%, 15% and 30% rainfall reduction treatments had higher grass biomass production than the 60% rainfall reduction. According to [
35], the dominant grasses such as
D. eriantha and
E. curvula at different treatments appear to contribute more biomass production than the less dominant grass species such as
A. conaesta and
M. repens, hence these grasses will determine whether the rangeland will be productive for animal production or not.
Rainfall reduction significantly reduced overall biomass production only at 60% rainfall reduction. This clearly shows that the grasslands were able to show resilience by making adjustments in terms of species composition up to the level of 30% reduction in rainfall, however severe reduction in rainfall amount (60%) will have a significantly negative impact on rangeland productivity, which will lead to less biomass production and subsequently fodder for animal production (
Figure 5). This basically means that in 2016/17 the rainfall was better and hence it was possible to improve resilience of the system or support better growth by modifying the resting period (changing from 75 to 90 days), but such benefit will not be sustained with modification of resting period once the rainfall reduction becomes severe.
The native pasture biomass production was not as such affected by up to 30% reduction in rainfall due to resilience of increaser species such as
E. curvula that dominated the 15% and 30% rainfall reduction plots (
Figure 7 and
Figure 8). In a study that was conducted by [
36], it was found that increasers are more tolerant to grazing than decreasers. In that study, three increaser grass species had a higher compensation regrowth after four episodes of clipping. As observed in our study,
E. curvula is able to regrow after it has been harvested, even at high frequencies.
There is the expectation that a reduction in rainfall associated with climate change will have a negative impact on natural grazing land in arid and semi-arid areas of sub-Saharan Africa [
37]. This is mainly based on the fact that forage production in these areas is sensitive to rainfall reduction as it is the case in our study site. This also concurs with the results that were reported by [
38], who reported that the rangeland in the Sub-Saharan Africa is sensitive to climate change. In general, as expected season had an impact on overall biomass production as the summer season had a higher biomass production than spring and autumn in both years. Here, higher temperature and rainfall is the main driving force for the higher biomass production recorded in summer than in spring and autumn. This agrees with other studies that reported similar results pertaining the response of grassland biomass production to simulated climate change [
9,
35,
39]. The cumulative biomass yield was significantly higher in 2016/17 than 2017/18 because of the higher rainfall amount received in 2016/17. This means that with the expected decrease in rainfall associated with climate change and subsequently decrease in biomass accumulation, the farmers need to consider to supplement the animals and/or destock where possible in the event of severe rainfall reduction encounters.
The decreasers are resilient to up to 15% reduction in rainfall thereafter, however they are more prone to rainfall reduction than the increaser grass species as they were dominant and performed much better at 0% and 15% rainfall reduction plots while increasers were better performers at 30% and relatively even at 60% rainfall reductions (
Table 6 and
Table 7). This implies that continued rainfall reduction will potentially lead to disappearance of decreaser grass species such as
D. eriantha and
T. triandra in favour of increaser grass species such as
E. curvula and
C. dactylon and possibly forbs. In this study, the increaser grass species were more resilient to severe reduction in rainfall (as judged from increase in rainfall reduction) than decreaser grass species. The results from this study concur with the findings reported by [
40], who reported that decreaser grass species fluctuate with variation in soil moisture while the increaser grass species were more resilient to water deficiency than decreaser grass species. This means that decreasers require more water to produce the same unit of biomass than increaser grass species or forbs that require less water. However, [
41] reported that vegetation changes induced by drought may shift vegetation from grass-dominated to forb dominated community. This is in agreement with the finding of this study where grasses had a higher biomass production at 0%, 15% and 30% rainfall reduction while forbs had higher biomass production at 60% rainfall reduction and dominated the 30% and 60% rain reduction plots. This could be partly related to the failure of grasses to produce seeds and grasses having shorter-lived seeds than dominant forbs [
41], that have heavy seeds. Differences in the root systems of the grasses and forbs could also explain why forbs are dominant in low rainfall treatments. The deep root system of the forbs allows the forbs to access water and nutrients from the deeper soil layers, while on the other hand grasses have a shallow root system, hence they are not able to access water from the deeper soil layers [
42]. In addition, in low rainfall reduction treatments, i.e., 0% and 15% RD, grasses may likely over shade and outcompete the smaller forbs for resources such as light, soil nutrients and soil water [
43].
The rainfall reduction effect not only affects the aboveground biomass production and height but also modifies the species richness, diversity and functional group richness [
31]. It can be said that the deep root system of the forbs enables the competitive dominance of these species over grasses under moisture deficit conditions [
44,
45,
46]. In this study, the forbs are outcompeted by grasses at 0% and 15% as the grasses dominated these plots because of adequate precipitation that was available to support the growth of the decreaser grasses that dominated at 0% and 15% rainfall reduction and increaser grasses that dominated the 30% rainfall reduction treatments. The decline in grass production with rainfall reduction leads to a gradual build up in forb production, while the decline in forbs populations leads to grass build up [
44]. Forbs perform better when there is weak grass competition while grasses are more responsive to precipitation. The differences in environmental favorability between grasses and forbs is as a result of the competition [
44].
Our results show that the longer resting periods in autumn resulted in less grass biomass production, which ultimately resulted in less overall aboveground biomass production, while the effect of resting on the forbs biomass production was not consistent. The pattern of variation in seasonal precipitation had a strong significant effect on the pattern of overall biomass production in both years. The 2016/17 spring and summer seasons had lower rainfall than the 2017/18 spring and summer seasons, while the 2016/17 autumn season had a higher rainfall than the 2017/18 autumn season (
Figure 1). Hence, the vegetation height and biomass production of 2017/18 in spring and summer was higher compared to the same period in 2016/17, while in autumn of 2016/17 the biomass production was higher compared to 2017/18. These results concur with those reported by [
47], who reported that seasonal precipitation is an important driver of temporal dynamics of perennial grass richness. The overall biomass production was directly proportional to the rainfall amount received in each season as this was reflected through the biomass production where the overall biomass production was lower in the spring and autumn seasons of 2016/17 than the summer season in both years.
The results observed in this study could also suggest that reduction in precipitation is not the only factor that plays a major role in terms of biomass production but its interaction with resting period and the season in which the precipitation reduction occurs also matter a lot in affecting biomass production [
47,
48,
49,
50,
51]. In this study, the summer season had the highest amount of precipitation in both years (2016/17 and 2017/18) and had the highest biomass production compared to the other two seasons (spring and autumn). The resting period of 90 days in the spring season in both years benefited plants by providing them enough time to recover from the physiological shock that might have been caused by cutting at the beginning of the spring season after winter and the low temperature of the winter season. The higher biomass at the 90 days resting period can be also partly attributed to grass flowering that occurred after the 70 days resting period as the flowering parts also add to biomass [
52]. The biomass production was higher in summer than the spring and autumn seasons, which is because of the high summer rainfall during summer season in both years.
Overall biomass production decreases with the increase in rainfall reduction. This trend agrees with the trend reported by [
40], who reported higher biomass production in plots that did not experience rainfall reduction than the ones that experienced severe rainfall reduction treatments. Seasons alter biomass production and hence it is important that interpretation of the response take seasons into consideration in order to capture these differences that occur in different seasons. This is supported by the variability of the drought effect observed from season to season elsewhere [
53,
54]. Annual seasonal rainfall patterns are the main drivers of rangeland ecosystem and its duration determines biomass yield [
50]. This reduction in rainfall is also accompanied by an increase in monthly average temperature which exceeded the 30 years monthly average temperatures of the study site for both years (
Figure 2).
The rain use efficiency (RUE) was higher at the 90 days resting period in spring at all plots than the 70 days resting period in 2016/17, while in summer of both years the highest RUE was recorded for plots with the 70 days resting period rather than the 90 days one. Our results showed that biomass production is directly proportional to rain use efficiency as the grasses with higher RUE produced higher biomass. The RUE can also be used as a tool to determine biomass production. These results agree with those reported by [
55], who reported that water use efficiency coincides with the pattern of biomass production. This study was limited to biomass production as it is the harvestable biomass yield that is of primary interest for animal production.
The PCA showed that decreaser grass species that were dominated by
D. eriantha were more abundant at 0% RD and 15% RD in both years. They are less competitive as compared to the increaser species dominated by
E. curvula under low rainfall conditions. This implies that decreaser grass species require more water to produce more biomass (
Figure 7 and
Figure 8). In contrast,
E. curvula, which is an increaser II grass species, was more dominant at 30% and 60% RD in 2017/18. Moreover, forbs produced more biomass at 60% RD, which means that they are able to better tolerate water stress compared to grasses (
Figure 7 and
Figure 8). Forbs can also be an important source of forage in overgrazed and water stressed environments [
43].










 ) indicate the direction of the relationship between vegetation parameters and treatments. The longer the vector radiating from the centre, the stronger the effects of the resting period and the resting period and the closer the dot to the vector the stronger the relationship between species and soil property. V1 = 0% RD, 70 RP, V2 = 0% RD, 90 RP, V3 = 15% RD, 70 RP, V4 = 15% RD, 90 RP, V5 = 30% RD, 70 RP, V6 = 30% RD, 90 RP, V7 = 60% RD, 70 RP, V8 = 60% RD, 90 RP. Cd = Cynodon dactylon, Die = Digitaria eriantha, Erc = Eragrostis curvula, Tht = Themeda triandra, Erl = Eragrostis lehmaniana, Spf = Sporobolus fimbriatus, Hec = Heteropogon contortus, Sen = Setaria nigrirostris, Ses = Setaria sphacelata, Cye = Cymbopogon excavatus, Umos = Urochloa mosambicensis, Erb = Eragrostis barbinodes, Arc = Aristida congesta, Erpl = Eragrostis plana, Brs = Brachiaria serata, Hyh = Hypperhenia hirta, Mer = Melinis repens Frbs = Forbs, Dec = Decreasers, Inc = Increasers.
) indicate the direction of the relationship between vegetation parameters and treatments. The longer the vector radiating from the centre, the stronger the effects of the resting period and the resting period and the closer the dot to the vector the stronger the relationship between species and soil property. V1 = 0% RD, 70 RP, V2 = 0% RD, 90 RP, V3 = 15% RD, 70 RP, V4 = 15% RD, 90 RP, V5 = 30% RD, 70 RP, V6 = 30% RD, 90 RP, V7 = 60% RD, 70 RP, V8 = 60% RD, 90 RP. Cd = Cynodon dactylon, Die = Digitaria eriantha, Erc = Eragrostis curvula, Tht = Themeda triandra, Erl = Eragrostis lehmaniana, Spf = Sporobolus fimbriatus, Hec = Heteropogon contortus, Sen = Setaria nigrirostris, Ses = Setaria sphacelata, Cye = Cymbopogon excavatus, Umos = Urochloa mosambicensis, Erb = Eragrostis barbinodes, Arc = Aristida congesta, Erpl = Eragrostis plana, Brs = Brachiaria serata, Hyh = Hypperhenia hirta, Mer = Melinis repens Frbs = Forbs, Dec = Decreasers, Inc = Increasers.

 ) indicate the direction of the relationship between vegetation parameters and treatments. The longer the vector radiating from the centre, the stronger the effects of the resting period and the resting period and the closer the dot to the vector the stronger the relationship between species and soil property. V1 = 0% RD, 70 RP, V2 = 0% RD, 90 RP, V3 = 15% RD, 70 RP, V4 = 15% RD, 90 RP, V5 = 30% RD, 70 RP, V6 = 30% RD, 90 RP, V7 = 60% RD, 70 RP, V8 = 60% RD, 90 RP. Cd= Cynodon dactylon, Die = Digitaria eriantha, Erc = Eragrostis curvula, Tht = Themeda triandra, Erl = Eragrostis lehmaniana, Spf = Sporobolus fimbriatus, Hec = Heteropogon contortus, Sen = Setaria nigrirostris, Ses = Setaria sphacelata, Cye = Cymbopogon excavatus, Umos = Urochloa mosambicensis, Erb = Eragrostis barbinodes, Arc = Aristida congesta, Erpl = Eragrostis plana, Brs = Brachiaria serata, Hyh = Hypperhenia hirta, Mer = Melinis repens, Hei = Height, Ob = Overall biomass, Grss = Grass, Frbs = Forbs, Dec = Decreasers, Inc = Increasers, RUE = Rain use efficiency.
) indicate the direction of the relationship between vegetation parameters and treatments. The longer the vector radiating from the centre, the stronger the effects of the resting period and the resting period and the closer the dot to the vector the stronger the relationship between species and soil property. V1 = 0% RD, 70 RP, V2 = 0% RD, 90 RP, V3 = 15% RD, 70 RP, V4 = 15% RD, 90 RP, V5 = 30% RD, 70 RP, V6 = 30% RD, 90 RP, V7 = 60% RD, 70 RP, V8 = 60% RD, 90 RP. Cd= Cynodon dactylon, Die = Digitaria eriantha, Erc = Eragrostis curvula, Tht = Themeda triandra, Erl = Eragrostis lehmaniana, Spf = Sporobolus fimbriatus, Hec = Heteropogon contortus, Sen = Setaria nigrirostris, Ses = Setaria sphacelata, Cye = Cymbopogon excavatus, Umos = Urochloa mosambicensis, Erb = Eragrostis barbinodes, Arc = Aristida congesta, Erpl = Eragrostis plana, Brs = Brachiaria serata, Hyh = Hypperhenia hirta, Mer = Melinis repens, Hei = Height, Ob = Overall biomass, Grss = Grass, Frbs = Forbs, Dec = Decreasers, Inc = Increasers, RUE = Rain use efficiency.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}