Optimizing Sowing Depth of Tef for Irrigated Mediterranean Conditions: From Laboratory to Field Studies



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Test-Tube Experiment
2.3. Pot Experiment
2.4. Field Experiments
2.5. Statistical Analysis
3. Results
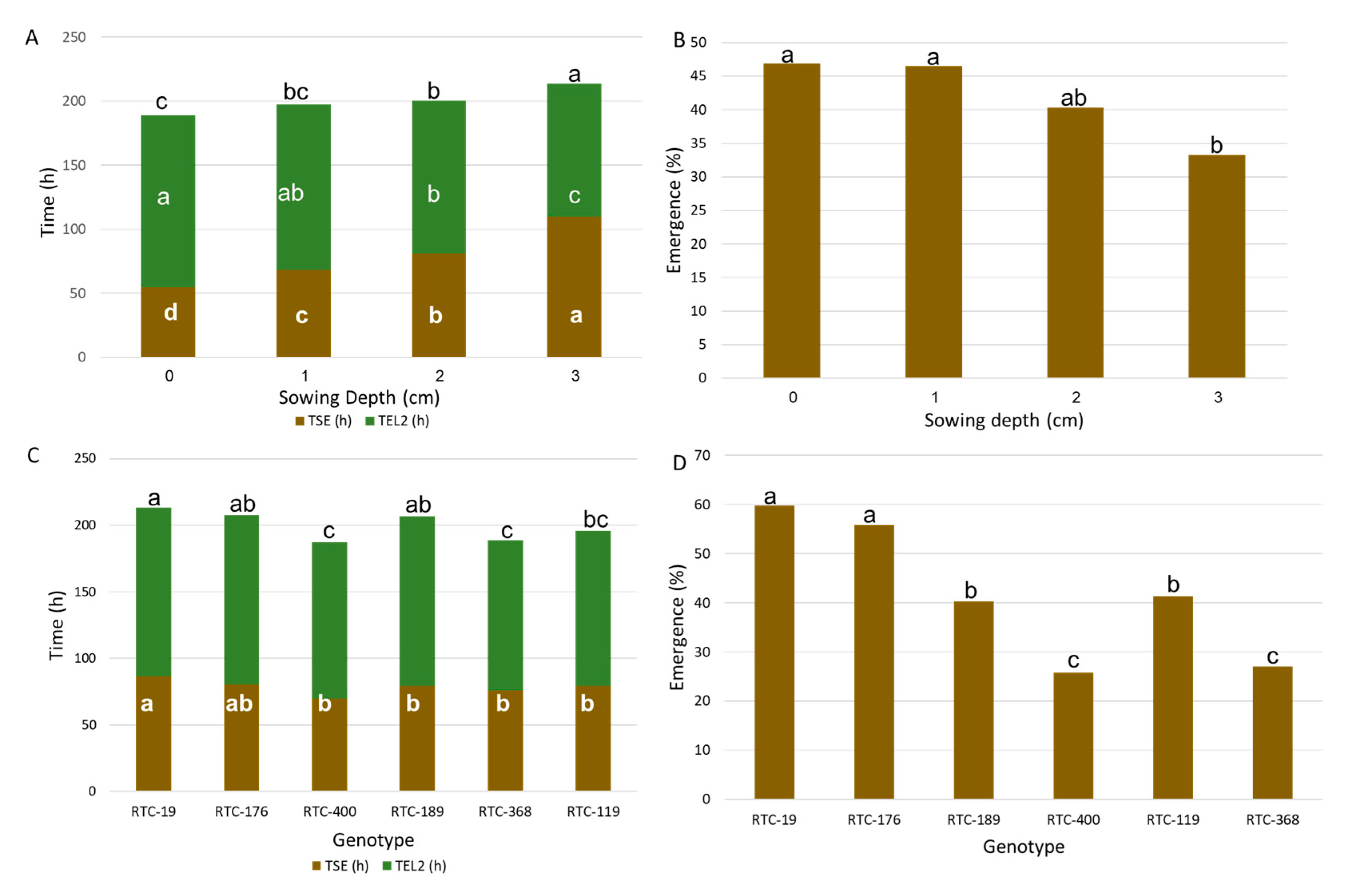
3.1. Test-Tube Experiment: Seedling Emergence and Establishment
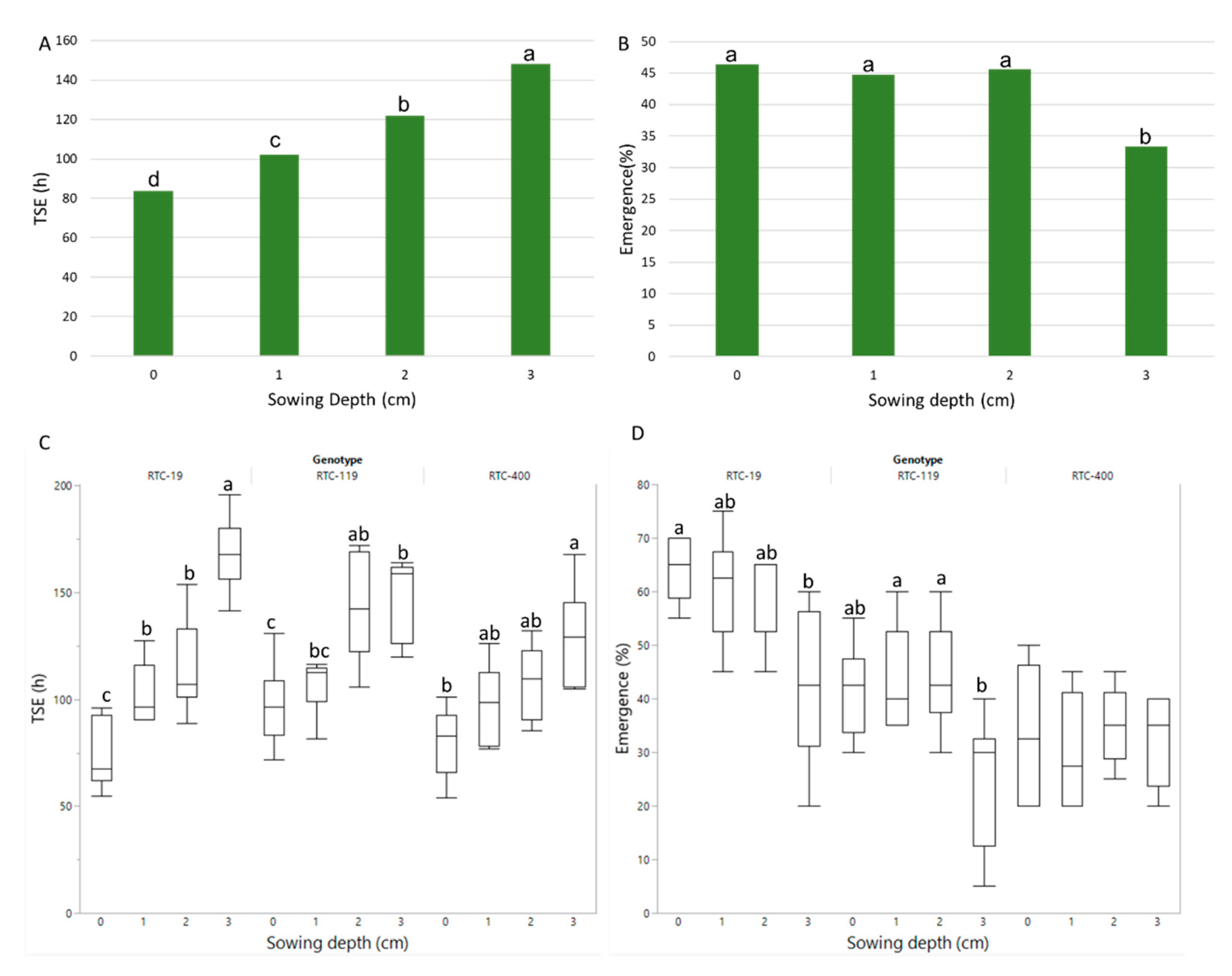
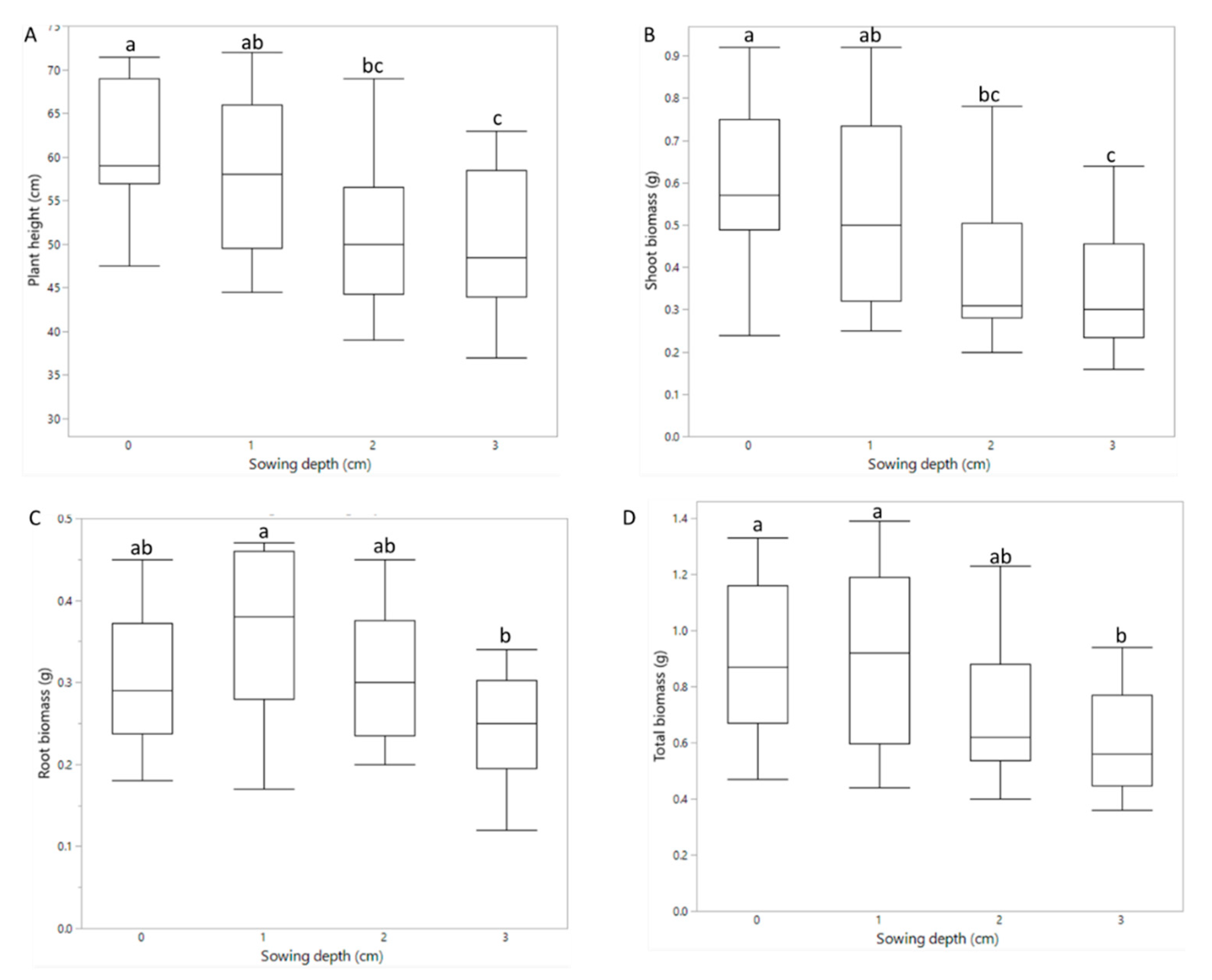
3.2. Pot Experiment: Emergence and Plant Performance
3.3. Pot Experiment: Lodging-Related Traits
3.4. Field Experiments: Emergence and Plant Performance
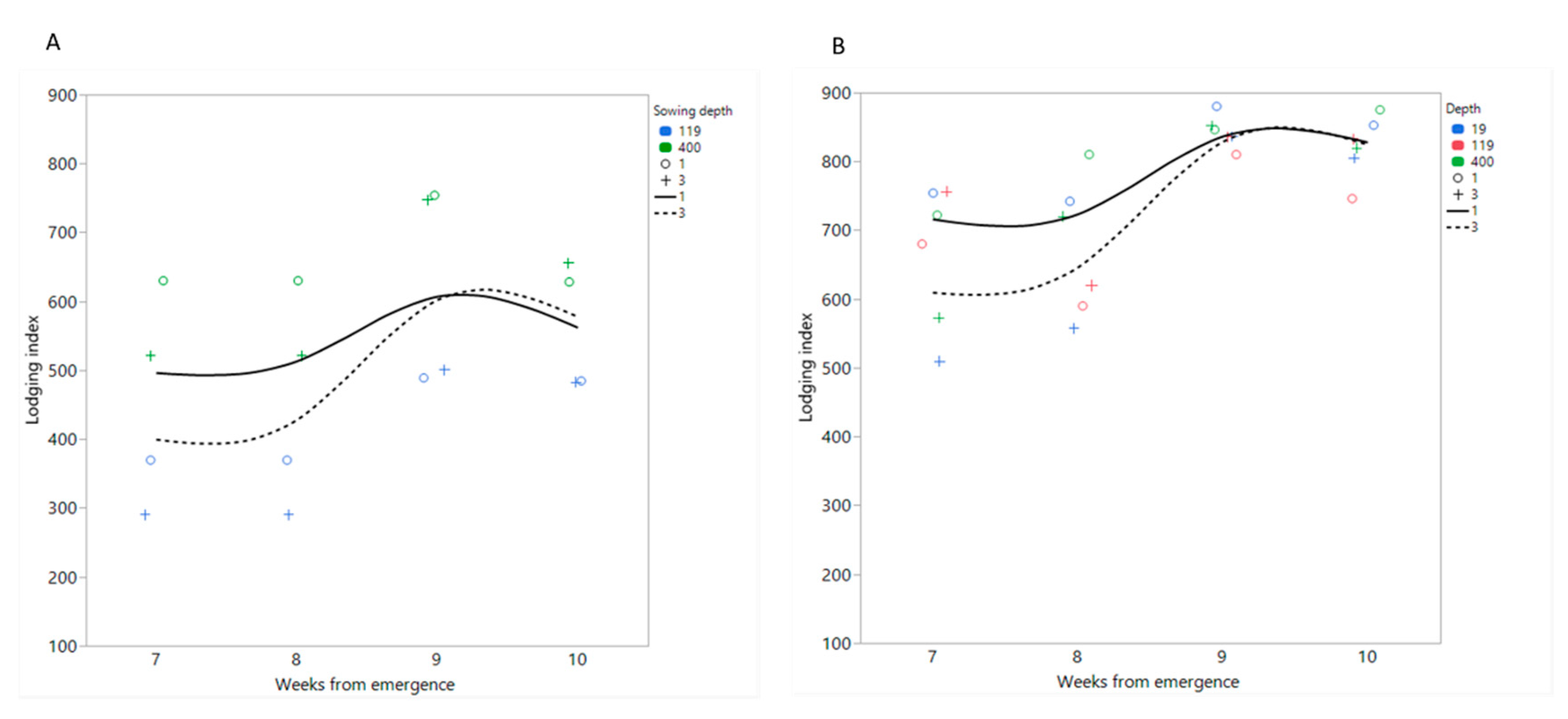
3.5. Field Experiments: Lodging
4. Discussion
4.1. Emergence Time and Rate
4.2. Plant Development and Performance
4.3. Effect of Sowing Depth on Lodging and Related Traits
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sillmann, J.; Kharin, V.V.; Zwiers, F.W.; Zhang, X.; Bronaugh, D. Climate extremes indices in the CMIP5 multimodel ensemble: Part 2. Future climate projections. J. Geophys. Res. Atmos. 2013, 118, 2473–2493. [Google Scholar] [CrossRef]
- Blösch, R.; Plaza-Wüthrich, S.; Barbier de Reuille, P.; Weichert, A.; Routier-Kierzkowska, A.-L.; Cannarozzi, G.; Robinson, S.; Tadele, Z. Panicle angle is an important factor in tef lodging tolerance. Front. Plant Sci. 2020, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Tatham, A.S.; Fido, R.J.; Moore, C.M.; Kasarda, D.D.; Kuzmicky, D.D.; Keen, J.N.; Shewry, P.R. Characterisation of the major prolamins of tef (Eragrostis tef) and finger millet (Eleusine coracana). J. Cereal Sci. 1996, 24, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Miller, D. Teff Grass: Crop Overview and Forage Production Guide. Available online: https://kingsagriseeds.com/wp-content/uploads/2014/12/Teff-Grass-Management-Guide.pdf (accessed on 18 November 2020).
- Norberg, S.; Roseberg, R.; Charlton, B.; Shock, C. Teff: A New Warm-season Annual Grass for Oregon. Oregon State University Extension Catalogue 2009. Available online: https://catalog.extension.oregonstate.edu/sites/catalog/files/project/pdf/em8970.pdf (accessed on 18 November 2020).
- Ben-Zeev, S.; Bimro, J.; Barak, V.; Saranga, Y. Phenotypic diversity and heritability in Eragrostis tef under irrigated Mediterranean conditions. Isr. J. Plant Sci. 2018, 65, 222–231. [Google Scholar] [CrossRef]
- Ketema, S. Tef (Eragrostis tef): Breeding, Genetic Resources, Agronomy, Utilization and Role in Ethiopian Agriculture; Institute of Agricultural Research: Adis Ababa, Ethopia, 1993.
- Stallknecht, G.F.; Gilbertson, K.M.; Eckhoff, J.L. Teff: Food crop for humans and animals. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 231–234. [Google Scholar]
- Gelaw, A.M.; Qureshi, A.S. Tef (Eragrostis tef): A superfood grain from Ethiopia with great potential as an alternative crop for marginal environments. In Emerging Research in Alternative Crops; Hirich, A., Choukr-Allah, R., Ragab, R., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 265–278. [Google Scholar]
- Government of Western Australia. Monitoring Sowing Depth. Available online: https://www.agric.wa.gov.au/mycrop/monitoring-sowing-depth (accessed on 18 November 2020).
- Ketema, S. Eragrostis tef (Zucc.) Trotter: Promoting the Conservation and Use of Underutilized and Neglected Crops; Institute of Plant Genetics and Crop Plant Research: Gatersleben, Germany; International Plant Genetic Resources Institute: Rome, Italy, 1997; ISBN 9290433043. [Google Scholar]
- Mihretie, F.; Tsunekawa, A.; Bitew, Y.; Cheklie, G.; Derebe, B.; Getahun, W.; Beshir, O.; Tadesse, Z.; Asfaw, M. Teff [Eragrostis tef (Zucc.)] rain-fed yield response to planting method, seeding density, and row spacing. Agron. J. 2020. [Google Scholar] [CrossRef]
- Evert, S.; Staggenborg, S.; Olson, B.L.S. Soil temperature and planting depth effects on tef emergence. J. Agron. Crop Sci. 2009, 195, 232–236. [Google Scholar] [CrossRef]
- Debelo, A. Germination, Yield, and Yield Components of Tef (Eragrostis tef (Zucc.) Trotter) as Affected by Environment, Tillage and Weed Control Practices; Oklahoma State University: Stillwater, OK, USA, 1992. [Google Scholar]
- Paff, K.; Asseng, S. A review of tef physiology for developing a tef crop model. Eur. J. Agron. 2018, 94, 54–66. [Google Scholar] [CrossRef]
- Bimro, J. Agronomic Aspects of Teff (Eragrostis tef) Cultivation in Israel; The Hebrew University of Jerusalem: Rehovot, Israel, 2016. [Google Scholar]
- Pinthus, M.J. Lodging in wheat, barley, and oats: The phenomenon, its causes, and preventive measures. Adv. Agron. 1974, 25, 209–263. [Google Scholar]
- Van Delden, S.H.; Vos, J.; Ennos, A.R.; Stomph, T.J. Analysing lodging of the panicle bearing cereal teff (Eragrostis tef). New Phytol. 2010, 186, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Ketema, S. Studies of Lodging Floral Biology and Breeding Techniques in Tef (Eragrostis tef); University of London: London, UK, 1983. [Google Scholar]
- Bayable, M.; Tsunekawa, A.; Haregeweyn, N.; Ishii, T.; Alemayehu, G.; Tsubo, M.; Adgo, E.; Tassew, A.; Tsuji, W.; Asaregew, F.; et al. Biomechanical properties and agro-morphological traits for improved lodging resistance in Ethiopian teff (Eragrostis tef (Zucc.) Trottor) accessions. Agronomy 2020, 10, 1012. [Google Scholar] [CrossRef]
- Ben-Zeev, S.; Rabinovitz, O.; Orlov-Levin, V.; Chen, A.; Graff, N.; Goldwasser, Y.; Saranga, Y. Less Is more: Lower sowing rate of irrigated tef (Eragrostis tef) alters plant morphology and reduces lodging. Agronomy 2020, 10, 570. [Google Scholar] [CrossRef]
- Caldicott, J.J.B.; Nuttall, A.M. A method for the assessment of lodging in cereal crops. J. Natl. Inst. Agric. Bot. 1979, 15, 88–91. [Google Scholar]
- Kirby, E.J.M. Effect of sowing depth on seedling emergence, growth and development in barley and wheat. Field Crops Res. 1993, 35, 101–111. [Google Scholar] [CrossRef]
- Photiades, I.; Hadjichristodoulou, A. Sowing date, sowing depth, seed rate and row spacing of wheat and barley under dryland conditions. Field Crops Res. 1984, 9, 151–162. [Google Scholar] [CrossRef]
- Hanway, J. How a Corn Plant Develops; Special Report 48; Iowa State University: Ames, IA, USA, 1966; Volume 48, pp. 1–18. [Google Scholar]
- Cornish, P.S.; Hindmarsh, S. Seed size influences the coleoptile length of wheat. Aust. J. Exp. Agric. 1988, 28, 521–523. [Google Scholar] [CrossRef]
- Harris, D. The effects of manure, genotype, seed priming, depth and date of sowing on the emergence and early growth Sorghum bicolor (L.) Moench in semi-arid Botswana. Soil Tillage Res. 1996, 40, 73–88. [Google Scholar]
- Berry, P.M.; Sterling, M.; Spink, J.H.; Baker, C.J.; Sylvester-Bradley, R.; Mooney, S.J.; Tams, A.R.; Ennos, A.R. Understanding and reducing lodging in cereals. Adv. Agron. 2004, 84, 217–271. [Google Scholar]
- Josst, M.; Esfeld, K.; Burian, A.; Cannarozzi, G.; Chanyalew, S.; Kuhlemeier, C.; Assefa, K.; Tadele, Z. Semi-dwarfism and lodging tolerance in tef (Eragrostis tef) is linked to a mutation in the alpha-Tubulin 1 gene. J. Exp. Bot. 2015, 66, 933–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Seed Size Group | TSW (mg) | Seed Color | DPH | Test-Tube Exp. | Pot Exp. | Field Exps. |
|---|---|---|---|---|---|---|---|
| RTC-119 | L | 408.4 | Brown | 47 | + | + | + |
| RTC-368 | L | 395.9 | White | 48 | + | ||
| RTC-400 | M | 362.1 | Brown | 45 | + | + | + |
| RTC-189 | M | 341.5 | White | 58 | + | ||
| RTC-176 | S | 272.1 | White | 64 | + | ||
| RTC-19 | S | 252.0 | Brown | 46 | + | + | +(only 2020) * |
| Source of Variation (d.f.) | TSE | Emergence Rate | TEL2 | TSL2 |
|---|---|---|---|---|
| F ratio | ||||
| Sowing depth (SD) (3) | 193.4 *** | 8.2 *** | 16.18 *** | 9.38 *** |
| Genotype (G) (5) | 6.0 *** | 26.4 *** | 3.12 * | 7.42 *** |
| SD*G interaction (15) | 1.1 | 0.9 | 0.58 | 0.48 |
| Block (4) | 0.4 | 0.2 | 8.14 *** | 8.13 *** |
| Source of Variation (d.f.) | TSE | Emergence Rate | Plant Height | Shoot Biomass | Root Biomass | Total Biomass |
|---|---|---|---|---|---|---|
| F ratio | ||||||
| Sowing depth (SD) (3) | 44.52 ** | 6.65 *** | 8.09 *** | 5.64 ** | 5.17 ** | 5.14 ** |
| Genotype (G) (2) | 7.77 ** | 34.66 *** | 2.51 | 0.06 | 0.69 | 0.19 |
| SD*G interaction (6) | 3.14 * | 1.76 | 0.96 | 0.85 | 0.54 | 0.84 |
| Rep. (5) | 3.59 ** | 0.93 | 0.55 | 0.43 | 1.91 | 0.71 |
| Source of Variation (d.f.) | Lodging Angle (°) | Internode number | Tiller # | Crown Diam. (mm) | Root Diam. (mm) | Crown Root # |
|---|---|---|---|---|---|---|
| F ratio | ||||||
| Sowing depth (SD) (3) | 2.33 | 8.18 *** | 1.04 | 2.66 | 2.4 | 0.11 |
| Genotype (G) (2) | 1.06 | 12.15 *** | 0.95 | 4.93 * | 8.29 *** | 0.35 |
| SD*G interaction (6) | 1.31 | 2.22 | 2.22 | 0.61 | 0.53 | 2.2 |
| Rep. (5) | 0.02 | 1.49 | 1.73 | 2.39 | 0.68 | 0.69 |
| Main effect of sowing depth | ||||||
| 0 cm | 62.2 A | 6.61 A | 1.93 A | 2.92 A | 0.68 A | 5.79 A |
| 1 cm | 53.4 AB | 6.16 AB | 2.03 A | 3.19 A | 0.72 A | 5.76 A |
| 2 cm | 51.5 AB | 5.61 B | 2.05 A | 3.09 A | 0.69 A | 5.72 A |
| 3 cm | 40.1 B | 5.58 B | 1.78 A | 2.83 A | 0.62 A | 5.91 A |
| Main effect of genotype | ||||||
| RTC-19 | 47.6 A | 5.5 B | 1.88 A | 3.11 A | 0.73 A | 5.67 A |
| RTC-119 | 54.5 A | 5.98 B | 1.9 A | 3.12 A | 0.7 A | 5.92 A |
| RTC-400 | 53.3 A | 6.5 A | 2.07 A | 2.78 B | 0.6 B | 5.79 A |
| 2019 | |||||
|---|---|---|---|---|---|
| Source of Variation (d.f.) | Seedling Density (No/m2) | Plant Height (cm) | Panicle Length (cm) | Grain Yield (g/m2) | Biomass (g/m2) |
| F ratio | |||||
| Sowing depth (SD) (1) | 5.07 * | 128.66 *** | 53.97 *** | 11.23 ** | 1.16 |
| Genotype (G) (1) | 0.53 | 0.05 | 0.55 | 6.2 * | 0.17 |
| SD*G interaction (1) | 0.52 | 4.72 * | 2.29 | 0.12 | 1.16 |
| Block (11) | 0.88 | 2.95 ** | 3.22 ** | 3.92 ** | 1.56 |
| Sowing depth main effect | |||||
| 1 cm | 201.96 A | 65.34 A | 44.23 A | 69.64 A | 1779 A |
| 3 cm | 173.25 B | 49.27 B | 34.94 B | 41.84 B | 1644 A |
| Genotype main effect | |||||
| RTC-119 | 182.96 A | 57.14 A | 39.12 A | 66.03 A | 1685 A |
| RTC-400 | 192.25 A | 57.47 A | 40.06 A | 45.46 B | 1737 A |
| 2020 | |||||
| Source of Variation (d.f.) | Seedling Density (No/m2) | Plant Height (cm) | Panicle Length (cm) | Grain Yield (g/m2) | Biomass (g/m2) |
| F ratio | |||||
| SD (1) | 5.36 * | 10.43 ** | 0.12 | 0.02 | 0.06 |
| G (2) | 10.79 ** | 0.46 | 1.22 | 23.56 *** | 0.63 |
| SD*G interaction (2) | 2.48 | 0.95 | 0.07 | 0.59 | 0.2 |
| Block (4) | 3.59 * | 0.38 | 3.07 * | 1.51 | 1.36 |
| Sowing depth main effect | |||||
| 1 cm | 226.74 A | 84.64 A | 36.29 A | 33.39 A | 1268 A |
| 3 cm | 190.35 B | 79.02 B | 35.59 A | 34 A | 1221 A |
| Genotype main effect | |||||
| RTC-19 | 256.65 A | 80.87 A | 37.1 A | 53.85 A | 1267 A |
| RTC-119 | 188.52 B | 82.9 A | 33.53 A | 16.65 C | 1099 A |
| RTC-400 | 180.45 B | 81.73 A | 37.18 A | 30.59 B | 1369 A |
| 2019 | |||||
|---|---|---|---|---|---|
| Source of Variation (d.f.) | LI 7 | LI 8 | LI 9 | LI 10 | LI All |
| F ratio | |||||
| Sowing depth (SD) (1) | 4.17 * | 4.17 * | 3.64 | 0.03 | 7.9 * |
| Genotype (G) (1) | 18.9 *** | 18.9 *** | 113.2 *** | 17.1 *** | 115.5 *** |
| SD*G interaction (1) | 0.03 | 0.03 | 0.18 | 0.09 | 0 |
| Block (11) | 4.3 *** | 4.3 *** | 10.6 *** | 2.1 | 14.8 *** |
| Week (3) | 17.51 *** | ||||
| Sowing depth main effect | |||||
| 1 cm | 466.4 A | 466.4 A | 607.22 A | 556.4 A | 516.49 A |
| 3 cm | 380.5 B | 380.5 B | 567.3 A | 563.0 A | 465.94 B |
| Genotype main effect | |||||
| RTC-119 | 330.2 B | 330.2 B | 473.4 B | 482.1 B | 393.65 B |
| RTC-400 | 516.7 A | 516.7 A | 701.1 A | 637.2 A | 588.79 A |
| 2020 | |||||
| Source of Variation (d.f.) | LI 7 | LI 8 | LI 9 | LI 10 | LI All |
| F ratio | |||||
| SD (1) | 4.2 | 1.8 | 0.1 | 0.1 | 4.1 * |
| G (1) | 0.52 | 6.1 * | 0.7 | 2.6 | 2.1 |
| G*D interaction (1) | 4.6 * | 2.2 | 0.9 | 5.1 * | 8.3 *** |
| Block (4) | 3.2 | 6.9 ** | 0.5 | 0.7 | 3.2 * |
| Week (3) | 19.4 *** | ||||
| Sowing depth main effect | |||||
| 1 cm | 701.7 A | 705.1 A | 839.56 A | 824.54 A | 767.8 A |
| 3 cm | 604.1 A | 642.7 A | 848.66 A | 820.41 A | 724.9 B |
| Genotype main effect | |||||
| RTC-19 | 644.7 A | 650 B | 862.1 A | 827.0A B | 739.22 A |
| RTC-119 | 688.0 A | 586.7 B | 821.3 A | 791.4 B | 723.67 A |
| RTC-400 | 626.0 A | 784.8 A | 848.9 A | 849.1 A | 776.16 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Zeev, S.; Kerzner, S.; Rabinovitz, O.; Saranga, Y. Optimizing Sowing Depth of Tef for Irrigated Mediterranean Conditions: From Laboratory to Field Studies. Agronomy 2020, 10, 1983. https://doi.org/10.3390/agronomy10121983
Ben-Zeev S, Kerzner S, Rabinovitz O, Saranga Y. Optimizing Sowing Depth of Tef for Irrigated Mediterranean Conditions: From Laboratory to Field Studies. Agronomy. 2020; 10(12):1983. https://doi.org/10.3390/agronomy10121983
Chicago/Turabian StyleBen-Zeev, Shiran, Shane Kerzner, Onn Rabinovitz, and Yehoshua Saranga. 2020. "Optimizing Sowing Depth of Tef for Irrigated Mediterranean Conditions: From Laboratory to Field Studies" Agronomy 10, no. 12: 1983. https://doi.org/10.3390/agronomy10121983
APA StyleBen-Zeev, S., Kerzner, S., Rabinovitz, O., & Saranga, Y. (2020). Optimizing Sowing Depth of Tef for Irrigated Mediterranean Conditions: From Laboratory to Field Studies. Agronomy, 10(12), 1983. https://doi.org/10.3390/agronomy10121983