Genetic Diversity and Population Structure of Algerian Endemic Plant Species Avena macrostachya Bal. ex Cross. et Durieu




Abstract
:1. Introduction
2. Materials and Methods
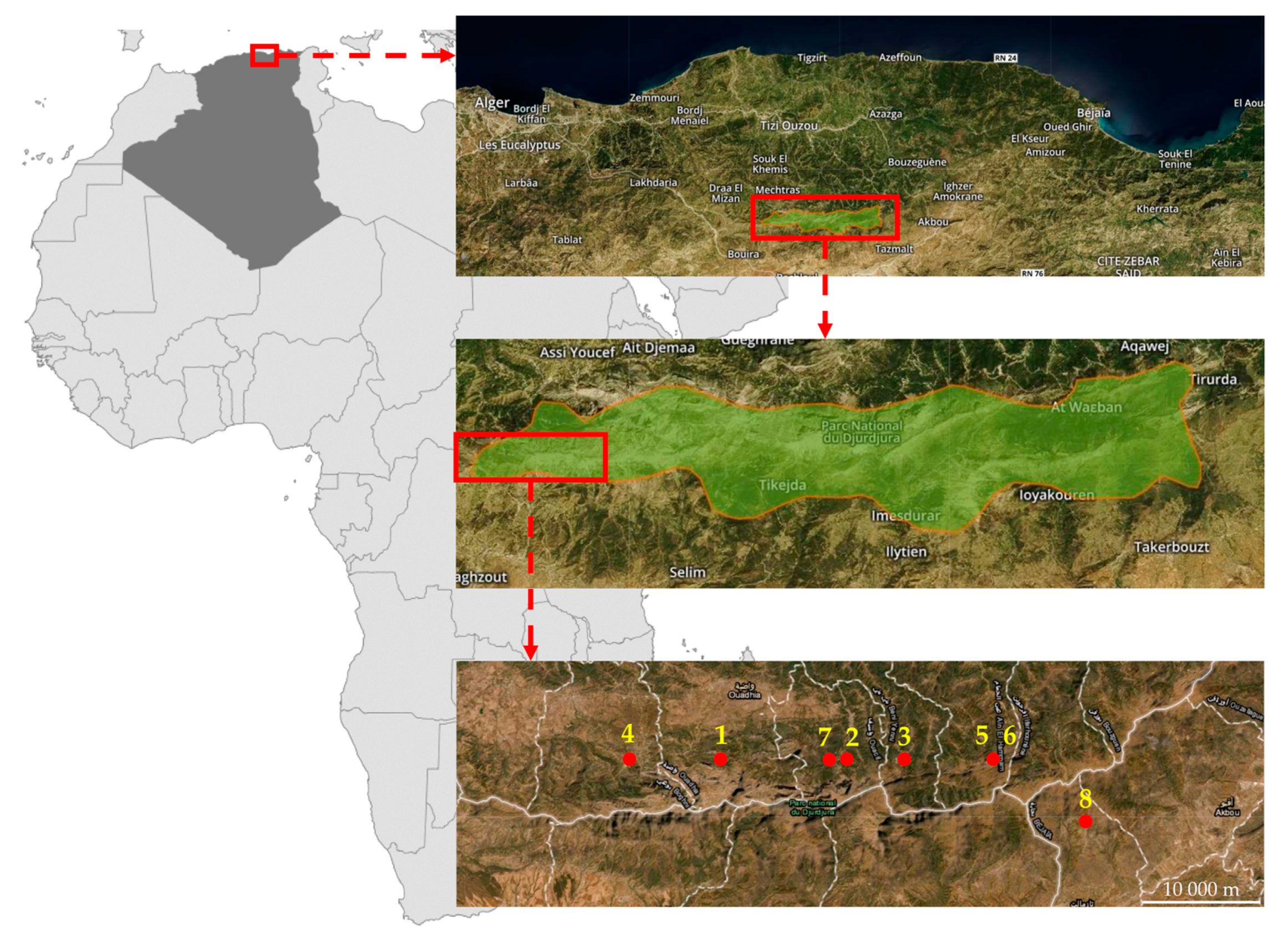
2.1. Plant Material
2.2. DNA Isolation
2.3. SRAP Analysis
2.4. Data Analysis
3. Results
3.1. Marker Informativeness
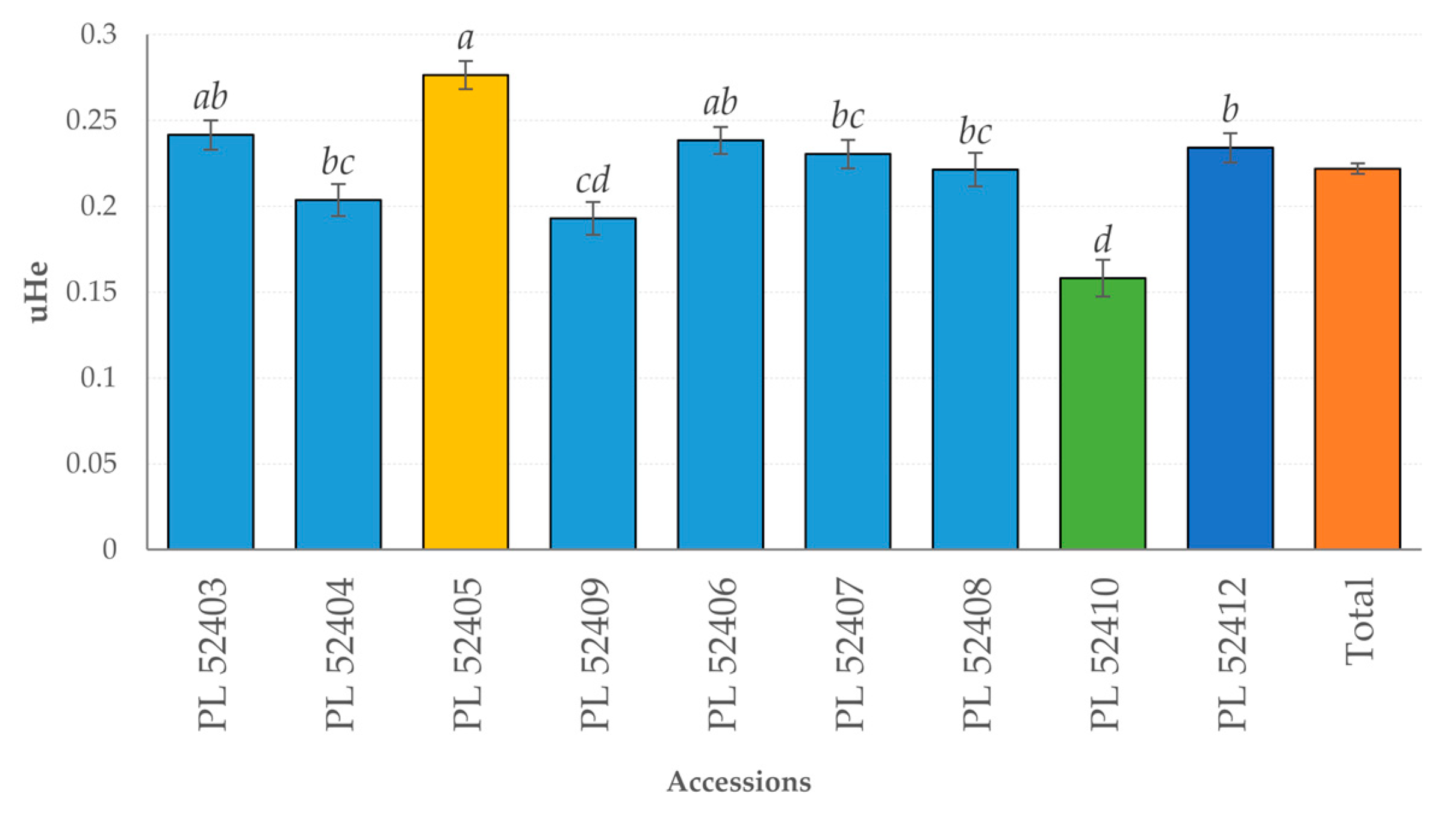
3.2. Genetic Diversity
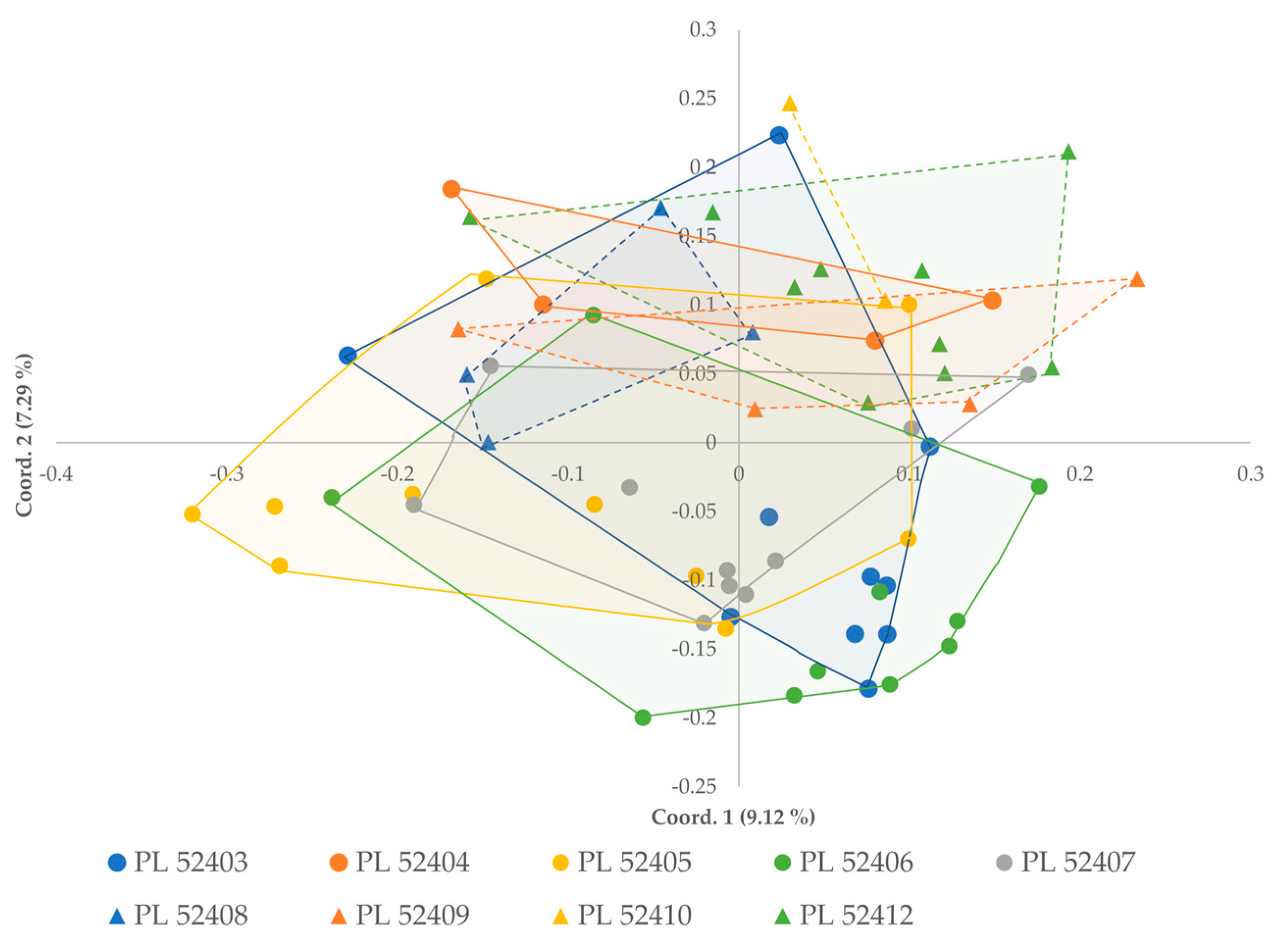
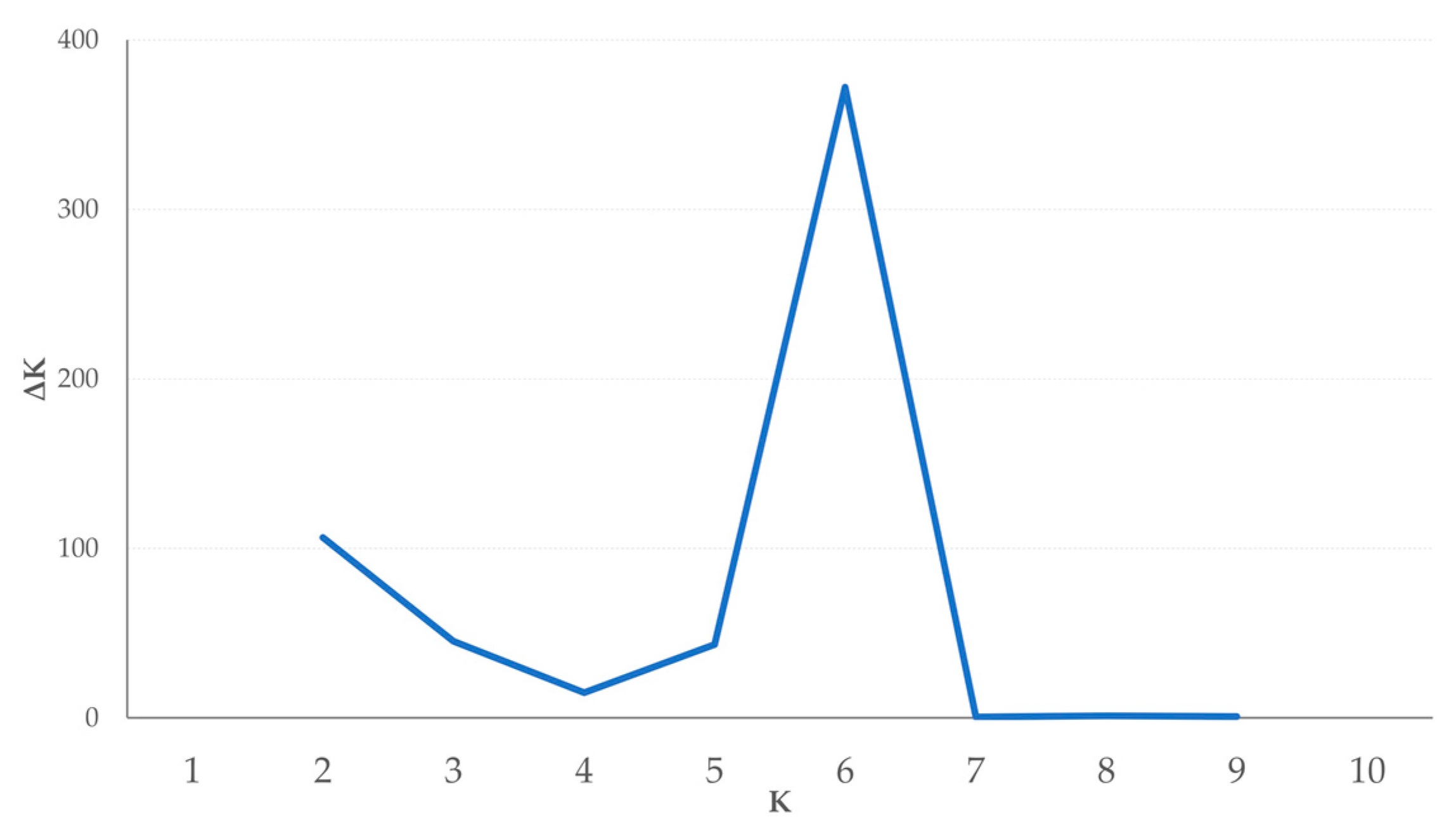
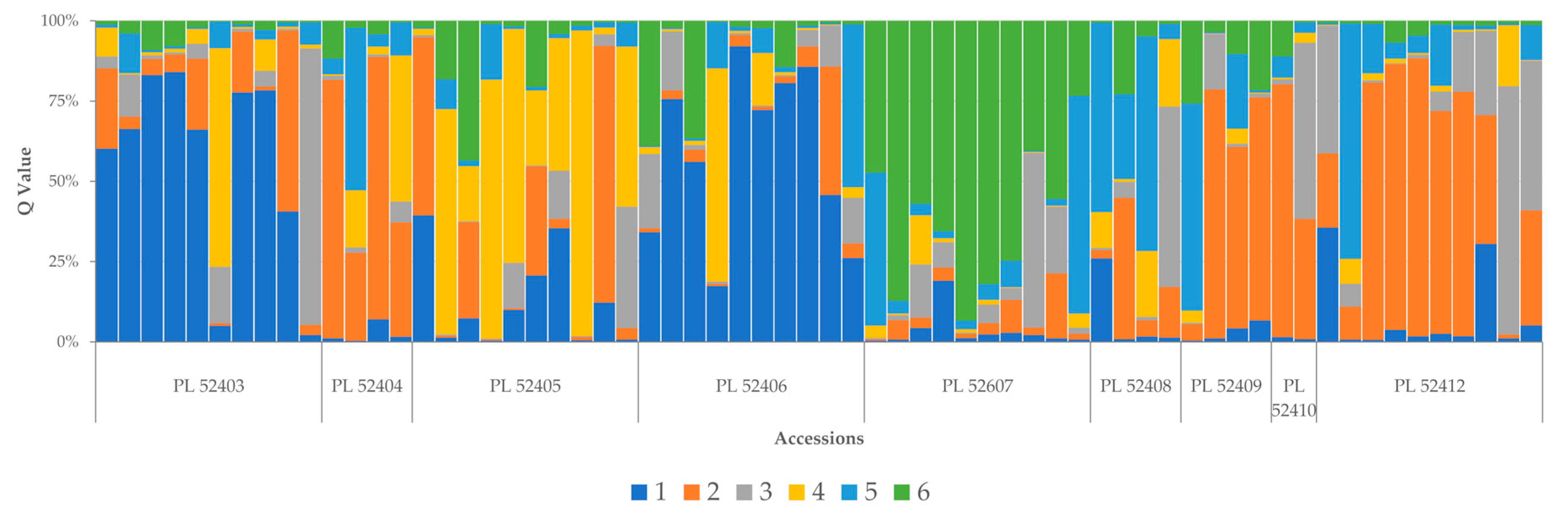
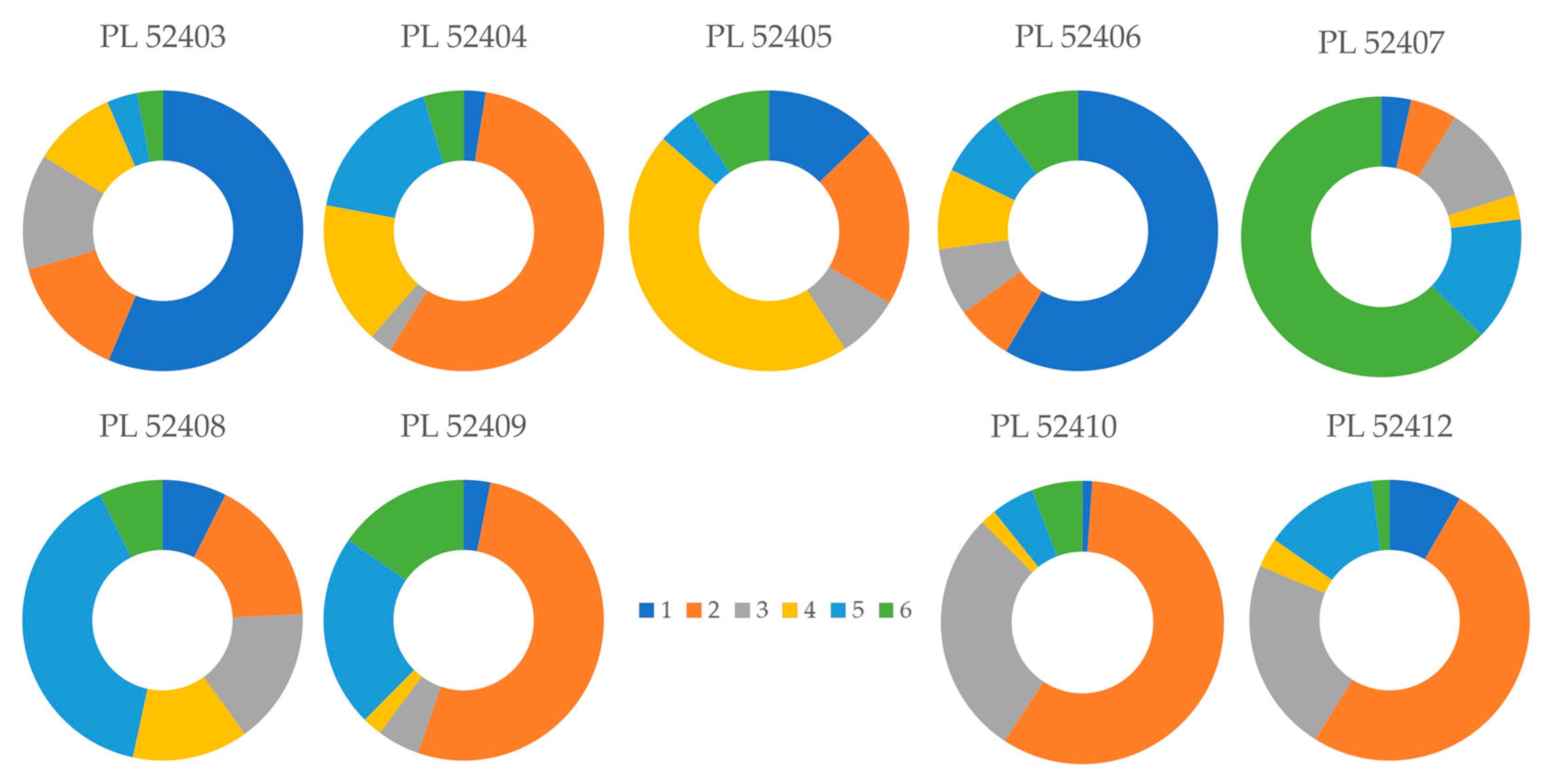
3.3. Population Structure
4. Discussion
4.1. Marker Effectivness
4.2. Genetic Diversity
4.3. Population Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Guarino, L.; Chadja, H.; Mokkadem, A. Collection of Avena Macrostachya Bal. Ex Coss. et Dur. (Poaceae) germplasm in Algeria. Econ. Bot. 1991, 45, 460. [Google Scholar] [CrossRef]
- Baum, B.R.; Rajhathy, T. A study of Avena macrostachya. Can. J. Bot. 1976, 54, 2434–2439. [Google Scholar] [CrossRef]
- Aissat, A.; Amirouche, R.; Amirouche, N. Cytotaxonomic investigation and meiotic behavior of natural populations of genus Avena in Algeria. Euphytica 2019, 215, 158. [Google Scholar] [CrossRef]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold Ltd.: London, UK, 1971. [Google Scholar]
- Pohler, W.; Hoppe, H.D. Homeology between the chromosomes of Avena macrostachya and the Avena C genome. Plant Breed. 1991, 106, 250–253. [Google Scholar] [CrossRef]
- Leggett, J. Further hybrids involving the perennial autotetraploid oat Avena macrostachya. Genome 1992, 35, 273–275. [Google Scholar] [CrossRef]
- Cheng, D.; Armstrong, K.; Drouin, G.; McElroy, A.; Fedak, G.; Molnar, S. Isolation and identification of Triticeae chromosome 1 receptor-like kinase genes (Lrk10) from diploid, tetraploid, and hexaploid species of the genus Avena. Genome 2003, 46, 119–127. [Google Scholar] [CrossRef]
- Rodionov, A.; Tyupa, N.; Kim, E.; Machs, E.; Loskutov, I. Genomic configuration of the autotetraploid oat species Avena macrostachya inferred from comparative analysis of ITS1 and ITS2 sequences: On the oat karyotype evolution during the early events of the Avena species divergence. Russ. J. Genet. 2005, 41, 518–528. [Google Scholar] [CrossRef]
- Peng, Y.-Y.; Wei, Y.-M.; Baum, B.R.; Zheng, Y.-L. Molecular diversity of the 5S rRNA gene and genomic relationships in the genus Avena (Poaceae: Aveneae). Genome 2008, 51, 137–154. [Google Scholar] [CrossRef]
- Altosaar, I.; Fabijanski, S. Molecular biology of oat storage protein genes: Probes for evaluating germplasm variation. In Proceedings of the Second International Oats Conference, Aberystwyth, UK, 15–18 July 1985; pp. 39–45. [Google Scholar]
- Baum, B.R.; Hadland, V. The epicuticular waxes of glumes of Avena: A scanning electron microscopic study of the morphological patterns in all the species. Can. J. Bot. 1975, 53, 1712–1718. [Google Scholar] [CrossRef]
- Clayton, W.D.; Vorontsova, M.S.; Harman, K.T.; Williamson, H. GrassBase—The Online World Grass Flora. Available online: http://www.kew.org/data/grasses-db/www/imp05260.htm (accessed on 25 May 2020).
- Medail, F.; Quezel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mol. Bot. Gard. 1997, 112–127. [Google Scholar] [CrossRef]
- Yahi, N.; Vela, E.; Benhouhou, S.; De Belair, G.; Gharzouli, R. Identifying important plants areas (key biodiversity areas for plants) in northern Algeria. J. Threat. Taxa 2012, 2753–2765. [Google Scholar] [CrossRef] [Green Version]
- Mediouni, K. Etude structurale de la série du cèdre à Aït-Ouabane Djurdjura. Forêt Méditerranéenne 1989, 11, 103–112. [Google Scholar]
- Dobignard, A.; Chatelain, C. Index Synonymique et Bibliographique de la Flore d’Afrique du Nord; ville de Genève editions des Conservatoire et Jardin botaniques, Genève, Switzerland 2010; Volumes 5. Available online: http://www.ville-ge.ch/musinfo/bd/cjb/africa/recherche.php (accessed on 10 September 2020).
- Rankou, H.; Culham, A.; Jury, S.L.; Christenhusz, M.J. The endemic flora of Morocco. Phytotaxa 2013, 78, 1–69. [Google Scholar] [CrossRef]
- Maire, R. Flore de l’Afrique du Nord (Maroc, Algérie, Tunisie, Tripolitaine, Cyrénaïque et Sahara); Lechevalier: Paris, France, 1952. [Google Scholar]
- GBIF.org. GBIF Occurrence. Available online: https://doi.org/10.15468/dl.u8hw78 (accessed on 18 November 2020).
- Boczkowska, M.; Tarczyk, E. Genetic diversity among Polish landraces of common oat (Avena sativa L.). Genet. Resour. Crop. Evol. 2013, 60, 2157–2169. [Google Scholar] [CrossRef] [Green Version]
- Boczkowska, M.; Podyma, W.; Łapiński, B. Oat. In Genetic and Genomic Resources for Grain Cereals Improvement; Elsevier: London, UK, 2016; pp. 159–225. [Google Scholar]
- Santos, A.; Livingston, D.; Murphy, J. Agronomic, Cytological and RAPD Evaluations of Avena Sativa x A. Macrostachya Populations. In Proceedings of the Eastern Wheat Workers and Southern Small Grain Workers Proceedings, Raleigh, NC, USA, 25 July 2002; Available online: http://www.ars.usda.gov/research/publications/Publications.htm (accessed on 2 September 2020).
- Łapinski, B.; Kała, M.; Nakielna, Z.; Jellen, R.; Livingston, D. The perennial wild species Avena macrostachya as a genetic source for improvement of winterhardiness in winter oat for cultivation in Poland. In Biotechnology and Plant Breeding—Perspectives; Behl, R.E.A., Ed.; Agrobios (International) Publishers: Jodphur, India, 2013; pp. 51–62. [Google Scholar]
- Yu, J.; Herrmann, M. Inheritance and mapping of a powdery mildew resistance gene introgressed from Avena macrostachya in cultivated oat. Theor. Appl. Genet. 2006, 113, 429–437. [Google Scholar] [CrossRef]
- Loskutov, I. Oat (Avena L.) Distribution, Taxonomy, Evolution and Breeding Value; VIR: Saint Petersburg, Russia, 2007. [Google Scholar]
- Weibull, J. Resistance in the wild crop relatives Avena macrostachya and Hordeum bogdani to the aphid Rhopalosiphum padi. Entomol. Exp. Applica. 1988, 48, 225–232. [Google Scholar] [CrossRef]
- Budak, H.; Shearman, R.; Parmaksiz, I.; Gaussoin, R.; Riordan, T.; Dweikat, I. Molecular characterization of buffalograss germplasm using sequence-related amplified polymorphism markers. Theor. Appl. Genet. 2004, 108, 328–334. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Resour. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poczai, P.; Varga, I.; Laos, M.; Cseh, A.; Bell, N.; Valkonen, J.P.; Hyvönen, J. Advances in plant gene-targeted and functional markers: A review. Plant Methods 2013, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Amar, M.H.; El-Zayat, M.A.S. Utilization of ISTR, ISSR and SRAP molecular markers to reveal and classify Egyptian pomegranates (‘Punica granatum’ L.). Plant Omics 2017, 10, 237. [Google Scholar] [CrossRef]
- Podyma, W.; Bolc, P.; Nocen, J.; Puchta, M.; Wlodarczyk, S.; Lapinski, B.; Boczkowska, M. A multilevel exploration of Avena strigosa diversity as a prelude to promote alternative crop. BMC Plant Biol. 2019, 19, 291. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.A.; Churchill, G.A.; Autrique, J.E.; Tanksley, S.D.; Sorrells, M.E. Optimising parental selection for genetic linkage maps. Genome 1992, 36, 181–186. [Google Scholar] [CrossRef]
- Podyma, W.; Boczkowska, M.; Wolko, B.; Dostatny, D.F. Morphological, isoenzymatic and ISSRs-based description of diversity of eight sand oat (Avena strigosa Schreb.) landraces. Genet. Resour. Crop. Evol. 2017, 64, 1661–1674. [Google Scholar] [CrossRef]
- Boczkowska, M.; Łapiński, B.; Kordulasińska, I.; Dostatny, D.F.; Czembor, J.H. Promoting the use of common oat genetic resources through diversity analysis and core collection construction. PLoS ONE 2016, 11, e0167855. [Google Scholar] [CrossRef] [Green Version]
- Boczkowska, M.; Onyśk, A. Unused genetic resources: A case study of Polish common oat germplasm. Ann. Appl. Biol. 2016, 169, 155–165. [Google Scholar] [CrossRef]
- Sales, E.; Nebauer, S.G.; Mus, M.; Segura, J. Population genetic study in the Balearic endemic plant species Digitalis minor (Scrophulariaceae) using RAPD markers. Am. J. Bot. 2001, 88, 1750–1759. [Google Scholar] [CrossRef] [PubMed]
- Pecinka, A.; Fang, W.; Rehmsmeier, M.; Levy, A.A.; Scheid, O.M. Polyploidization increases meiotic recombination frequency in Arabidopsis. BMC Biol. 2011, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budak, H.; Shearman, R.C.; Gulsen, O.; Dweikat, I. Understanding ploidy complex and geographic origin of the Buchloe dactyloides genome using cytoplasmic and nuclear marker systems. Theor. Appl. Genet. 2005, 111, 1545–1552. [Google Scholar] [CrossRef]
- Gulsen, O.; Shearman, R.C.; Vogel, K.P.; Lee, D.J.; Baenziger, P.S.; Heng-Moss, T.M.; Budak, H. Nuclear genome diversity and relationships among naturally occurring buffalograss genotypes determined by sequence-related amplified polymorphism markers. Hort. Sci. 2005, 40, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Gulsen, O.; Sever-Mutlu, S.; Mutlu, N.; Tuna, M.; Karaguzel, O.; Shearman, R.C.; Riordan, T.P.; Heng-Moss, T.M. Polyploidy creates higher diversity among Cynodon accessions as assessed by molecular markers. Theor. Appl. Genet. 2009, 118, 1309–1319. [Google Scholar] [CrossRef] [Green Version]
- Allaby, R.G.; Ware, R.L.; Kistler, L. A re-evaluation of the domestication bottleneck from archaeogenomic evidence. Evol. Appl. 2019, 12, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Loveless, M.D.; Hamrick, J.L. Ecological determinants of genetic structure in plant populations. Annu. Rev. Ecol. Evol. Syst. 1984, 15, 65–95. [Google Scholar] [CrossRef]
- Francisco-Ortega, J.; Santos-Guerra, A.; Kim, S.C.; Crawford, D.J. Plant genetic diversity in the Canary Islands: A conservation perspective. Am. J. Bot. 2000, 87, 909–919. [Google Scholar] [CrossRef] [Green Version]
- Bolibok-Brągoszewska, H.; Targońska, M.; Bolibok, L.; Kilian, A.; Rakoczy-Trojanowska, M. Genome-wide characterization of genetic diversity and population structure in Secale. BMC Plant Biol. 2014, 14, 184. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Huang, J.; Yin, X.; Lian, C.; Li, X. Genetic diversity and population structure of sour jujube, Ziziphus acidojujuba. Tree Genet. Genomes 2015, 11, 809. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, M.; Guo, Z.; Guan, Y.; Liu, J.; Yan, X.; Guo, Y. Genetic Diversity and Population Structure of Bermudagrass (Cynodon dactylon (L.) Pers.) along Latitudinal Gradients and the Relationship with Polyploidy Level. Diversity 2019, 11, 135. [Google Scholar] [CrossRef] [Green Version]
- Shang, H.-Y.; Wei, Y.-M.; Wang, X.-R.; Zheng, Y.-L. Genetic diversity and phylogenetic relationships in the rye genus Secale L.(rye) based on Secale cereale microsatellite markers. Genet. Mol. Biol. 2006, 29, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xie, W.; Wang, Y.; Zhao, X. Potential of start codon targeted (SCoT) markers to estimate genetic diversity and relationships among chinese Elymus sibiricus accessions. Molecules 2015, 20, 5987–6001. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1949, 15, 323–354. [Google Scholar] [CrossRef]
- Wright, S. Isolation by distance under diverse systems of mating. Genetics 1946, 31, 39. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession Number | Collection Site | Latitude | Longitude | Acquisition | Number of Plant |
|---|---|---|---|---|---|---|
| 1 | PL 52403 | Tikjda, 6 km W (Djurdjura) | N 36°30′ | E 4°5′ | expedition [1] | 10 |
| 2 | PL 52404 | Tikjda, 3 km E (Djurdjura) | N 36°30′ | E 4°12′ | expedition [1] | 4 |
| 3 | PL 52405 | Tikjda, 8 km E (Djurdjura) | N 36°30′ | E 4°15′ | expedition [1] | 10 |
| 4 | PL 52406 | Tala Guilef (Djurdjura) | N 36°30′ | E 4°0′ | expedition [1] | 10 |
| 5 | PL 52407 | Tirourda (Djurdjura) | N 36°30′ | E 4°20′ | expedition [1] | 10 |
| 6 | PL 52408 | Col de Tirouda (Djurdjura) | N 36°30′ | E 4°20′ | expedition | 4 |
| 7 | PL 52409 | (Djurdjura) | N 36°30′ | E 4°11′ | Expedition NCPGR | 4 |
| 8 | PL 52410 | Tikjda (Djurdjura) | N 36°27′ | E 4°25′ | exchange PGRC CAV 5264 [2] | 2 |
| 9 | PL 52412 | n.a. | n.a. | n.a. | open-pollination PL 52411 and PL 52409 | 10 |
| Primer | Sequence | Dye | |
|---|---|---|---|
| Forward | Me1 | TGA GTC CAA ACC GGA TA | 6FAM |
| Me2 | TGA GTC CAA ACC GGA GC | VIC | |
| Me3 | TGA GTC CAA ACC GGA AT | NED | |
| Me4 | TGA GTC CAA ACC GGA CC | PET | |
| Reverse | Em3 | GAC TGC GTA CGA ATT GAC | - |
| Em4 | GAC TGC GTA CGA ATT TGA | - | |
| Em6 | GAC TGC GTA CGA ATT GCA | - | |
| Em7 | GAC TGC GTA CGA ATT CAA | - | |
| Em8 | GAC TGC GTA CGA ATT CAC | - | |
| Em13 | GAC TGC GTA CGA ATT CTG | - |
| Set | Forward | Reverse |
|---|---|---|
| 1 | Me1 | Em4 |
| Me2 | Em3 | |
| Me3 | Em3 | |
| Me4 | Em8 | |
| 2 | Me1 | Em13 |
| Me2 | Em7 | |
| Me3 | Em6 | |
| Me4 | Em7 |
| Primers | No. of Fragments | % of Polymorphic Fragments | % of Rare Fragments | PIC |
|---|---|---|---|---|
| Me1/Em4 | 113 | 78% | 17% | 0.269 |
| Me1/Em13 | 93 | 69% | 20% | 0.233 |
| Me2/Em3 | 109 | 81% | 17% | 0.270 |
| Me2/Em7 | 102 | 70% | 22% | 0.228 |
| Me3/Em3 | 93 | 67% | 31% | 0.221 |
| Me3/Em6 | 107 | 66% | 33% | 0.223 |
| Me4/Em8 | 78 | 56% | 35% | 0.171 |
| Me4/Em7 | 86 | 67% | 26% | 0.204 |
| Mean | 97.6 | 69.3% | 27.8% | 0.227 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolc, P.; Łapiński, B.; Podyma, W.; Boczkowska, M. Genetic Diversity and Population Structure of Algerian Endemic Plant Species Avena macrostachya Bal. ex Cross. et Durieu. Agronomy 2020, 10, 1984. https://doi.org/10.3390/agronomy10121984
Bolc P, Łapiński B, Podyma W, Boczkowska M. Genetic Diversity and Population Structure of Algerian Endemic Plant Species Avena macrostachya Bal. ex Cross. et Durieu. Agronomy. 2020; 10(12):1984. https://doi.org/10.3390/agronomy10121984
Chicago/Turabian StyleBolc, Paulina, Bogusław Łapiński, Wiesław Podyma, and Maja Boczkowska. 2020. "Genetic Diversity and Population Structure of Algerian Endemic Plant Species Avena macrostachya Bal. ex Cross. et Durieu" Agronomy 10, no. 12: 1984. https://doi.org/10.3390/agronomy10121984
APA StyleBolc, P., Łapiński, B., Podyma, W., & Boczkowska, M. (2020). Genetic Diversity and Population Structure of Algerian Endemic Plant Species Avena macrostachya Bal. ex Cross. et Durieu. Agronomy, 10(12), 1984. https://doi.org/10.3390/agronomy10121984