Adult Desert Locust Swarms, Schistocerca gregaria, Preferentially Roost in the Tallest Plants at Any Given Site in the Sahara Desert


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Species
2.3. Sampling Regime
2.4. Night-Roosting Plants
2.5. Locust Group Size
2.6. Defensive Response
2.7. Statistical Analysis
3. Results
3.1. Plant Characteristics
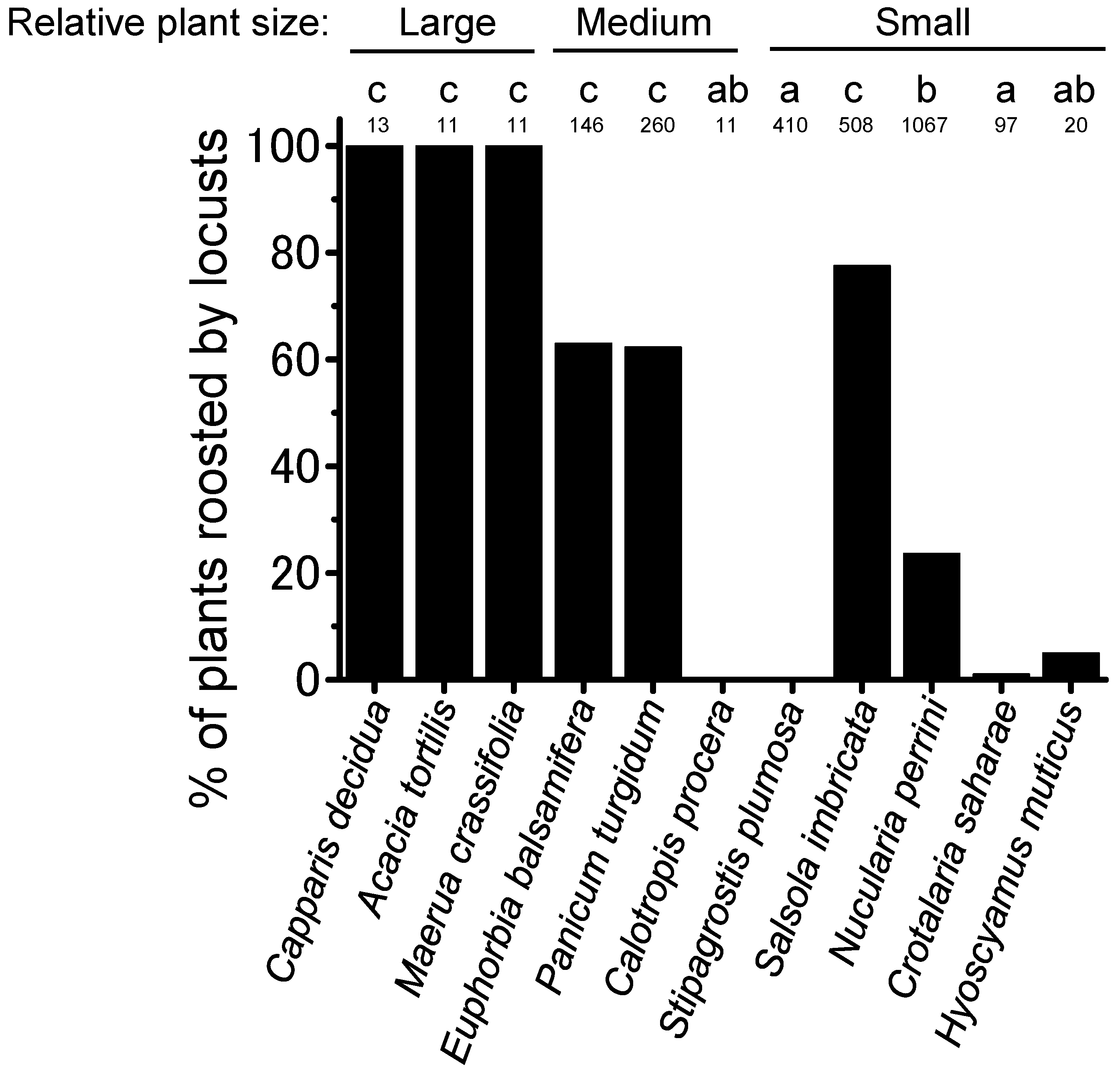
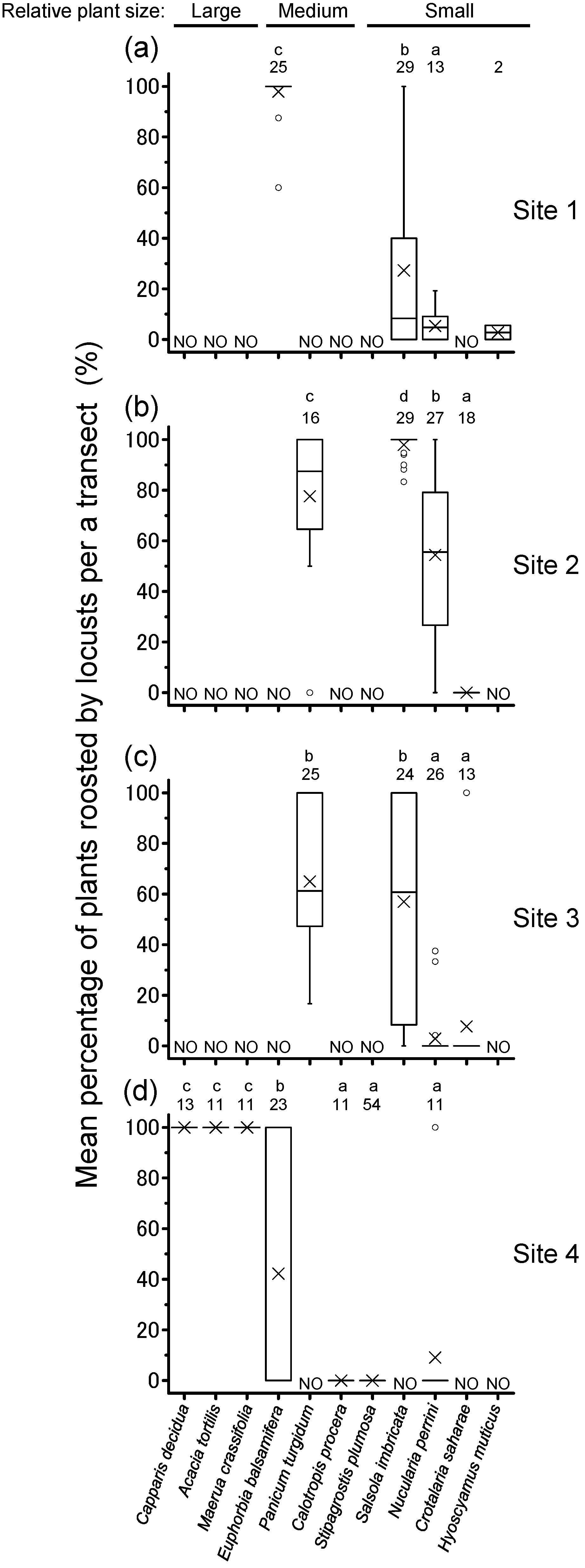
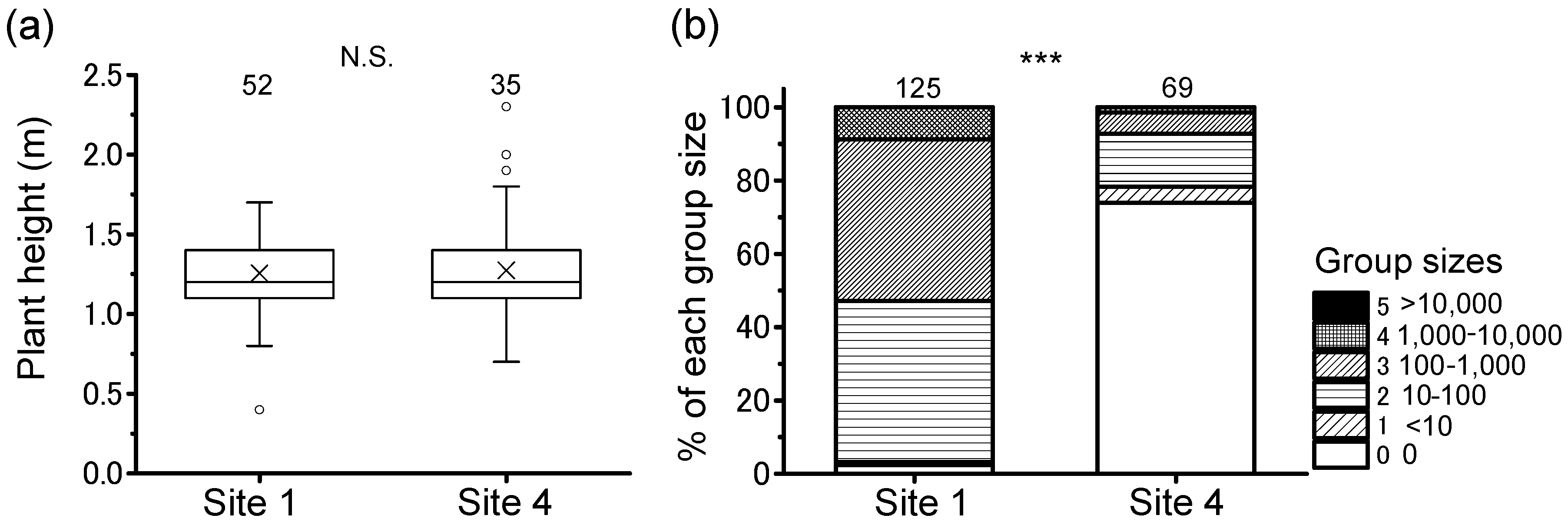
3.2. Daily Cyclical Movement and Night-Roosting Plant Preference
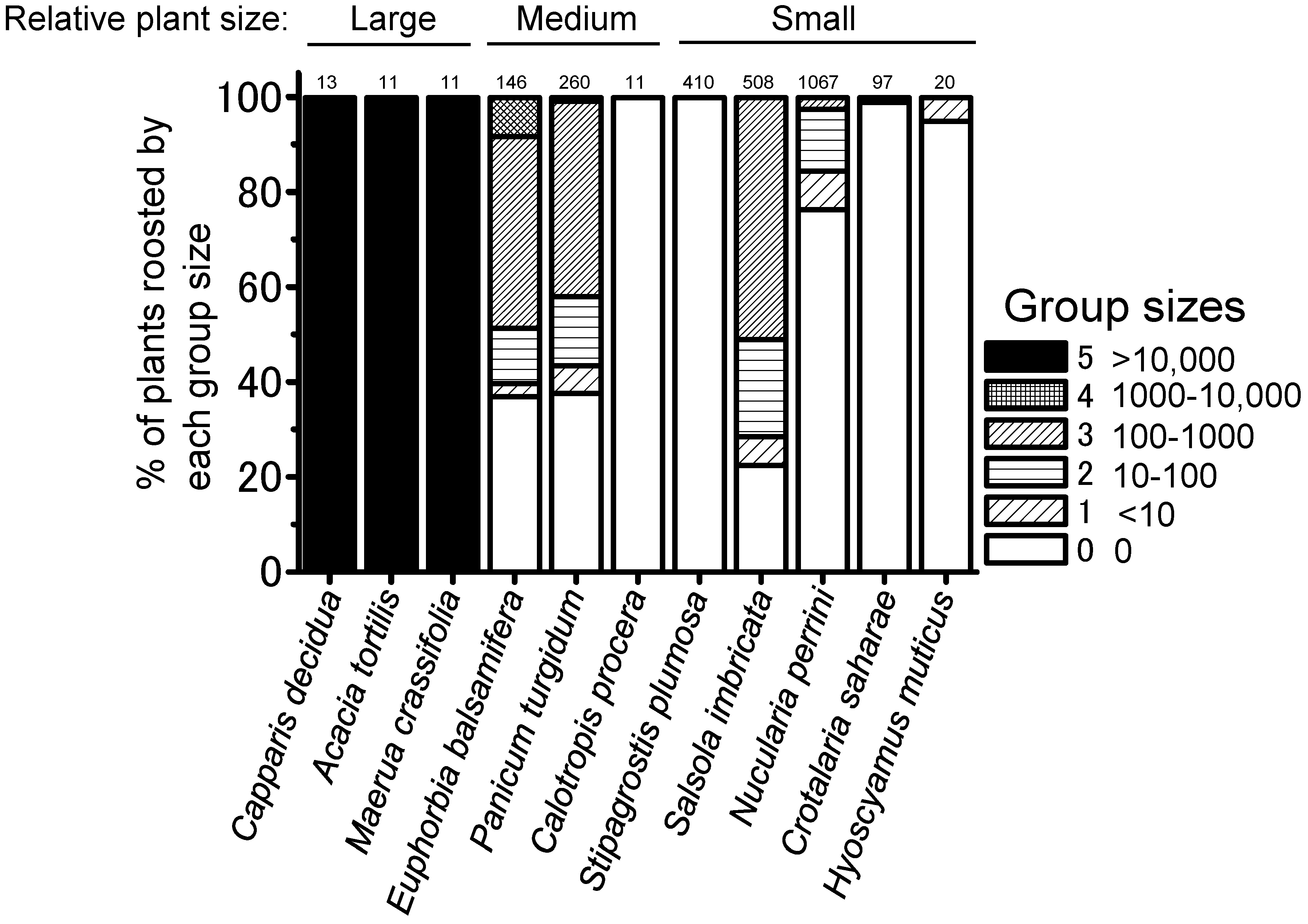
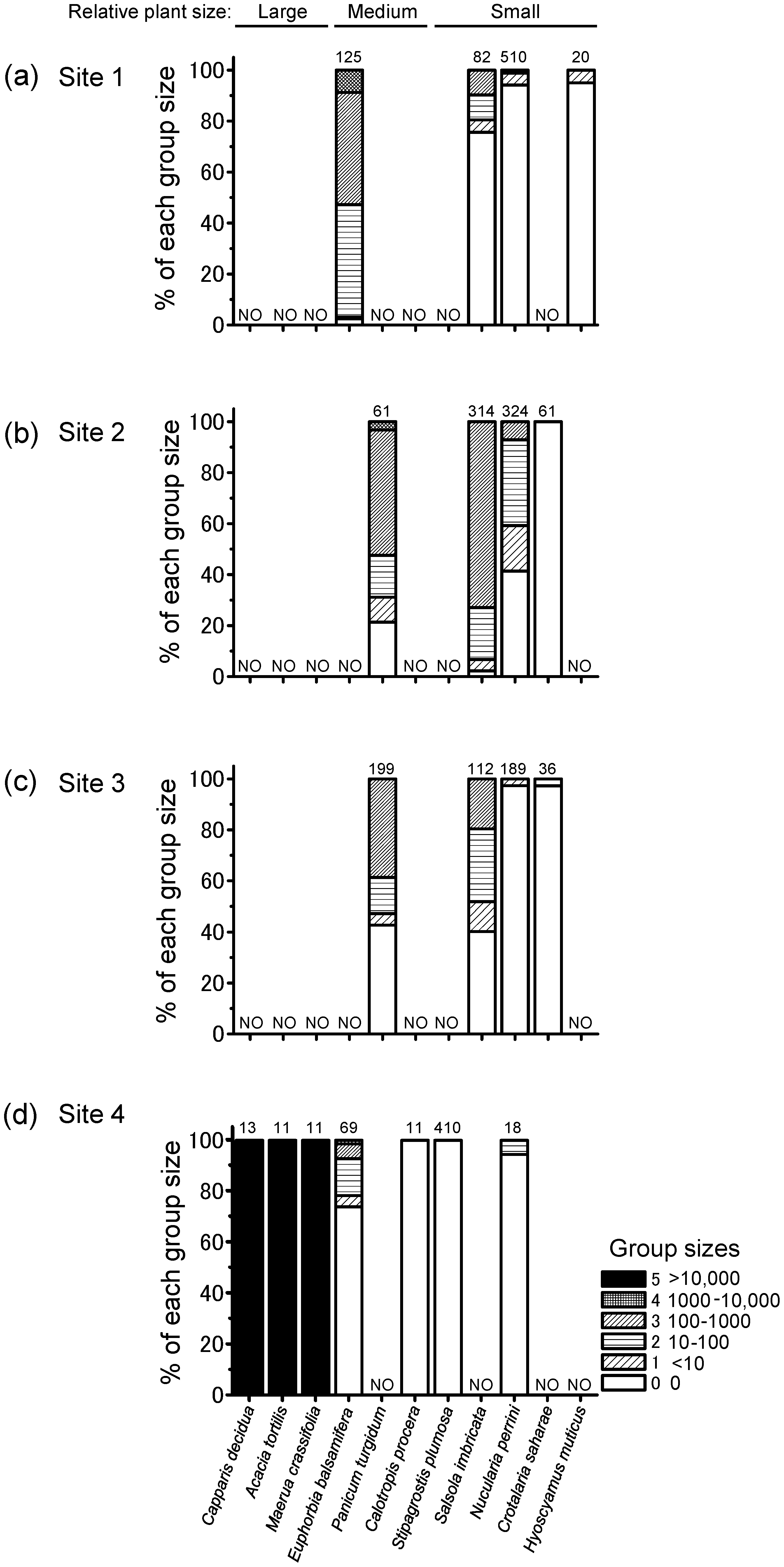
3.3. Group Size on Roosting Plants
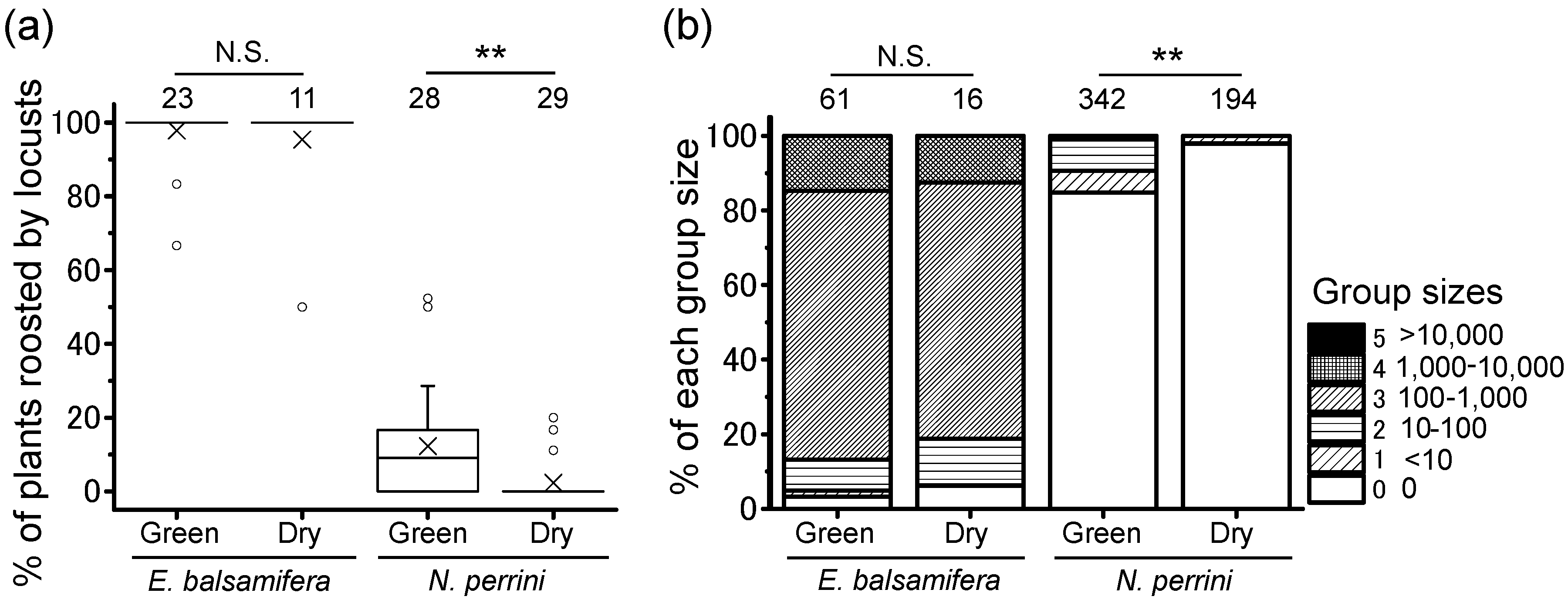
3.4. Plant Conditions: Green vs. Dry
4. Discussion
4.1. Night-Roosting Plant Preference
4.2. Local Locust Group Size on Night-Roosting Plants
4.3. Applications and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sword, G.A.; Lecoq, M.; Simpson, S.J. Phase polyphenism and preventative locust management. J. Insect Physiol. 2010, 56, 949–957. [Google Scholar] [CrossRef]
- Le Gall, M.; Overson, R.; Cease, A. A global review on locusts (Orthoptera: Acrididae) and their interactions with livestock grazing practices. Front. Ecol. Evol. 2019, 7, 263. [Google Scholar] [CrossRef] [Green Version]
- Brader, L.; DJibo, H.; Faya, F.G.; Ghaout, S.; Lazar, M.; Luzietoso, P.N.; Ould Babah, M.A. Towards a More Effective Response to Desert Locusts and Their Impacts on Food Security, Livelihood and Poverty; Multilateral Evaluation of the 2003-05 Desert Locust Campaign; FAO: Rome, Italy, 2006; p. 114. [Google Scholar]
- Cressman, K. Desert Locust. In Biological and Environmental Hazards, Risks, and Disasters; Shroder, J.F., Sivanpillai, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; ISBN 9780123948472. [Google Scholar]
- Van Huis, A.; Cressman, K.; Magor, J.I. Preventing desert locust plagues: Optimizing management interventions. Entomol. Exp. Appl. 2017, 122, 191–214. [Google Scholar] [CrossRef]
- Waloff, Z. The upsurges and recessions of the desert locust plague: An historical survey. Anti Locust Mem. 1966, 8, 1–111. [Google Scholar]
- Steedman, A. The Locust Handbook; Natural Resources Institute: Chatham, UK, 1990. [Google Scholar]
- Showler, A.T. Desert locust control: The effectiveness of proactive interventions and the goal of outbreak prevention. Am. Entomol. 2019, 65, 180–191. [Google Scholar] [CrossRef]
- Lecoq, M. Desert locust management: From ecology to anthropology. J. Orthoptera Res. 2005, 14, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and grasshopper management. Annu. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef]
- Cullen, D.A.; Cease, A.J.; Latchininsky, A.V.; Ayali, A.; Berry, K.; Buhl, J.; De Keyser, R.; Foquet, B.; Hadrich, J.C.; Matheson, T.; et al. From molecules to management: Mechanisms and consequences of locust phase polyphenism. Adv. Insect Phys. 2017, 53, 167–285. [Google Scholar] [CrossRef]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism:an update. Adv. Insect Phys. 2009, 36, 1–286. [Google Scholar] [CrossRef]
- Ayali, A. The puzzle of locust density-dependent phase polyphenism. Curr. Opin. Insect Sci. 2019, 35, 41–47. [Google Scholar] [CrossRef]
- Rogers, S.M.; Cullen, D.A.; Anstey, M.L.; Burrows, M.; Despland, E.; Dodgson, T.; Matheson, T.; Ott, S.R.; Stettin, K.; Sword, G.A.; et al. Rapid behavioural gregarization in the desert locust, Schistocerca gregaria entails synchronous changes in both activity and attraction to conspecifics. J. Insect Physiol. 2014, 65, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Uvarov, B.P. Grasshoppers and Locusts; Centre for Overseas Pest Research: London, UK, 1977; Volume 2. [Google Scholar]
- Magor, J.I.; Lecoq, M.; Hunter, D.M. Preventive control and Desert Locust plagues. Crop Prot. 2008, 27, 1527–1533. [Google Scholar] [CrossRef]
- Krall, S.; Peveling, R.; Diallo, D.B. Economics of Desert Locust Control. In New Strategies in Locust Control; Birkhäuser Verlag: Basel, Switzerland, 1997. [Google Scholar]
- Kimathi, E.; Tonnang, H.E.Z.; Subramanian, S.; Cressman, K.; Abdel-Rahman, E.M.; Tesfayohannes, M.; Niassy, S.; Torto, B.; Dubois, T.; Tanga, C.M.; et al. Prediction of breeding regions for the desert locust Schistocerca gregaria in East Africa. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Gay, P.E.; Lecoq, M.; Piou, C. The limitations of locust preventive management faced with spatial uncertainty: Exploration with a multi-agent model. Pest Manag. Sci. 2020, 76, 1094–1102. [Google Scholar] [CrossRef]
- Dobson, H.M. Desert Locust Guidelines 4: Control; FAO: Rome, Italy, 2001. [Google Scholar]
- Rainey, R.C.; Sayer, H.J. Some recent developments in the use of aircraft against flying locust swarms. Nature 1953, 172, 224–228. [Google Scholar] [CrossRef]
- Gunn, D.L.; Perry, F.C.; Seymour, W.G.; Telford, T.M.; Wright, E.N.; Yeo, D. Behaviour of the Desert Locust (Schistocerca gregaria Forskål) in Kenya in relation to aircraft spraying. Anti Locust Bull. 1948, 3, 1–70. [Google Scholar]
- Joyce, R.J.V. The evolution of an aerial application system for the control of Desert Locusts. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1979, 287, 305–314. [Google Scholar] [CrossRef]
- Symmons, P.M.; Cressman, K. Desert Locust Guidelines 1: Biology and Behavior; FAO: Rome, Italy, 2001. [Google Scholar]
- Maeno, K.O.; Ould Ely, S.; Nakamura, S.; Abdellaoui, K.; Cissé, S.; Jaavar, M.E.; Ould Mohamed, S.; Atheimine, M.; Ould Babah, M.A. Daily microhabitat shifting of solitarious-phase Desert locust adults: Implications for meaningful population monitoring. Springerplus 2016, 5, 107. [Google Scholar] [CrossRef] [Green Version]
- Maeno, K.O.; Ould Ely, S.; Ould Mohamed, S.; Jaavar, M.E.H.; Nakamura, S.; Ould Babah Ebbe, M.A. Defence tactics cycle with diel microhabitat choice and body temperature in the desert locust, Schistocerca gregaria. Ethology 2019, 125, 250–261. [Google Scholar] [CrossRef]
- Babah Ebbe, M.A. Biogéographie du Criquet pèlerin en Mauritanie; Hermann: Paris, France, 2010. [Google Scholar]
- Duranton, J.-F.; Foucart, A.; Gay, P.-E. Florule des Biotopes du Criquet Pèlerin en Afrique de l’Ouest et u Nord-Ouest à L’usage des Prospecteurs de la Lutte Antiacridienne; FAO-CLCPRO: Montpellier, France, 2012. [Google Scholar]
- Cisse, S.; Ghaout, S.; Mazih, A.; Ould Babah Ebbe, M.A.; Benahi, A.S.; Piou, C. Effect of vegetation on density thresholds of adult desert locust gregarization from survey data in Mauritania. Entomol. Exp. Appl. 2013, 149, 159–165. [Google Scholar] [CrossRef]
- Ould Babah, M.A.; Sword, G.A. Linking locust gregarization to local resource distribution patterns across a large spatial scale. Environ. Entomol. 2004, 33, 1577–1583. [Google Scholar] [CrossRef]
- Maeno, K.O.; Piou, C.; Ould Ely, S.; Ould Mohamed, S.; Jaavar, M.E.H.; Ould Babah, M.A.; Nakamura, S. Field observations of the sheltering behavior of the solitarious phase of the Desert locust, Schistocerca gregaria, with particular reference to antipredator strategies. Jpn. Agric. Res. Q. 2012, 46, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Rainey, R.C. Meteorology and the migration of desert locusts. Anti Locust Mem. 1963, 7, 1–115. [Google Scholar]
- Maeno, K.O.; Ould Babah Ebbe, M.A. Aggregation site choice by gregarious nymphs of the desert locust, Schistocerca gregaria, in the Sahara Desert of Mauritania. Insects 2018, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Piou, C.; Jaavar Bacar, M.E.H.; Ould Babah Ebbe, M.A.; Chihrane, J.; Ghaout, S.; Cisse, S.; Lecoq, M.; Ben Halima, T. Mapping the spatiotemporal distributions of the Desert Locust in Mauritania and Morocco to improve preventive management. Basic Appl. Ecol. 2017, 25, 37–47. [Google Scholar] [CrossRef]
- Piou, C.; Gay, P.E.; Benahi, A.S.; Babah Ebbe, M.A.O.; Chihrane, J.; Ghaout, S.; Cisse, S.; Diakite, F.; Lazar, M.; Cressman, K.; et al. Soil moisture from remote sensing to forecast desert locust presence. J. Appl. Ecol. 2019, 56, 966–975. [Google Scholar] [CrossRef]
- Uvarov, B.P. Grasshoppers and Locusts; Cambridge University Press: Cambridge, UK, 1966; Volume 1. [Google Scholar]
- Norris, M.J. Sexual maturation in the desert locust with special reference to the effects of grouping. Anti Locust Bull. 1954, 18, 1–43. [Google Scholar]
- Maeno, K.O.; Piou, C.; Ould Ely, S.; Ould Babah, M.A.; Pélissié, B.; Ould Mohamed, S.; Jaavar, M.E.H.; Etheimine, M.; Nakamura, S. Plant size-dependent escaping behavior of gregarious nymphs of the Desert locust, Schistocerca gregaria. J. Insect Behav. 2013, 26, 623–633. [Google Scholar] [CrossRef]
- Maeno, K.O.; Ould Ely, S.; Ould Mohamed, S.; Jaavar, M.E.H.; Nakamura, S.; Ould Babah Ebbe, M.A. Behavioral plasticity in anti-predator defense in the desert locust. J. Arid Environ. 2018, 158, 47–50. [Google Scholar] [CrossRef]
- R Development Core T. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Alcock, J.; Dodson, G. The diverse mating systems of hilltopping insects. Am. Entomol. 2008, 54, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.E., Jr.; Blumstein, D.T. Escaping from Predators: An Integrative View of Escape Decisions; Cambridge University Press: London, UK, 2015. [Google Scholar]
- Whitman, D.W. Function and evolution of thermoregulation in the desert grasshopper Taeniopoda eques. J. Anim. Ecol. 1988, 57, 369–383. [Google Scholar] [CrossRef]
- Chappell, M.A.; Whitman, D.W. Grasshopper Thermoregulation. In Biololgy Grasshoppers; Chapman, R.F., Joern, A., Eds.; John Wiley: New York, NY, USA, 1990; pp. 143–172. [Google Scholar]
- Hamilton, W.D. Geometry for the selfish herd. J. Theor. Biol. 1971, 31, 295–311. [Google Scholar] [CrossRef]
- Stauffer, T.W.; Hegrenes, S.G.; Whitman, D.W. A laboratory study of oviposition site preferences in the lubber grasshopper, Romalea guttata (Houttuyn). J. Orthoptera Res. 1998, 1998, 217–221. [Google Scholar] [CrossRef]
- Sword, G.A.; Lorch, P.D.; Gwynne, D.T. Migratory bands give crickets protection. Nature 2005, 433, 703. [Google Scholar] [CrossRef] [PubMed]
- Fonteyn, P.J.; Mahall, B.E. An experimental analysis of structure in a desert plant community. J. Ecol. 1981, 69, 883–896. [Google Scholar] [CrossRef]
- Kennedy, J.S. The behaviour of the desert locust Schistocerca gregaria (Forsk.)(Orthoptera) in an outbreak centre. Trans. R. Entomol. Soc. Lond. 1939, 89, 385–542. [Google Scholar] [CrossRef]
- Ellis, P.E.; Ashall, C. Field studies on diurnal behaviour, movement and aggregation in the desert locust (Schistocerca gregaria Forskål). Anti Locust Bull. 1957, 25, 1–103. [Google Scholar]
- Collett, M.; Despland, E.; Simpson, S.J.; Krakauer, D.C. Spatial scales of desert locust gregarization. Proc. Natl. Acad. Sci. USA 1998, 95, 13052–13055. [Google Scholar] [CrossRef] [Green Version]
- Despland, E.; Collett, M.; Simpson, S.J. Small-scale processes in desert locust swarm formation: How vegetation patterns influence gregarization. Oikos 2000, 88, 652–662. [Google Scholar] [CrossRef]
- Van der Werf, W.; Woldewahid, G.; van Huis, A.; Butrous, M.; Sykora, K. Plant communities can predict the distribution of solitarious desert locust, Schistocerca gregaria. J. Appl. Ecol. 2005, 42, 989–997. [Google Scholar] [CrossRef]
- Simpson, S.J.; Despland, E.; Hägele, B.F.; Dodgson, T. Gregarious behavior in desert locusts is evoked by touching their back legs. Proc. Natl. Acad. Sci. USA 2001, 98, 3895–3897. [Google Scholar] [CrossRef] [Green Version]
- Cullen, D.A.; Sword, G.A.; Dodgson, T.; Simpson, S.J. Behavioural phase change in the Australian plague locust, Chortoicetes terminifera, is triggered by tactile stimulation of the antennae. J. Insect Physiol. 2010, 56, 937–942. [Google Scholar] [CrossRef]
- Maeno, K.; Tanaka, S.; Harano, K.-I. Tactile stimuli perceived by the antennae cause the isolated females to produce gregarious offspring in the desert locust, Schistocerca gregaria. J. Insect Physiol. 2011, 57, 74–82. [Google Scholar] [CrossRef]
- Maeno, K.O.; Piou, C.; Ghaout, S. The desert locust, Schistocerca gregaria, plastically manipulates egg size by regulating both egg numbers and production rate according to population density. J. Insect Physiol. 2020, 122, 104020. [Google Scholar] [CrossRef]
- Beauchamp, G. Social Predation: How Group Living Benefits Predators and Prey; Academic Press: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-407228-2. [Google Scholar]
- Roffey, J.; Popov, G. Environmental and behavioural processes in a desert locust outbreak. Nature 1968, 219, 446–450. [Google Scholar] [CrossRef]
- Shashar, N.; Sabbah, S.; Aharoni, N. Migrating locusts can detect polarized reflections to avoid flying over the sea. Biol. Lett. 2005, 1, 472–475. [Google Scholar] [CrossRef] [Green Version]
- Piou, C.; Lebourgeois, V.; Benahi, A.S.; Bonnal, V.; Jaavar, M.E.H.; Lecoq, M.; Vassal, J.-M. Coupling historical prospection data and a remotely-sensed vegetation index for the preventative control of Desert locusts. Basic Appl. Ecol. 2013, 14, 593–604. [Google Scholar] [CrossRef]
- Latchininsky, A.; Piou, C.; Franc, A.; Soti, V. Applications of Remote Sensing to Locust Management. In Land Surface Remote Sensing, Environment and Risks; Baghdadi, N., Zribi, M., Eds.; ISTE Press: London, UK; Elsevier: Oxford, UK, 2016; pp. 263–293. ISBN 9780081012659. [Google Scholar]
- Cressman, K. Role of remote sensing in desert locust early warning. J. Appl. Remote Sens. 2013, 7, 075098. [Google Scholar] [CrossRef]
- Gay, P.E.; Lecoq, M.; Piou, C. Improving preventive locust management: Insights from a multi-agent model. Pest Manag. Sci. 2018, 74, 46–58. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Site 1 | Site 2 | Site 3 | Site 4 | |||
|---|---|---|---|---|---|---|---|
| Data | 2013/12/22 | 2013/12/23 | 2013/12/24 | 2013/12/29 | |||
| Location | 19°55′ N, 16°14′ W | 19°53′ N, 16°18′ W | 19°53′ N, 16°18′ W | 19°24′ N, 16°13′ W | |||
| Plant Species | Family | Types of Plant | Plant Form | ||||
| Capparis decidua | Capparaceae | Perennial | Tree | - | - | - | 0.2 ± 0.3 a |
| Acacia tortilis | Fabaceae | Perennial | Tree | - | - | - | 0.2 ± 0.3 a |
| Maerua crassifolia | Capparaceae | Perennial | Tree | - | - | - | 0.2 ± 0.3 a |
| Euphorbia balsamifera | Euphorbiaceae | Perennial | Tree | 4.2 ± 0.9 b | - | - | 1.2 ± 0.3 a |
| Calotropis procera | Poaceae | Perennial | Bush & tree | - | - | - | 0.2 ± 0.3 a |
| Panicum turgidum | Apocynaceae | Perennial | Bush | - | 2.0 ± 0.9 a | 6.6 ± 0.8 b | - |
| Stipagrostis plumosa | Poaceae | Annual | Bush | - | - | - | 6.8 ± 0.3 b |
| Salsola imbricata | Amaranthaceae | Annual | Bush | 2.7 ± 0.9 ab | 10.5 ± 0.9 b | 3.7 ± 0.8 ab | - |
| Nucularia perrini | Amaranthaceae | Perennial | Bush | 17.0 ± 0.9 c | 10.8 ± 0.9 b | 6.3 ± 0.8 b | 0.3 ± 0.3 a |
| Crotalaria saharae | Fabaceae | Perennial | Bush | - | 2.0 ± 0.9 a | 1.2 ± 0.8 a | - |
| Hyoscyamus muticus | Solanaceae | Annual | Bush | 0.7 ± 0.9 a | - | - | - |
| No. of transects | 30 | 30 | 30 | 60 |
| Plant Species | Relative Size | Maximum Width (m) | Maximum Height (m) | Volume (m3) | n Plants Measured |
|---|---|---|---|---|---|
| Capparis decidua | Large | 9.34 ± 0.23 h | 3.71 ± 0.08 h | 331.5 ± 13.2 d | 18 |
| Acacia tortilis | Large | 7.08 ± 0.20 g | 2.84 ± 0.07 g | 117.5 ± 11.2 c | 25 |
| Maerua crassifolia | Large | 5.03 ± 0.25 f | 2.29 ± 0.08 f | 55.7 ± 14.0 b | 16 |
| Euphorbia balsamifera | Medium | 2.20 ± 0.11 e | 1.26 ± 0.04 e | 5.9 ± 6.0 a | 87 |
| Panicum turgidum | Medium | 2.10 ± 0.12 de | 1.18 ± 0.04 e | 4.9 ± 6.7 a | 70 |
| Calotropis procera | Medium | 0.71 ± 0.26 abc | 1.03 ± 0.09 de | 0.6 ± 15.0 ab | 14 |
| Stipagrostis plumosa | Small | 1.53 ± 0.17 cd | 0.74 ± 0.06 dc | 1.5 ± 9.5 ab | 35 |
| Salsola imbricata | Small | 1.23 ± 0.09 bc | 0.54 ± 0.03 bc | 0.8 ± 5.3 a | 110 |
| Nucularia perrini | Small | 1.00 ± 0.08 abc | 0.43 ± 0.03 ab | 0.5 ± 4.5 a | 157 |
| Crotalaria saharae | Small | 0.76 ± 0.12 ab | 0.32 ± 0.04 a | 0.2 ± 6.6 a | 71 |
| Hyoscyamus muticus | Small | 0.41 ± 0.22 a | 0.34± 0.08 ab | 0.1 ± 12.5 ab | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeno, K.O.; Ould Ely, S.; Ould Mohamed, S.; Jaavar, M.E.H.; Ould Babah Ebbe, M.A. Adult Desert Locust Swarms, Schistocerca gregaria, Preferentially Roost in the Tallest Plants at Any Given Site in the Sahara Desert. Agronomy 2020, 10, 1923. https://doi.org/10.3390/agronomy10121923
Maeno KO, Ould Ely S, Ould Mohamed S, Jaavar MEH, Ould Babah Ebbe MA. Adult Desert Locust Swarms, Schistocerca gregaria, Preferentially Roost in the Tallest Plants at Any Given Site in the Sahara Desert. Agronomy. 2020; 10(12):1923. https://doi.org/10.3390/agronomy10121923
Chicago/Turabian StyleMaeno, Koutaro Ould, Sidi Ould Ely, Sid’Ahmed Ould Mohamed, Mohamed El Hacen Jaavar, and Mohamed Abdallahi Ould Babah Ebbe. 2020. "Adult Desert Locust Swarms, Schistocerca gregaria, Preferentially Roost in the Tallest Plants at Any Given Site in the Sahara Desert" Agronomy 10, no. 12: 1923. https://doi.org/10.3390/agronomy10121923
APA StyleMaeno, K. O., Ould Ely, S., Ould Mohamed, S., Jaavar, M. E. H., & Ould Babah Ebbe, M. A. (2020). Adult Desert Locust Swarms, Schistocerca gregaria, Preferentially Roost in the Tallest Plants at Any Given Site in the Sahara Desert. Agronomy, 10(12), 1923. https://doi.org/10.3390/agronomy10121923