The Impact of Grazing on the Grass Composition in Temperate Grassland

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Field Measurements and Calculations
2.4. Soil Nutrient Contents
2.5. Soil Moisture and Temperature Measurements
2.6. Palatability Measurements
2.7. Cover and Species Richness
2.8. Data Analysis
3. Results
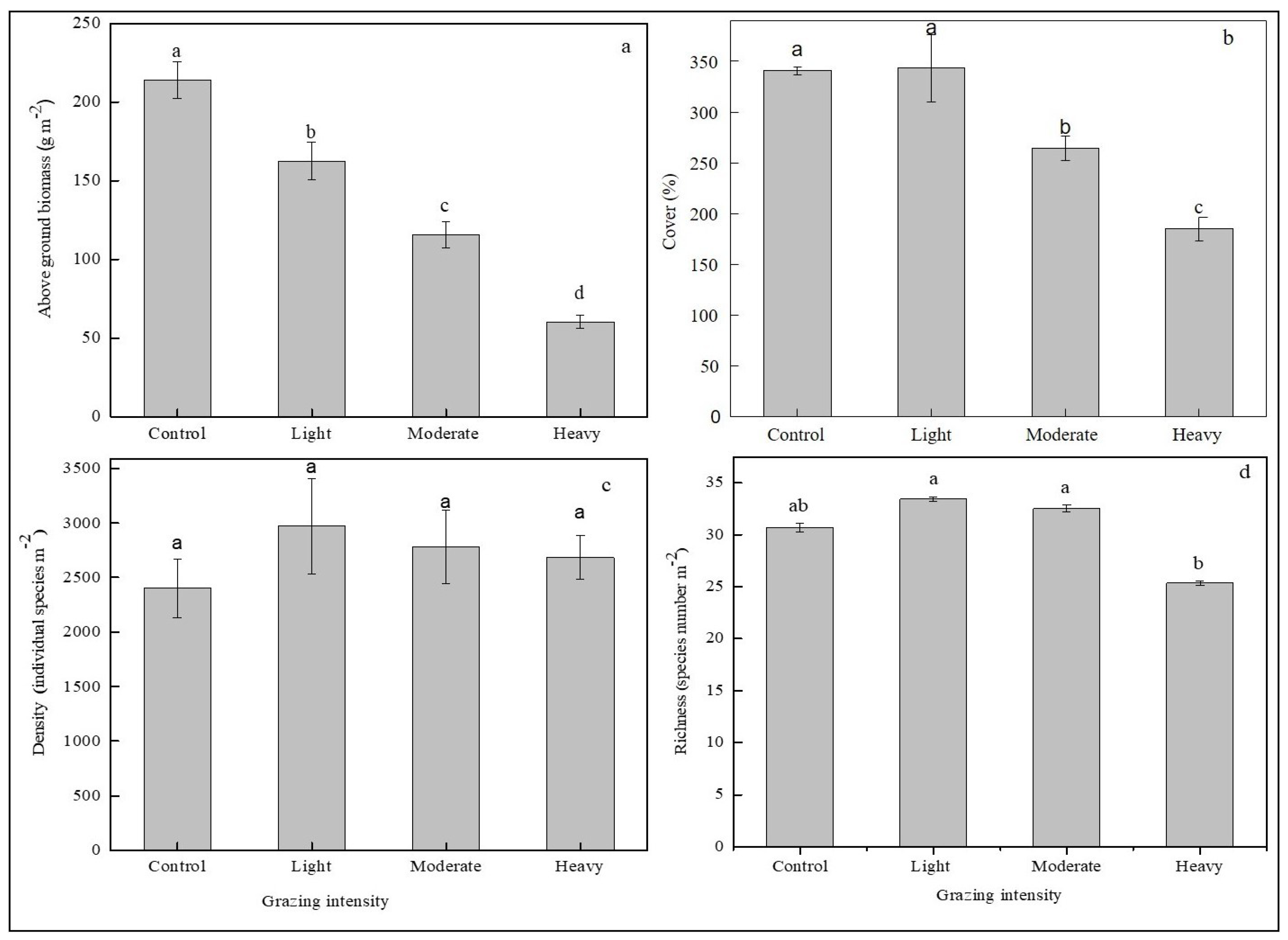
3.1. Grazing Impact on Community Characteristics
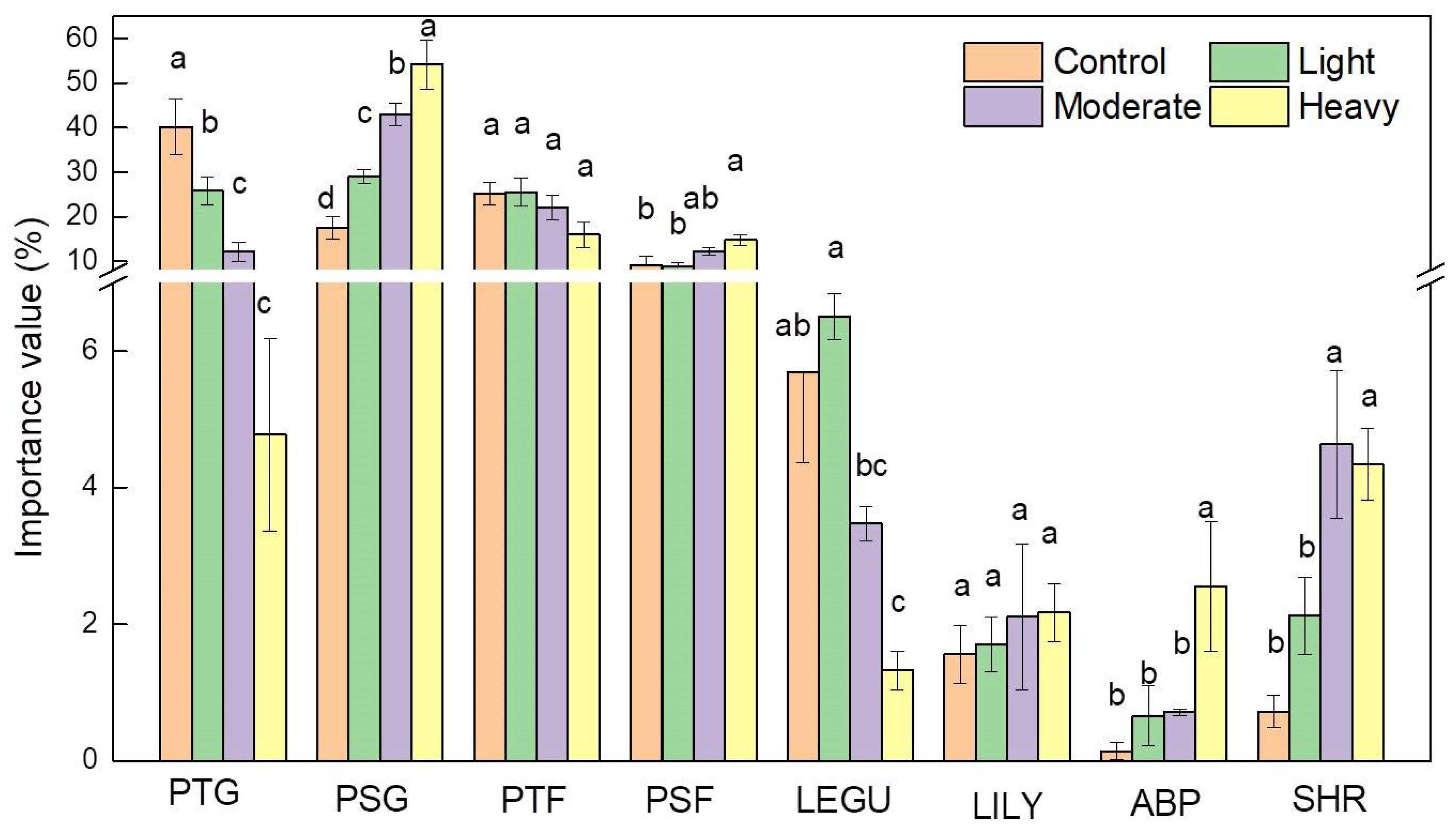
3.2. Response of Plant Functional Groups to Grazing Intensities
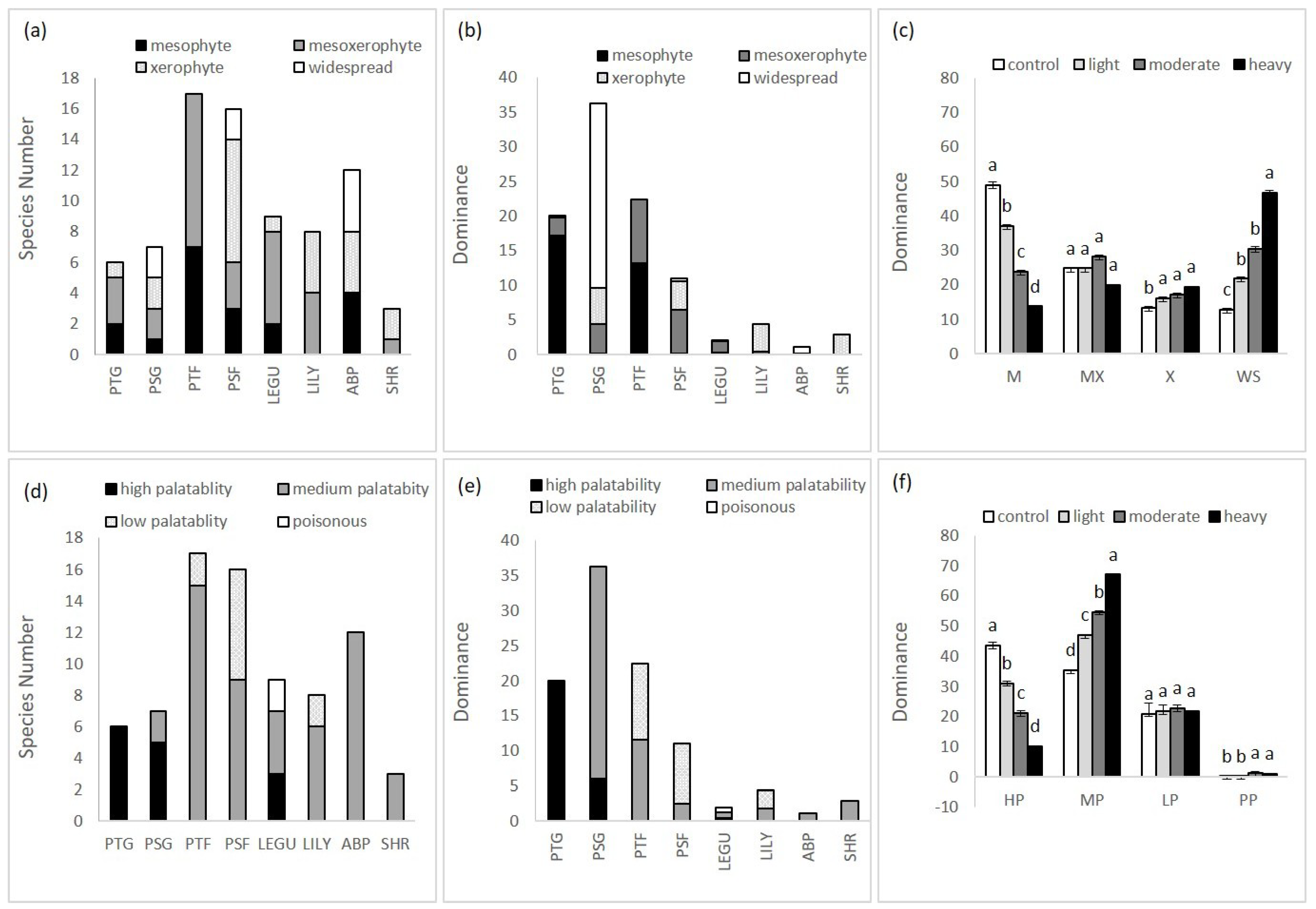
3.3. Change of Plant Species Functional Groups in Grazing Succession
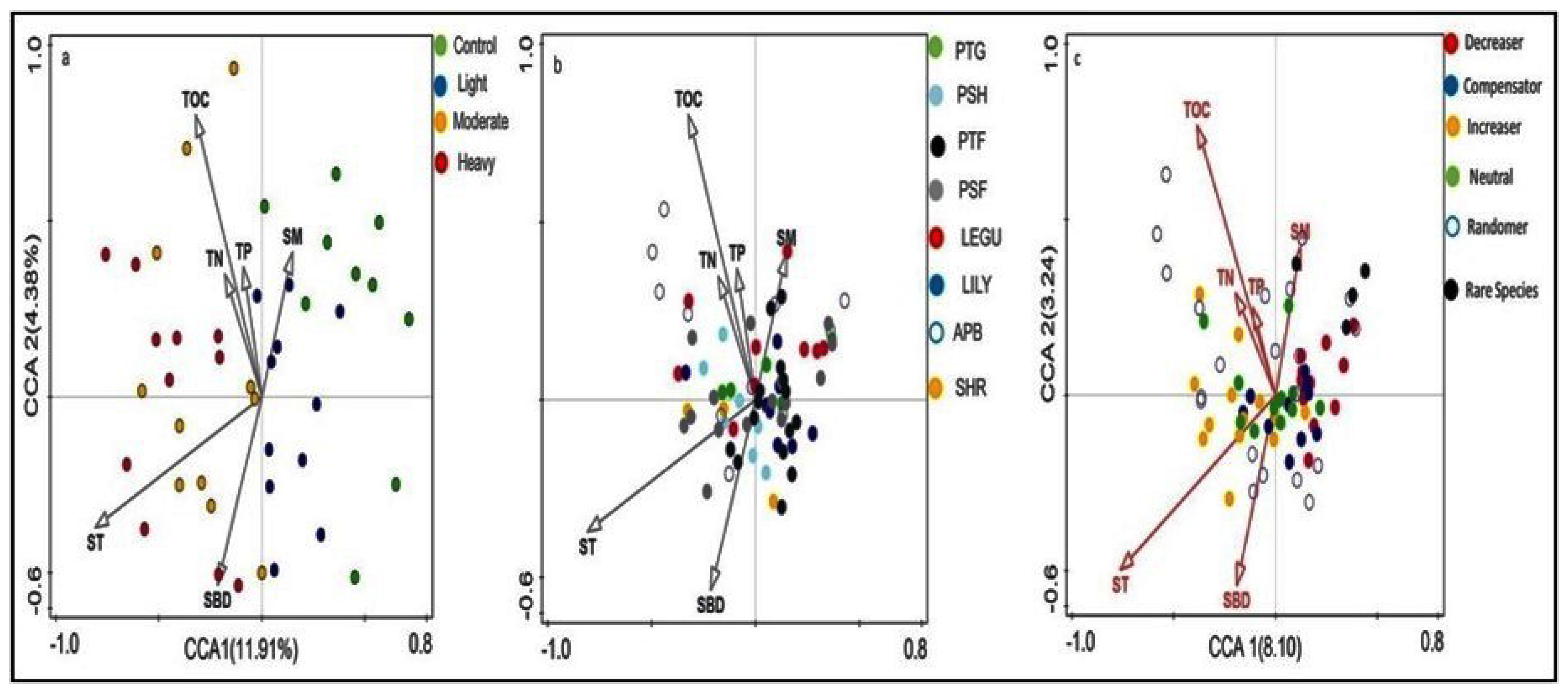
3.4. Relationship between Plant Functional Group and Soil Factors
4. Discussion
4.1. Grazing Impact on Community Characteristics
4.2. Response of Plant Functional Groups to Grazing Intensities
4.3. Change of Plant Species Functional Groups in Grazing Succession
4.4. Relationship between Plant Functional Group and Soil Factors
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- Christiansen, S.; Svejcar, T. Grazing effects on shoot and root dynamics and above- and below-ground non-structural carbohydrate in Caucasian bluestem. Grass Forage Sci. 1988, 43, 111–119. [Google Scholar] [CrossRef]
- Biondini, M.E.; Patton, B.D.; Nyren, P.E. Grazing intensity and ecosystem processes in a northern mixed-grass prairie, USA. Ecol. Appl. 1998, 8, 469–479. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative Effects of Grazing on Vegetation and Soils Over a Global Range of Environments. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]
- McNaughton, S.J.; Milchunas, D.G.; Frank, D.A. How can net Primary Productivity be Measured in Grazing Ecosystems? Ecology 1996, 77, 974–977. [Google Scholar] [CrossRef]
- Cingolani, A.M.; Noy-Meir, I.; Díaz, S. Grazing effects on rangeland diversity: A synthesis of contemporary models. Ecol. Appl. 2005, 15, 757–773. [Google Scholar] [CrossRef]
- Lavorel, S.; Díaz, S.; Cornelissen, J.H.C.; Garnier, E.; Harrison, S.P.; McIntyre, S.; Pausas, J.G.; Pérez-Harguindeguy, N.; Roumet, C.; Urcelay, C. Plant Functional Types: Are We Getting Any Closer to the Holy Grail? Springer-Verlag: Berlin, Germany, 2007; pp. 149–164. [Google Scholar]
- Hallett, L.; Stein, C.; Suding, K.N. Functional diversity increases ecological stability in a grazed grassland. Oecologia 2017, 183, 831–840. [Google Scholar] [CrossRef]
- Li, W.; Xu, F.; Zheng, S.; Taube, F.; Bai, Y. Patterns and thresholds of grazing-induced changes in community structure and ecosystem functioning: Species-level responses and the critical role of species traits. J. Appl. Ecol. 2016, 54, 963–975. [Google Scholar] [CrossRef]
- Oñatibia, G.R.; Boyero, L.; Aguiar, M.R. Regional productivity mediates the effects of grazing disturbance on plant cover and patch-size distribution in arid and semi-arid communities. Oikos 2018, 127, 1205–1215. [Google Scholar] [CrossRef]
- Proulx, M.; Mazumder, A. Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems. Ecology 1998, 79, 2581–2592. [Google Scholar] [CrossRef]
- Ganjurjav, H.; Gornish, E.S.; Hu, G.; Wan, Y.; Li, Y.; Danjiu, L.; Gao, Q. Temperature leads to annual changes of plant community composition in alpine grasslands on the Qinghai-Tibetan Plateau. Environ. Monit. Assess. 2018, 190, 585. [Google Scholar] [CrossRef]
- Katoh, K.; Takeuchi, K.; Jiang, D.; Nan, Y.; Kou, Z. Vegetation restoration by seasonal exclosure in the Kerqin Sandy Land, Inner Mongolia. Plant Ecol. 1998, 139, 133–144. [Google Scholar] [CrossRef]
- Yan, Y.; Lu, X. Is grazing exclusion effective in restoring vegetation in degraded alpine grasslands in Tibet, China? PeerJ 2015, 3, e1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renzhong, W.; A Ripley, E. Effects of grazing on a Leymus chinensis grassland on the Songnen plain of North-Eastern China. J. Arid. Environ. 1997, 36, 307–318. [Google Scholar] [CrossRef]
- Schönbach, P.; Wan, H.; Schiborra, A.; Gierus, M.; Bai, Y.; Müller, K.; Glindemann, T.; Wang, C.; Susenbeth, A.; Taube, F. Short-term management and stocking rate effects of grazing sheep on herbage quality and productivity of Inner Mongolia steppe. Crop. Pasture Sci. 2009, 60, 963–974. [Google Scholar] [CrossRef]
- Briske, D.D. Plant response to defoliation: Morphological considerations and allocation priorities. In Rangelands: A Resource under Siege, Proceedings of the 2nd International Rangeland Congress, Adelaide, Australia, 13–18 May 1984; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1986; pp. 425–427. [Google Scholar]
- Stuart-Hill, G.; Tainton, N.; Barnard, H. The influence of an Acacia karroo tree on grass production in its vicinity. J. Grassl. Soc. S. Afr. 1987, 4, 83–88. [Google Scholar] [CrossRef]
- Ma, L.; Yuan, F.; Liang, H.; Rong, Y. The effects of grazing management strategies on the vegetation, diet quality, intake and performance of free grazing sheep. Livest. Sci. 2014, 161, 185–192. [Google Scholar] [CrossRef]
- Patton, B.D.; Dong, X.; Nyren, P.E.; Nyren, A. Effects of Grazing Intensity, Precipitation, and Temperature on Forage Production. Rangel. Ecol. Manag. 2007, 60, 656–665. [Google Scholar] [CrossRef]
- McNaughton, S.J. Grazing as an Optimization Process: Grass-Ungulate Relationships in the Serengeti. Am. Nat. 1979, 113, 691–703. [Google Scholar] [CrossRef]
- Kent, M. Vegetation Description and Data Analysis: A Practical Approach, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; p. 428. [Google Scholar]
- Bao, S.D. Soil Agricultural Chemical Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000; pp. 265–267. [Google Scholar]
- Wang, Z.; Johnson, D.A.; Rong, Y.; Wang, K. Grazing effects on soil characteristics and vegetation of grassland in northern China. Solid Earth 2016, 7, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Peth, S.; Krümmelbein, J.; Horn, R.; Wang, Z.; Steffens, M.; Hoffmann, C.; Peng, X. Spatial variability of soil properties affected by grazing intensity in Inner Mongolia grassland. Ecol. Model. 2007, 205, 241–254. [Google Scholar] [CrossRef]
- Li, X.-Y.; Zhang, S.-Y.; Peng, H.-Y.; Hu, X.; Ma, Y.-J. Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: Results from Inner Mongolia steppe ecosystem of north China. Agric. For. Meteorol. 2013, 171, 20–30. [Google Scholar] [CrossRef]
- Larbi, A.; Osakwe, I.I.; Lambourne, J.W. Variation in relative palatability to sheep among Gliricidia sepium provenances. Agrofor. Syst. 1993, 22, 221–224. [Google Scholar] [CrossRef]
- Lambert, M.G.; Jung, G.A.; Fletcher, R.H.; Budding, P.J.; Costall, D.A. Forage shrubs in North Island hill country 2. Sheep and goat preferences. N. Z. J. Agric. Res. 1989, 32, 485–490. [Google Scholar] [CrossRef]
- Obour, R.; Oppong, S.K. Forage palatability of Broussonetia papyrifera an invasive species in Ghana: Relative preference and palatability by sheep and goats. J. Energy Nat. Resour. Manag. 2015, 2, 63–70. [Google Scholar] [CrossRef]
- Margalef, R. On Certain Unifying Principles in Ecology. Am. Nat. 1963, 97, 357–374. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version). 2002. Available online: www.canoco.com (accessed on 19 August 2020).
- Yoshioka, P. Misidentification of the Bray-Curtis similarity index. Mar. Ecol. Prog. Ser. 2008, 368, 309–310. [Google Scholar] [CrossRef] [Green Version]
- Milchunas, D.G.; Lauenroth, W.K.; Chapman, P.L.; Kazempour, M.K. Effects of grazing, topography, and precipitation on the structure of a semiarid grassland. Vegetatio 1989, 80, 11–23. [Google Scholar] [CrossRef]
- Na, Y.; Li, J.; Hoshino, B.; Bao, S.; Qin, F.; Myagmartseren, P. Effects of Different Grazing Systems on Aboveground Biomass and Plant Species Dominance in Typical Chinese and Mongolian Steppes. Sustainability 2018, 10, 4753. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, Q.; Jing, Z.; Wang, W. The effects of protective enclosure on vegetation diversity, and productivity of degraded alpine Kobresia meadow (Qinghai-Tibetan Plateau). Pol. J. Ecol. 2009, 57, 495–502. [Google Scholar]
- Warren, S.D.; Eldridge, D.J. Biological Soil Crusts and Livestock in Arid Ecosystems: Are They Compatible? In Biological Soil Crusts: Structure, Function, and Management; Belnap, J., Lange, O.L., Eds.; Springer: Berlin, Germany, 2001; Volume 150, pp. 401–415. [Google Scholar]
- Liu, H.; Han, X.; Li, L.; Huang, J.; Liu, H.; Li, X. Grazing Density Effects on Cover, Species Composition, and Nitrogen Fixation of Biological Soil Crust in an Inner Mongolia Steppe. Rangel. Ecol. Manag. 2009, 62, 321–327. [Google Scholar] [CrossRef]
- Yates, C.J.; Norton, D.A.; Hobbs, R.J. Grazing effects on plant cover, soil and microclimate in fragmented woodlands in south-western Australia: Implications for restoration. Austral Ecol. 2000, 25, 36–47. [Google Scholar] [CrossRef]
- Anderson, J.L. Exclosures on the Beaver Dam slope in Arizona and Utah. Rangelands 1994, 16, 184–188. [Google Scholar]
- McIntyre, S.; Lavorel, S. Livestock grazing in subtropical pastures: Steps in the analysis of attribute response and plant functional types. J. Ecol. 2001, 89, 209–226. [Google Scholar] [CrossRef]
- Vesk, P.A.; Westoby, M. Predicting plant species’ responses to grazing. J. Appl. Ecol. 2002, 38, 897–909. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Lavorel, S.; McIntyre, S.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.M.; Sternberg, M.; Noy-Meir, I.; et al. Plant trait responses to grazing ? a global synthesis. Glob. Chang. Boil. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- McIntyre, S.; Lavorel, S.; Landsberg, J.; Forbes, T. Disturbance response in vegetation–towards a global perspective on functional traits. J. Veg. Sci. 1999, 10, 621–630. [Google Scholar] [CrossRef]
- Jost, L.; Chao, A.; Chazdon, R.L. Compositional similarity and β (beta) diversity. In Biological Diversity: Frontiers in Measurement and Assessment, 1st ed.; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 66–84. [Google Scholar]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P.; Information, P.E.K.F.C. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Hao, M.; Corral-Rivas, J.J.; González-Elizondo, M.S.; Ganeshaiah, K.N.; Nava-Miranda, M.G.; Zhang, C.; Zhao, X.; Von Gadow, K. Assessing biological dissimilarities between five forest communities. For. Ecosyst. 2019, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Fleischner, T.L. Ecological Costs of Livestock Grazing in Western North America. Conserv. Boil. 1994, 8, 629–644. [Google Scholar] [CrossRef] [Green Version]
- Huntly, N. Herbivores and the dynamics of communities and ecosystems. Annu. Rev. Ecol. Syst. 1991, 22, 477–503. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Noy-Meir, I.; Cabido, M. Can grazing response of herbaceous plants be predicted from simple vegetative traits? J. Appl. Ecol. 2001, 38, 497–508. [Google Scholar] [CrossRef] [Green Version]
- Jauffret, S.; Lavorel, S. Are plant functional types relevant to describe degradation in arid, southern Tunisian steppes? J. Veg. Sci. 2003, 14, 399–408. [Google Scholar] [CrossRef]
- Navarro, T.; Alados, C.; Cabezudo, B. Changes in plant functional types in response to goat and sheep grazing in two semi-arid shrublands of SE Spain. J. Arid. Environ. 2006, 64, 298–322. [Google Scholar] [CrossRef]
- Rusch, G.M.; Pausas, J.G.; Lepš, J. Plant functional types in relation to disturbance and land use: Introduction. J. Veg. Sci. 2003, 14, 307–310. [Google Scholar] [CrossRef]
- Derner, J.D.; Lauenroth, W.K.; Stapp, P.; Augustine, D.J. Livestock as Ecosystem Engineers for Grassland Bird Habitat in the Western Great Plains of North America. Rangel. Ecol. Manag. 2009, 62, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E. The effect of herbivory on vegetation structure. In Plant Form and Vegetation Structure; Werger, M.J.A., van der Aart, P.J.M., During, H.J., Verhoeven, J.T.A., Eds.; SPB Academic Publishing: Hague, The Netherlands, 1988; pp. 317–330. [Google Scholar]
- Liu, C.; Song, X.; Wang, L.; Wang, D.; Zhou, X.; Liu, J.; Zhao, X.; Li, J.; Lin, H. Effects of grazing on soil nitrogen spatial heterogeneity depend on herbivore assemblage and pre-grazing plant diversity. J. Appl. Ecol. 2015, 53, 242–250. [Google Scholar] [CrossRef]
- Yan, R.; Xin, X.; Yan, Y.; Wang, X.; Zhang, B.; Yang, G.; Liu, S.; Deng, Y.; Li, L. Impacts of Differing Grazing Rates on Canopy Structure and Species Composition in Hulunber Meadow Steppe. Rangel. Ecol. Manag. 2015, 68, 54–64. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.; Douzet, R.; Aubert, S.; Lavorel, S. A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Funct. Ecol. 2010, 24, 1192–1201. [Google Scholar] [CrossRef]
- Roche, P.; Díaz-Burlinson, N.; Gachet, S. Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable? Plant Ecol. 2004, 174, 37–48. [Google Scholar] [CrossRef]
- Reid, R.S.; Galvin, K.A.; Kruska, R.S. Global Significance of Extensive Grazing Lands and Pastoral Societies: An Introduction. In Fragmentation in Semi-Arid and Arid Landscapes; Springer: Dordrecht, The Netherlands, 2008; pp. 1–24. [Google Scholar]
- Ganjurjav, H.; Zhang, Y.; Gornish, E.S.; Hu, G.; Li, Y.; Wan, Y.; Gao, Q. Differential resistance and resilience of functional groups to livestock grazing maintain ecosystem stability in an alpine steppe on the Qinghai-Tibetan Plateau. J. Environ. Manag. 2019, 251, 109579. [Google Scholar] [CrossRef] [PubMed]
- Porensky, L.; Derner, J.D.; Augustine, D.J.; Milchunas, D.G. Plant Community Composition After 75 Yr of Sustained Grazing Intensity Treatments in Shortgrass Steppe. Rangel. Ecol. Manag. 2017, 70, 456–464. [Google Scholar] [CrossRef]
- Semmartin, M.; Aguiar, M.R.; Distel, R.A.; Moretto, A.S.; Ghersa, C.M. Litter quality and nutrient cycling affected by grazing-induced species replacements along a precipitation gradient. Oikos 2004, 107, 148–160. [Google Scholar] [CrossRef]
- Jones, W.M.; Fraser, L.H.; Curtis, P.J. Plant community functional shifts in response to livestock grazing in intermountain depressional wetlands in British Columbia, Canada. Biol. Conserv. 2011, 144, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Xiong, W.; Xu, Y.; Jiang, L.; Zhu, E.; Zhan, W.; He, Y.; Zhu, D.; Zhu, Q.; Peng, C.; et al. Soil properties and species composition under different grazing intensity in an alpine meadow on the eastern Tibetan Plateau, China. Environ. Monit. Assess. 2016, 188, 678. [Google Scholar] [CrossRef]
- Miehe, G.; Miehe, S.; Bach, K.; Nölling, J.; Hanspach, J.; Reudenbach, C.; Kaiser, K.; Wesche, K.; Mosbrugger, V.; Yang, Y.; et al. Plant communities of central Tibetan pastures in the Alpine Steppe/Kobresia pygmaea ecotone. J. Arid. Environ. 2011, 75, 711–723. [Google Scholar] [CrossRef]
- Adler, P.; Milchunas, D.G.; Sala, O.E.; Burke, I.C.; Lauenroth, W.K. Plant traits and ecosystem grazing effects: Comparison of U.S. sagebrush steppe and patagonian steppe. Ecol. Appl. 2005, 15, 774–792. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D. The ecological consequences of changes in biodiversity: A search for general principles. Ecology 1999, 80, 1455–1474. [Google Scholar]
- Dumont, B.; Rossignol, N.; Loucougaray, G.; Carrère, P.; Chadoeuf, J.; Fleurance, G.; Bonis, A.; Farruggia, A.; Gaucherand, S.; Ginane, C.; et al. When does grazing generate stable vegetation patterns in temperate pastures? Agric. Ecosyst. Environ. 2012, 153, 50–56. [Google Scholar] [CrossRef]
- Pillar, V.D.; Blanco, C.C.; Müller, S.C.; Sosinski, E.E.; Joner, F.; Duarte, L.D.S. Functional redundancy and stability in plant communities. J. Veg. Sci. 2013, 24, 963–974. [Google Scholar] [CrossRef]
- Bermejo, L.; De Nascimento, L.; Mata, J.; Fernandez-Lugo, S.; Camacho, A.; Arévalo, J.R. Responses of plant functional groups in grazed and abandoned areas of a Natural Protected Area. Basic Appl. Ecol. 2012, 13, 312–318. [Google Scholar] [CrossRef]
- Naeem, S. Species Redundancy and Ecosystem Reliability. Conserv. Boil. 1998, 12, 39–45. [Google Scholar] [CrossRef]
- Wellnitz, T.; Poff, N.L. Functional redundancy in heterogeneous environments: Implications for conservation. Ecol. Lett. 2001, 4, 177–179. [Google Scholar] [CrossRef]
- Yuan, W.; Zhou, G. Responses of three Stipa communities net primary productivity along Northeast China Transect to seasonal distribution of precipitation. J. Appl. Ecol. 2005, 16, 605–609. [Google Scholar]
- Yan, Y.; Yan, R.; Chen, J.; Xin, X.; Eldridge, D.J.; Shao, C.; Wang, X.; Lv, S.; Jin, D.; Chen, J.; et al. Grazing modulates soil temperature and moisture in a Eurasian steppe. Agric. For. Meteorol. 2018, 262, 157–165. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
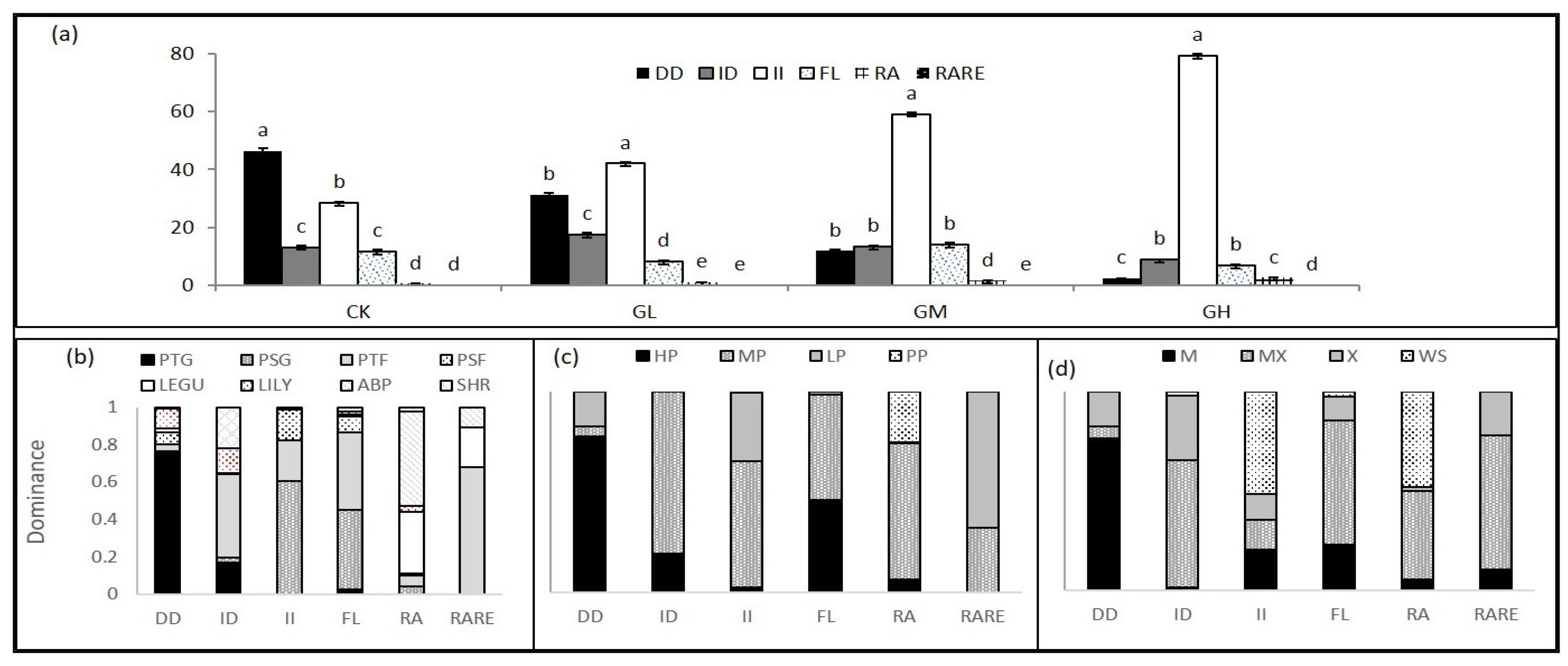{kind=link}
| Change Pattern | Number of Species | Species (Ranked by Dominance) |
|---|---|---|
| Decreaser | 13 | L. chinensis, I. ventricosa, H. altaicus, Bromus inermis, Galium verum, Cymbaria dahurica, Achnatherum sibiricum, Astragalus melilotoides, Vicia amoena, Veronica incana, Clematis hexapetla, T. petaloideum, Lilium tenuifolium. |
| Compensator | 11 | T. squarrosum, Artemisia frigida, Stipa capelata, Allium bidentatum, Bupleurum, Vicia angustifolia, A. tenuissimum, A. ramosum, Poa pratensis, Dianthus chinensis, Helictotrichon schellianum. |
| Increaser | 14 | C. duriuscula, A. laciniata, Pulsatilla turczaninovii, C. squarrosa, K. cristata, Potentilla acaulis, Schizonepeta multifida, A. adsurgens, Taraxacum mongolicum, P. verticillaris, P. tanacetifolia, Oxytropis myriophylla, Sibbaldia adpressa, Scorzonera divaricata. |
| Neutral | 11 | C. pediformis, Serratula centaur oides, A. dracunculus, P. bifurca, Adenophora stenophylla, A. cristatum, Ixeris sonchifolia, I. tigridia, Melissilus ruthenicus, Tephroseris kirlowii, Viola dissecta |
| Randomer | 25 | Plantago depressa, T. lanceolata, Hierochloe glabra, G. multiflora, Festuca ovina, A. commutate, A. gmelinii, veratrum nigrum, A. condensatum, L. apetalum, Orobanche coerulescens, G. squarrosa, Gentianopsis barbata, Stilpnolepis centiflora, C. glaucum, Convolvulus ammannii, Lychnis sibirica, L. leontopodioides, Euphorbia esula, S. divaricata, Sanguisorba officinalis, Thlaspi cochleariform, Thesium longifolium, Inula japonica, Saussurea scabrida. (IV > 0.05) |
| Rare species | 4 | Dracocephalum ruyschiana, A. galactites, Dontostemon micranthus, Silene jenisseensis |
| Treatment | Species Pattern | |||||
|---|---|---|---|---|---|---|
| Grazing Intensity | Decrease | Compensator | Increaser | Neutral | Random | RARE |
| Control | 46.29a | 13.16b | 28.43d | 11.62b | 0.49c | 0.01d |
| Light | 31.30b | 17.52a | 42.41c | 8.13c | 0.88c | 0.09a |
| Moderate | 11.83c | 13.34b | 59.18b | 14.01a | 1.42b | 0.07b |
| Heavy | 2.43c | 8.8c | 79.24a | 6.92d | 2.56a | 0.05c |
| Plant Functional groups | ||||||
| PTG | 17.55a | 2.22c | 0.00d | 0.27bc | 0.00c | 0.00a |
| PSG | 0.00e | 0.36e | 31.54a | 4.30a | 0.06c | 0.00a |
| PTF | 0.8d | 5.85a | 11.40b | 4.22a | 0.08c | 0.04a |
| PSF | 1.54c | 0.11f | 8.51c | 0.87b | 0.01c | 0.00a |
| LEGU | 0.54d | 0.00f | 0.77d | 0.00c | 0.46b | 0.04a |
| LILY | 2.44b | 1.74d | 0.00d | 0.14bc | 0.04c | 0.00a |
| ABP | 0.10e | 0.00f | 0.00d | 0.25bc | 0.69a | 0.01a |
| SHR | 0.00e | 2.91b | 0.00d | 0.13bc | 0.037c | 0.00a |
| Water ecotype | ||||||
| Mesophyte | 17.59a | 0.20c | 10.74b | 2.69b | 0.07b | 0.01b |
| Mesoxerophyte | 1.34c | 8.45a | 7.74c | 6.33a | 0.61a | 0.04a |
| Xerophyte | 4.01b | 4.28b | 6.94c | 1.24c | 0.04b | 0.01b |
| Widespread | 0.00d | 0.27c | 26.81a | 1.24c | 0.66a | 0.00b |
| Palatability | ||||||
| High palatability | 17.83a | 2.59b | 1.27c | 4.70a | 0.07c | 0.00c |
| Medium palatability | 1.22c | 10.61a | 32.90a | 5.34a | 0.92a | 0.02b |
| Low palatability | 3.93b | 0.00c | 17.73b | 0.14b | 0.00c | 0.04a |
| Poisonous plants | 0.00c | 0.00c | 0.29c | 0.00b | 0.35b | 0.00c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zainelabdeen, Y.M.; Yan, R.; Xin, X.; Yan, Y.; Ahmed, A.I.; Hou, L.; Zhang, Y. The Impact of Grazing on the Grass Composition in Temperate Grassland. Agronomy 2020, 10, 1230. https://doi.org/10.3390/agronomy10091230
Zainelabdeen YM, Yan R, Xin X, Yan Y, Ahmed AI, Hou L, Zhang Y. The Impact of Grazing on the Grass Composition in Temperate Grassland. Agronomy. 2020; 10(9):1230. https://doi.org/10.3390/agronomy10091230
Chicago/Turabian StyleZainelabdeen, Yousif Mohamed, Ruirui Yan, Xiaoping Xin, Yuchun Yan, Ahmed Ibrahim Ahmed, Lulu Hou, and Yu Zhang. 2020. "The Impact of Grazing on the Grass Composition in Temperate Grassland" Agronomy 10, no. 9: 1230. https://doi.org/10.3390/agronomy10091230
APA StyleZainelabdeen, Y. M., Yan, R., Xin, X., Yan, Y., Ahmed, A. I., Hou, L., & Zhang, Y. (2020). The Impact of Grazing on the Grass Composition in Temperate Grassland. Agronomy, 10(9), 1230. https://doi.org/10.3390/agronomy10091230