1. Introduction
The spectral composition of solar radiation plays one of the main informative and signaling roles in plant development. The possibility of monitoring the number, quality and direction of radiation changes allows plants to optimize light energy absorption, which is necessary for photosynthesis. Using transmitted and reflected radiation, they detect the presence of neighboring plants and then create appropriate competitive responses, allowing them to get an advantage over the other plants. The capability of plants to detect the presence of neighboring plants has caused the development of some mechanisms that allow them to survival in adverse environmental conditions. One of the important stress factors for plants is shading; however, they have two ways of doing this: avoid it or tolerate it.
The avoidance of shading is one of the more necessary competitive strategies. The ability to detect and respond to upcoming shading is an important feature of plants growing in natural environments. Competition between single individuals is a result of the response to light shortage and the spectral composition of radiation. One of the main signals of shading is a decrease of the red (R) to far red (FR) ratio in transmitted radiation reflected from neighboring plants [
1,
2,
3]. Shade avoidance syndrome (SAS) [
2,
4,
5] is frequently caused by FR reflected from neighboring plants, even before the growth of the stand, which means that plants can detect and react to potential future competition just before the shading [
1,
6,
7,
8]. Natural shading in stands give a low R/FR ratio [
9,
10] when compared to natural solar radiation in an open space [
11,
12] because a high absorption of chlorophyll and other photosynthetic pigments in the range of the blue and red spectrum causes the absorption of radiation in this range to be high, while in the range above 700 nm it is relatively low.
The detection of neighboring plants is possible by radiation from the range of the blue spectrum [
5,
13,
14], which, to a great degree, is reduced in its absorption effect by competitive plants. An important signal for avoiding shading is also a low value of radiation, especially in the range of Photosynthetically Active Radiation (PAR) [
1,
5,
15]. An important element in the protection strategy against unfavorable conditions is the ability to receive light signals by photoreceptors. Phytochromes belong to the more important among them.
In the angiosperms, in the most numerous group of plants in the world, three main phytochromes generally occur: phyA, phyB and phyC [
15].
The phyA is a photoreceptor registering FR presence in the environment and then mediating photo-morphogenetic reactions as a response to a signal from this spectrum range [
16]. PhyB strongly reacts to changes of the R/FR ratio [
16,
17], and even its small changes cause great photo-morphogenetic effects [
18]. Under conditions of radiation, a reduction of phyB occurs with a low R/FR ratio, a form of phytochrome that absorbs far red (P
fr), and at the same time the initiation of a reaction referred to as shading avoidance syndrome (SAS) follows. PhyB plays a dominant role in mediating other neighbors’ plants detection [
5] and shading avoidance reactions [
15].
Phytochromes A and B perform a very important function, which consists in the mediation of radiation (in the range of red) in the promotion of stomata production [
19]. The role of the phytochrome D (phyD) is to evaluate changes in the R/FR ratio. It was found that phyE mediated the reaction of shading avoidance and responses to the R/FR ratio [
20,
21,
22,
23,
24,
25].
With the exception of performing the role of radiation detection, a phytochrome is a key player, playing decidedly the most important role in the shading avoidance process. In 1982, Smith [
4] put forward a hypothesis about the perception of the R/FR ratio as a fundamental function of a phytochrome. A plant evaluates the presence of a neighbor by monitoring a signal reflected from green tissues, characterized by a greater share of far red in relation to red. Changes in the amount of far red are correlated with the plant’s density and proximity to neighboring plants [
6,
7,
26,
27]. The R/FR ratio in the stand decreases together with the density of green plants [
26,
28,
29]. Through the perception of the R/FR ratio via the phytochrome, plants determine the degree of shading. In response to this signal, plants trigger shading avoidance reactions. A relatively small increase of the FR amount causes an enlarged elongation growth and even a negative phototropic response (as a bending over from neighbors’ plants) [
5]. PhyB is mostly responsible for the plant’s „shade avoidance response” to a low R/FR ratio [
4], and to a lesser degree phyD and phyE take part in this reaction [
15,
16,
25]. It is particularly emphasized that even small changes in the R/FR ratio cause considerable changes in photo-morphogenesis [
27].
It should also be noted that under the conditions of a radiation with a low R/FR ratio, the activity of auxins (IAA19) increases considerably, which also takes part in the process of shading avoidance by interacting with DELLA protein [
28].
Radiation in the blue range also plays a significant role in the photo-morphogenesis. A little amount of this radiation determines some photo-morphogenetic reactions [
25,
29].
Radiation from the range of R is absorbed by the chlorophyll, whereas radiation from the range FR is reflected and transmitted by the leaves, causing a considerable increase in the FR amount in the surrounding of the plants [
30]. The first signal indicated by the presence of neighboring plants is generated at the moment when the FR participation increases. Together with plant growth, leaves’ density decreases and the reflected radiation in the range of the FR increases, so consequently a lowering of the R/FR ratio occurs.
The enrichment of radiation with the additional FR during a few minutes before the dark period caused the number of stomata on the lower and upper side of the tobacco leaves to be lower than for plants that were lit by an additional amount of R [
31]. The biggest degree of stomata opening was observed after radiation by the blue and red parts of the spectrum, and using a system in which plants were first treated with the radiation ranges R and B and then FR caused a lower degree of their opening [
32].
A few minutes of exposure of FR before the dark period causes chloroplasts in the tobacco leaves to contain a lower granum than after a similar treatment with red or full white light [
33,
34].
The aim of this research, led under precisely controlled conditions, was the determination of the spring barley response to different spectral compositions, especially in the range of R and FR.
2. Materials and Methods
The experiments were conducted under control conditions in two climatic chambers, HERAEUS firm, with the use of artificial sources of radiation. In each of these chambers different spectral compositions of radiation were used through the use of bulbs and fluorescent lamps from the Philips company and halogen lamps of the PAR30 rosé type form the Paulmann company (
Table 1).
The selection of the radiation sources under these conditions with proper spectral characteristics of radiation was led with the use of spectroradiometric measurements. The distance between the fluorescent lamps, Philips bulb, Paulmann lamp and spectroradiometer’s sensor amounted to 20 cm (
Figure 1 and
Figure 2). In both chambers, radiation from fluorescent lamp from Philips company (58 W) was used. Moreover, in the A chamber, additional radiation originating from Philips company bulbs (60 W) was used, and in the B chamber radiation from halogen lamps of type PAR 30 rosé (75 W) from the Paulmann company was used. The temperature, air moisture, and length of a day and a night were the same in both chambers. In the studies, we used the day and night cycle occurring under natural conditions. The experiments were started under the conditions of a sunrise and sunset simulation, which occurred in Puławy (51°24′46” N, 21°58′00” E) on 26 March when the length of day/night amounted to 12 h and 30 min/11 h and 30 min (while for the longest day of the year—21 June—it amounted to 16 h and 38 min/7 h and 22 min), and at the end of vegetation the length of the day/night reached 15 h and 30 min/8 h and 30 min. For every following day, the length of the day was increasing and the length of the night was decreasing. During the whole period of experimentation, the air temperature was changing in the chambers respectively to the temperature occurring under natural conditions in Puławy in the months of the year following March 26, and thus in the beginning of the experiment the mean day/night temperature amounted to 7.5/2.5 °C, while in April it was 12/4 °C, in May it was 19.4/7.8 °C, in June it was 24.0/10.0 °C, and in July it was 25.9/11.3 °C.
2.1. Spectral Composition of Radiation
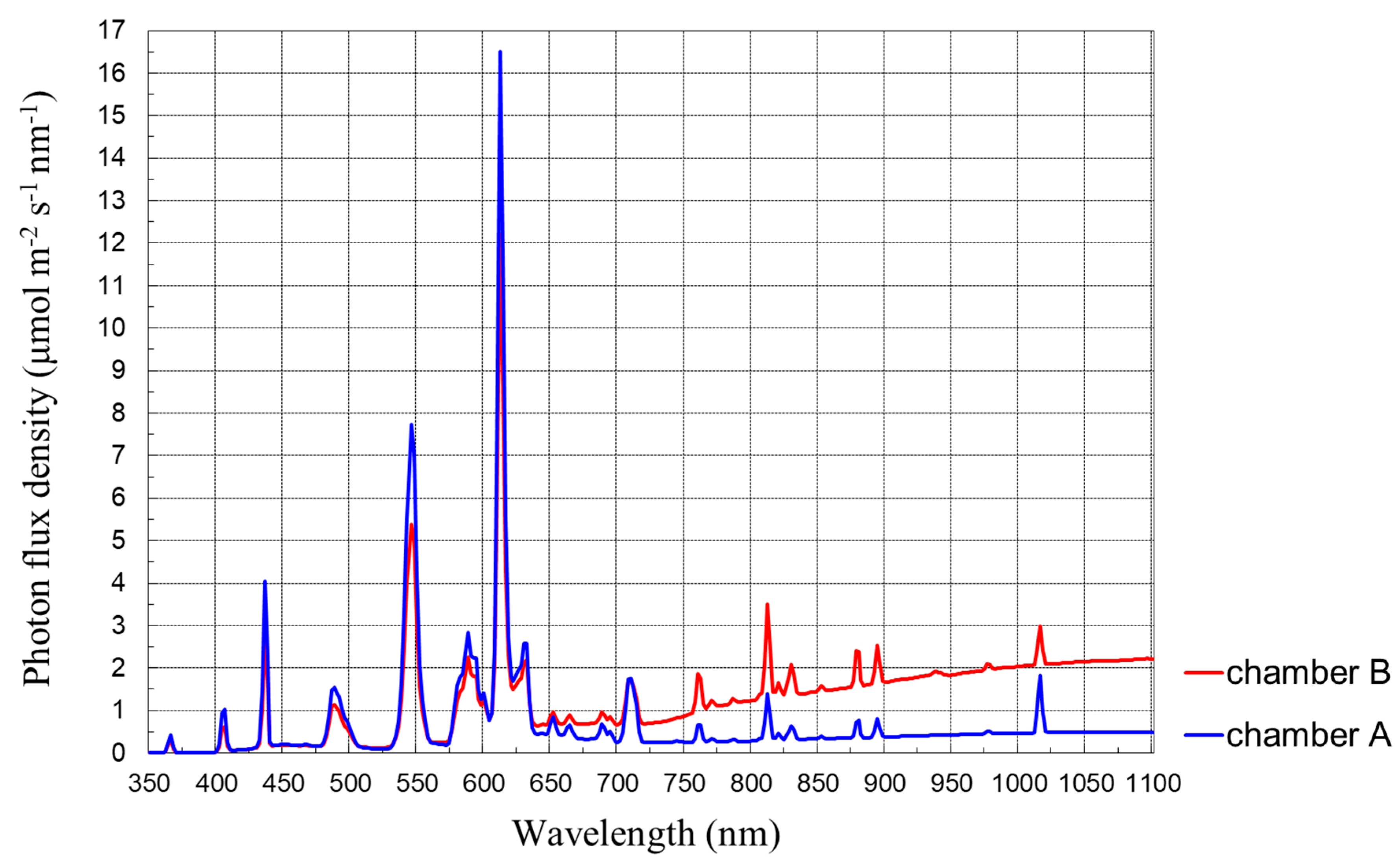
The spectral composition of the radiation was measured with the use of a spectrometer LI-1800, which worked in the range of 350–1100 nm, with a half-width of 4 nm. Radiation originating from the fluorescent lamps was characterized by the occurrence of many peaks (
Figure 1). In the range of photosynthetic active radiation (400–700 nm), three main maxima occurred: about 430–440 nm (range violet-blue), 540–560 nm (green-yellow) and 610–620 nm (orange, as well as three smaller ones: 480–490 nm (blue-green), 580–590 nm (yellow) and about 630 nm (red) (
Figure 1 and
Figure 2).
To obtain a similar amount of photosynthetic active radiation (PAR 400–700 nm) to reach the plants (the density of photon stream PAR was about 300 µmol m
−2 s
−1), fluorescent lamps, bulbs and lamps were placed at a differentiated distance under the plants (
Table 1). The spectral characterization of PAR is presented in
Figure 2.
The spectral composition of the Philips bulb and Paulmann halogen lamp radiation shows the significant participation of long-term radiation (
Figure 3). These sources of radiation are characterized by an increase of irradiation as the wavelength increases from the range of violet to near infrared. However, the increase of radiation above 563 nm (in the range of yellow, orange and red) is considerably greater for Paulmann lamps than for Philips bulbs.
After the application of a properly chosen set of fluorescent lamps, bulbs and lamps, and a differentiated distance of plants from the source of radiation, we obtained various compositions of radiation in each chamber (
Figure 4).
The Paulmann type PAR30 rosé lamps emitted radiation that was characterized by a big differentiation between the range of 600 and 700 nm, considerably greater than that occurring in the radiation of the Philips fluorescent lamps. The use of lamps with different spectral compositions led to us obtaining the needed conditions of radiation, which showed a differentiated R/FR ratio in the particular chambers. The values of the R/FR ratio at the radiation used in the growth chambers are shown in
Table 2.
The integrated values of radiation in the selected spectral ranges used in the chambers are shown in
Table 3. Radiation below 400 nm was similar in both chambers, whereas differences occurred for radiation above 400 nm. The greatest radiation in the spectral ranges 400–499 and 500–599 nm occurred in chamber A, and it was a little smaller in chamber B. Meanwhile, the wavelength increased above 600 nm, so the differences in radiation between both chambers were larger and larger; however, a smaller radiation occurred in the chamber A and a greater occurred in the B one.
The use of different sources of radiation caused very big differences in the amounts of radiation in the range of 700–1100 nm, which is a considerable part of the radiation that is far red (780–1400 nm). The biggest radiation in this range occurred in chamber B, and a smaller one occurred in the A one. The differences in the amounts of radiation in the visible range (about 380–750 nm) between chambers were relatively small, especially in the range of short waves: violet, blue, green, yellow and orange. The greatest radiation values from the visible range occurred in chamber A, and they were smaller for chamber B. In the growth and development of plants, the most important radiation is radiation from the photosynthetic active range—PAR. Radiation in the wavelength exceeding 700 nm, especially long-wave radiation—near infrared (NIR, 700–1100 nm)—has a smaller effect on growth parameters because in this range both the transmission and reflection of radiation are very big in comparison to PAR, amounting in each case to more than 45%. This results in plants’ absorption of radiation in this range only reaching some percentages.
In
Figure 5, the differences in the spectral composition of radiation in the growth chambers are visible(
Figure 5). Greater differences occurred for the red and purple color (far red) (
Figure 4,
Table 2).
The radiation in the ranges of 620–699 and 700–759 nm was decidedly lower in chamber A than in chamber B. This difference was caused by the very low emission of fluorescent lamps in these ranges of radiation and, at the same time, the very high emission of Paulmann halogen lamps in the longwave part of the spectrum.
In the range of radiation of 650–750 nm, one high pick with a maximum at 710 nm occurred, as well as four lower picks at 653, 666, 688 and 695 nm (
Figure 6).
2.2. Research Material
A subject of the study was spring barley cv. Rastik. Five grains of barley were sown in every Mitscherlich pot and filled with 3 kg of substrate based on high peat (sphagnum peat). The pHH2O of this substrate was 5.5–6.5, and the concentrations of N, P (P2O5) and K (K2O) were in a ratio of 14:16:18, respectively. It also contained some amounts of MgO, as well as microelements: Fe, Mn, Cu, Zn, Mo and B. The dry mass of the substrate amounted to 35%, and the organic substance amounted to 85%. On the third day after emergence, plants were tinned, and one plant was left in each pot. In both growth chambers, 24 Mitscherlich pots were placed.
Measurements of photosynthesis, the intracellular concentration of CO2 and stomatal conductance were performed with the use of the LI 6400 (LI-COR). The photosynthetic activity of plants was measured two times: first, during the phase of 7 leaves (measurement on the sixth leaf) and, second, during the phase of the fully developed flag leaf of barley (measurement on the flag leaf).
Transpiration was determined with the use of water use efficiency (WUE) as an indicator of the water efficiency at photosynthesis, which characterizes gas exchange in the assimilation organs:
where: Pn—photosynthesis (µmol CO
2 m
−2s
−1) and E—transpiration (mmol H
2O m
−2s
−1).
The number of stomata was determined using an optical microscope (Nikon company) at a 400-fold enlargement in the field of view.
The chlorophyll content in the leaves was determined with the use of the optical device HYDRO N-tester (Minolta 502), which measures the difference between the absorption of radiation at wavelength 650 nm (maximum absorption of radiation in the range of red) and 940 nm (radiation transmitted by the leaf tissue). The average chlorophyll content was counted on the basis of 30 properly performed measurements on the leaves of the same plant. The measurement result is given in SPAD units (Soil Plant Analysis Development) on a scale from 1 to 800.
The average values of the studied plant features were compared with the statistical program STATGRAPHYCS Plus v. 2.1 for Windows with the use of a multiple comparison test (Multiple Range Tests).
4. Discussion
The studies conducted on the effect of far red and red showed the significant effect of radiation conditions on the physiological processes of plants. The obtained results fully proved that barley plants responded very strongly to the spectral composition of radiation [
12]. Among the tested species of cereal plants, the strongest response to an enlarged share of radiation in the FR range was shown by spring wheat [
12]. The studies conducted by authors on spring barley also indicated that this species belongs to the group of plants showing a strong response to relations between R and FR. Undoubtedly, the observed cereal’s response fully deserves to be named as an SAS reaction occurring in plants growing in dense stands, although there was a lack of competitive conditions. These aspects are the results of radiation conditions, which invoke certain physiological processes, and which are, to a great extent, determined by the photosynthetic effect of gas exchange and CO
2 assimilation. The volume of gas exchange that consists of photosynthesis, stomatal respiration and the WUE index is dependent, among others, on spectral composition and radiation intensity [
35]. Plant responses such as the shoot elongation, height of plant and internodes are more connected to the spectral composition than to the radiation intensity. A very similar amount of radiation was used in the studies conducted in the growth chambers, so it can be expected that the main differential element of the plant behavior was the spectral composition of the radiation. The plant responses under the different radiation conditions were respective processes connected to gas exchange and water management.
Photosynthesis efficiency, expressed as the CO
2 gas exchange of sixth and flag leaf of barley, showed significantly higher values in objects with a high R/FR ratio than in those with a low R/FR ratio. This indicates that plants that were growing under radiation conditions with a relatively low R/FR ratio led a more economical CO
2 consumption. Similarly, for tobacco irradiated with a low R/FR ratio, the net assimilation of CO
2 was lower than after irradiation with a high R/FR ratio [
30]. Sleeman et al. also reported about the greater photosynthesis efficiency after the use of radiation with a high R/FR ratio = 10.0 [
35]. The photosynthesis efficiency measured by CO
2 gas exchange in
Mercurialis annua—a plant from the
Euphorbiaceae family—under radiation conditions with an R/FR ratio = 1.1–1.4 was lower, similar to the experiment with wheat [
36] and to the present experiment with barley (chamber B). Photosynthesis efficiency is a physiological process that is very sensitive to stress conditions. Under an optimal soil moisture, CO
2 was assimilated by grasses to a considerably greater degree than under a water deficit [
37].
Besides the energy expressed as the amount of absorbed photons, the second very important factor that enables the course of photosynthesis is the supply of CO
2, which is the intracellular concentration of carbon dioxide (CO
2). The intracellular concentration of carbon dioxide was considerably higher in the wheat [
36] and barley, which grow under radiation conditions with a high R/FR ratio. The obtained result means that plants growing in chambers with a high R/FR ratio had better access to carbon dioxide, which caused the increase of their biomass production. The increase of starch production in the leaves of tomato was reported by Czarnowski and Starzecki, among others [
38].
Another physiological aspect is water management. Barley growing in chambers with a lower R/FR ratio showed a considerably lower consumption of water than plants growing at a high ratio. The response of wheat plants was similar, although less spectacular [
36]. On this basis, it can be assumed that these responses are undoubtedly a manifestation of adaptation of plants growing under radiation conditions with a low R/FR ratio to restrict water consumption with regards to expected competition or droughts because a lower R/FR ratio occurs in a dry atmosphere and a higher one occurs in an atmosphere with a lot of moisture [
11]. Similar results were obtained in relation to the transpiration of cotton and
Abutilon theophrasti plants, which also showed a lower use of water at a low R/FR ratio than under radiation with a higher value of this ratio [
39,
40]. The mechanisms by which a phytochrome influences the water economy may be more or less complicated [
41,
42,
43]. Some recent reports showed that the phytochrome increased tolerance to a high evaporative demand [
44] and that phytochromes A and B could modulate drought stress responses [
45].
A synthetic indicator that describes water transpiration from plants and photosynthesis efficiency is water use efficiency (WUE), also known as the index of water efficiency under photosynthesis. It characterizes the gas exchange in the assimilation organs and water consumption. In the conducted studies with the velvet leaf plants, a small increase of WUE values treated with a high R/FR ratio was obtained, but the differences were too little to be significant [
35]. Considerably higher WUE values were obtained for barley plants growing at a radiation with a low R/FR ratio when compared to a radiation with a higher ratio. These results indicated that in competitive conditions (with a low R/FR ratio), the efficiency of water consumed under barley photosynthesis was considerably higher than in non-stressful conditions (with a high R/FR ratio). In the present studies, higher WUE values also occurred for barley plants subjected to drought stress. Similar results were obtained by Staniak with four species of fodder grasses under competitive conditions connected with water deficit [
37].
Barley conducted water to a much greater extent under conditions with a high than with a low R/FR ratio. The obtained stomatal conductance results indicate that plants growing under radiation conditions with a low share of FR (which shows a lack of competition) conducted considerably more water than plants growing under conditions with a higher share of radiation from this range, i.e., under competitive conditions. Very similar results concerning the stomatal conductance were obtained by Sleeman et al. [
35] with
Mercurialis annua plants during experiments with the same conditions of radiation and by Staniak [
37] with plants growing under water shortage.
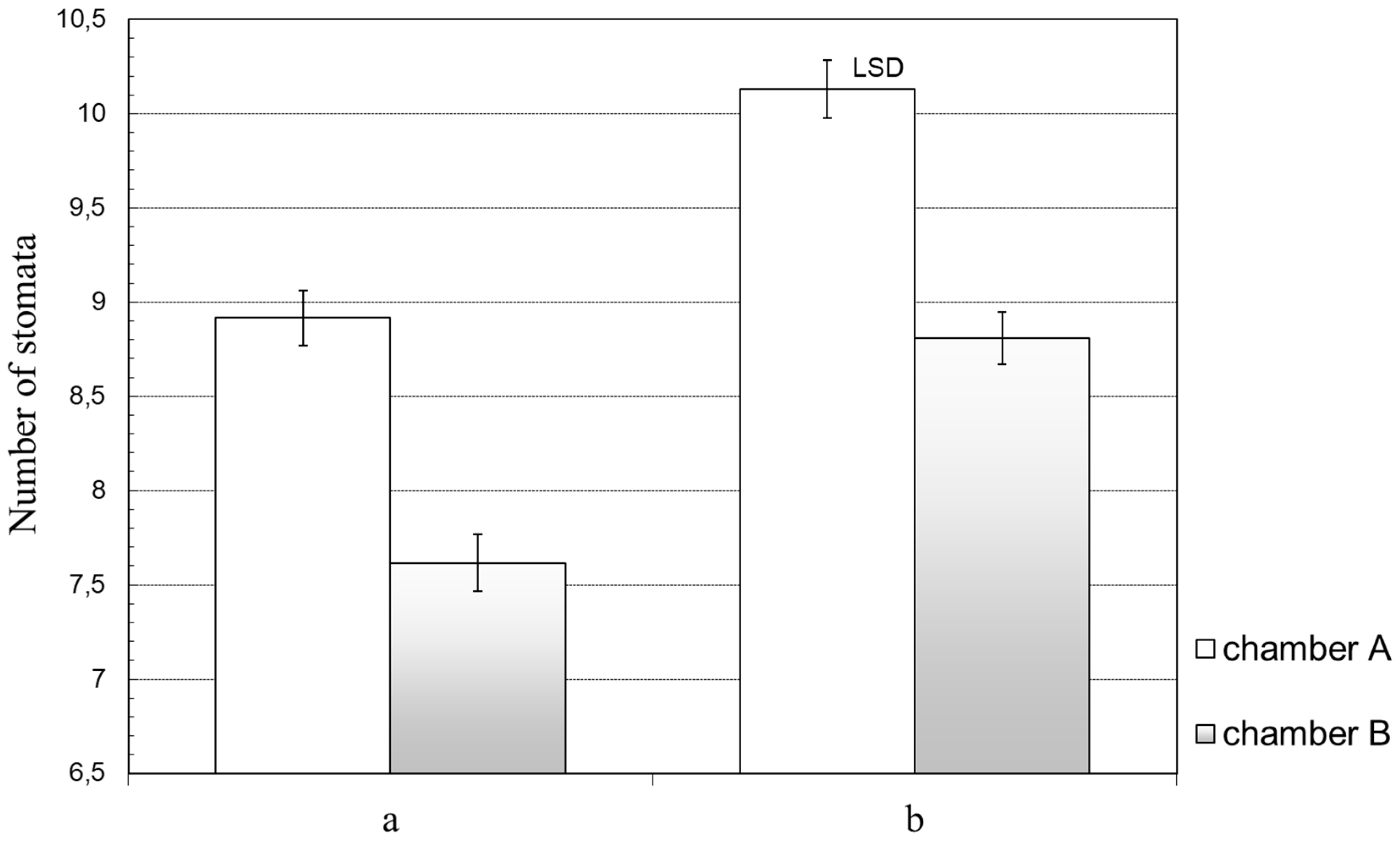
The size of the R/FR ratio may regulate the number of stomata. In previous studies, Doroszewski [
36] found that different spectral compositions caused the number of stomata formed by the wheat to be dependent on the value of the R/FR ratio, as was the case with barley. Plants receiving radiation with a high R/FR ratio produced considerably more stomata on both sides of the leaves than those growing at a low ratio. Close to similar effects were reported by Kasperbauer and Peaslee [
31]. In their experiment with tobacco, the number of stomata on both sides of the leaves was lower for plants radiated with a low R/FR ratio than for plants treated with a high ratio. The results concerning the number of barley and wheat stomata [
36] indicate the plants’ adaptation to the competitive conditions. An increase of the stomata number prevents excessive transpiration. Competitive conditions contribute to an insufficient water supply, so an adaptive response of plants consists not only in closing the stomata but also in limiting their number. In the unfavorable light (chamber B), the number of stomata was decidedly lower because barley behaved as if it grew under typical competitive conditions. The response of barley plants to the spectral composition of the radiation clearly indicated their adaptation to the light conditions.
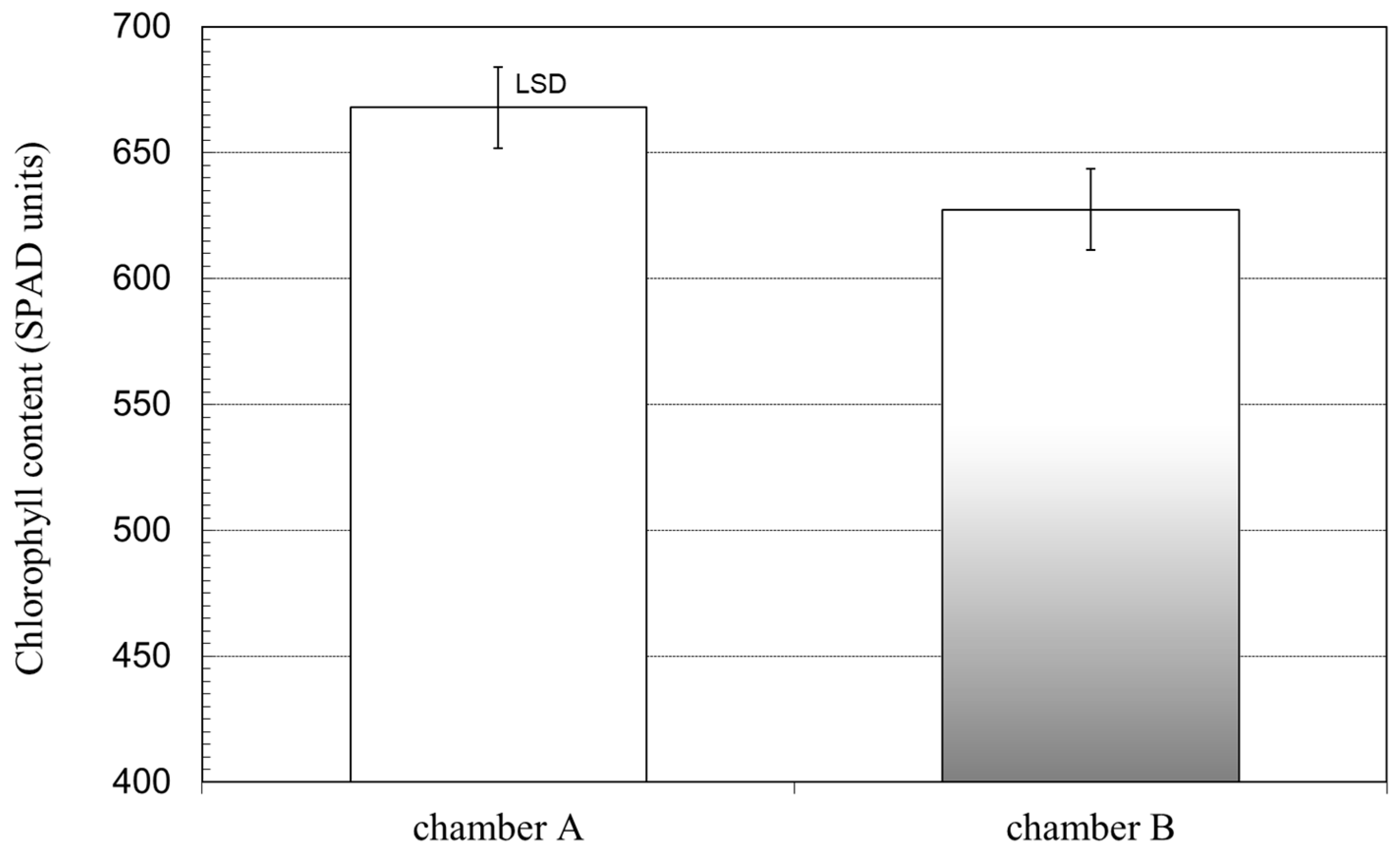
Another element that indicated a differentiated response to the spectral composition of radiation was the chlorophyll content. Both wheat [
36] and, in this study, barley treated with a high R/FR ratio were characterized by a higher chlorophyll concentration than plants submitted to radiation with a low R/FR ratio.
From the spectral-radiometric measurements performed in the outdoor positions [
11], it appears that the R/FR ratio in the solar radiation is close to 1, while in the dense cereal stands it can be lower than 0.07 [
36]. The R/FR ratio has a significant effect on photochromic reactions, which have a considerable effect on the development and yielding of plants. Phytochrome is a photoreceptor responsible for the registration of changes in radiation (solar or artificial). It belongs to the family of photoreceptors, which regulate the plant development in response to environmental radiation. Phytochrome reactions are very important during the whole ontogenesis, starting with seeds’ germination, de-etiolation by the growth of a seedling, the detection of a neighboring plant, the reaction of avoidance of green seedlings’ shading and the induction of reproductive behavior. The basic function of a phytochrome is the perception of R/FR radiation, which, after the signal has been reached (such as these wavelengths), starts the proper metabolic pathways, which allow for the induction of the mechanisms ensuring the optimal growth and development of plants. Smith [
4] put forward a hypothesis about the perception of the R/FR ratio as the fundamental function of a phytochrome. Studies conducted with spring barley fully supported this hypothesis.
The information obtained on the environment in the form of radiation intensity and spectral composition are transmitted by the phytochrome, which regulates many metabolic changes and, respectively, effects on the plant morphology, ensuring that the expected competition is met. The results of the performed measurements under artificial conditions mean that during the whole plant development, physiological processes, e.g., the decrease of stomatal conductance, transpiration and the production of a lower number of stomata, were directed to a final effect—the maximization of adjustment processes in response to the existing conditions of radiation and to survival under conditions of probable stress connected with a lack of water.
The conducted research, which had the character of a model, led to the evaluation of the great importance of plants’ neighborhood and proximity. The need to ensure a certain distance between plants (plant spacing) is particularly important, not only because of the availability of nutrients. It should be underlined that the observed changes on plants’ physiology were not the result of competition but only their earlier preparation for it. This feature may be explained by the existence of a stimulus in the form of radiation with an increased share of FR, which signals the possibility of the occurrence of potential competition.


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}