Phytostabilization of Polluted Military Soil Supported by Bioaugmentation with PGP-Trace Element Tolerant Bacteria Isolated from Helianthus petiolaris

 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Isolation of Endophytic and Rhizospheric Trace Element-Tolerant Bacteria
2.2. Genotypic Characterization of Cd and Pb Tolerant Strains
2.3. In-Vitro PGP Activity of the Strains
2.4. In-Vitro Trace Element Uptake by Bacteria in Liquid Cultures-Scanning Electron Microscopy (SEM-EDX) Analysis
2.5. In-Vitro Inoculation on Vertical Agar Plates (VAPs), Germination and Growth Promotion
2.6. Bioaugmentation in Microcosm Assays, Growth Promotion and Element Bio-Accumulation
2.7. Bacterial Survival and Colonization of Plant Tissues in Microcosms-ARISA
2.8. Statistical Analysis
3. Results
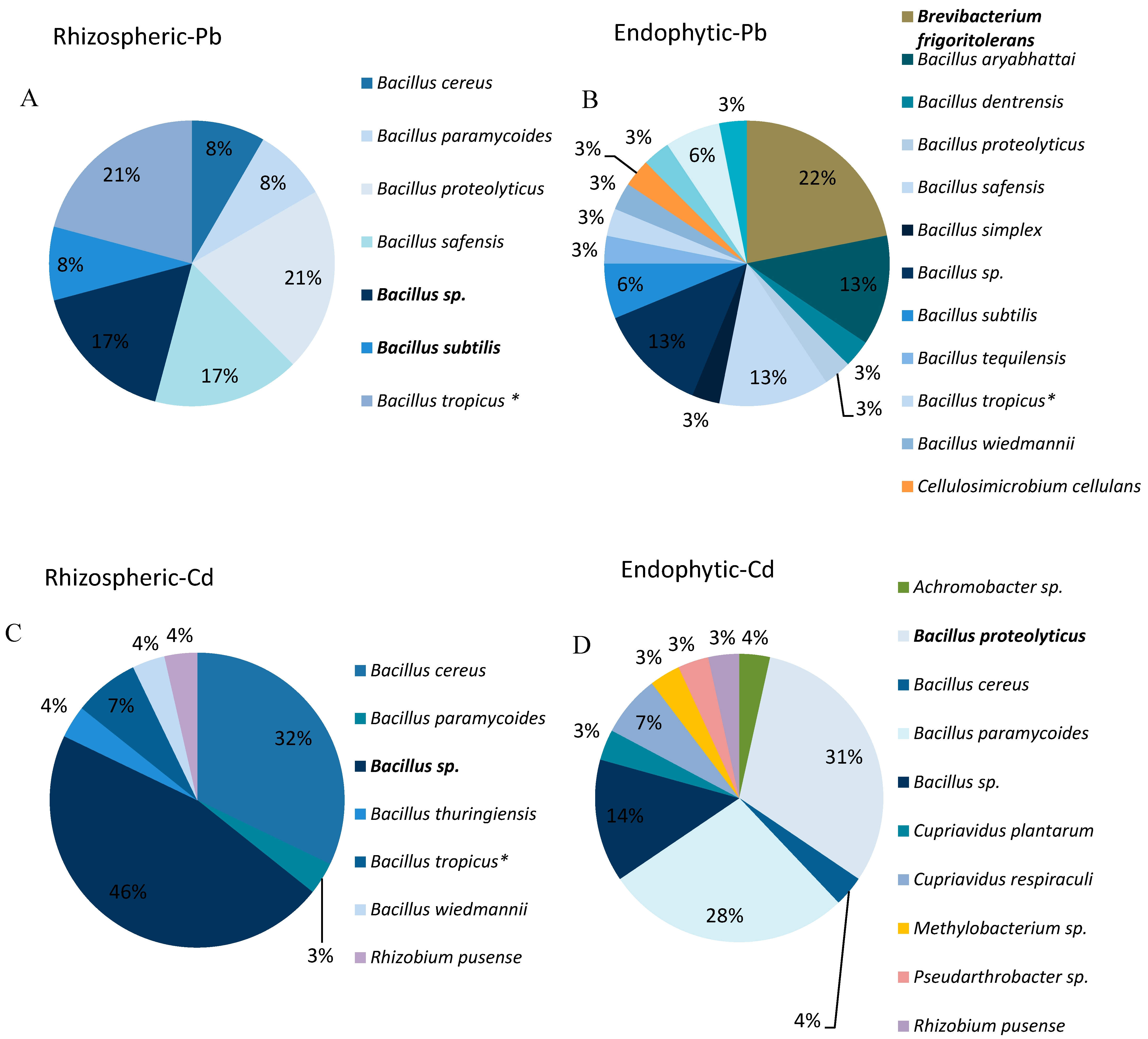
3.1. Isolation of Endophytic and Rhizospheric Trace Element-Tolerant Bacteria
3.2. Genotypic Characterization of Cd and Pb-Tolerant Strains
3.3. In-Vitro PGP Activity of the Strains
3.4. In-Vitro Trace Element Uptake in Culture Media and Scanning Electron Microscopy (SEM-EDX) Analysis
3.5. Germination and Growth Promotion on Vertical Agar Plates (VAPs)
3.6. Growth Promotion and Element Bio-Accumulation in Microcosm Systems
3.7. Bacterial Survival and Colonization of Plant Tissues in Microcosms-ARISA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rizwan, M.; Ali, S.; Rizvi, H.; Rinklebe, J.; Tsang, D.C.; Meers, E.; Ok, Y.S.; Ishaque, W. Phytomanagement of heavy metals in contaminated soils using sunflower: A review. Crit. Rev. Environ. Sci. Technol. 2016, 46, 1498–1528. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metals Toxicity and the Environment; Springer: Basel, Switzerland, 2012; Volume 101, pp. 133–164. [Google Scholar]
- Parab, H.; Joshi, S.; Sudersanan, M.; Shenoy, N.; Lali, A.; Sarma, U. Removal and recovery of cobalt from aqueous solutions by adsorption using low cost lignocellulosic biomass-coir pith. J. Environ. Sci. Health A 2010, 45, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Kalderis, D.; Juhasz, A.L.; Boopathy, R.; Comfort, S. Soils contaminated with explosives: Environmental fate and evaluation of state-of-the-art remediation processes (IUPACtechnical report). Pure Appl. Chem. 2011, 83, 1407–1484. [Google Scholar] [CrossRef]
- Khazaei, M.; Nasseri, S.; Ganjali, M.R.; Khoobi, M.; Nabizadeh, R.; Mahvi, A.H. Response surface modeling of lead, removal by graphene oxide-Fe3O4 nanocomposite using central composite design. J. Environ. Health Sci. Eng. 2016, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Vangronsveld, J.; Herzig, R.; Weyens, N.; Boulet, J.; Adriaensen, K.; Ruttens, A.; Thewys, T.; Vassilev, A.; Meers, E.; Nehnevajova, E.; et al. Phytoremediation of contaminated soils and groundwater: Lessons from the field. Environ. Sci. Pollut. Res. 2009, 16, 765–794. [Google Scholar] [CrossRef]
- Greipsson, S. Phytoremediation. Nat. Educ. Knowl. 2011, 2, 7–12. [Google Scholar]
- Mendez, M.O.; Maier, R.M. Phytostabilization of mine tailings in arid and semiarid environments—An emerging remediation technology. Environ. Health Perspect. 2008, 116, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Lessl, J.T.; Luo, J.; Ma, L.Q. Pteris vittata continuously removed arsenic from non-labile fraction in three contaminated-soils during 3.5 years of phytoextraction. J. Hazard. Mater. 2014, 279, 485–492. [Google Scholar] [CrossRef]
- Adamidis, G.C.; Aloupi, M.; Mastoras, P.; Papadaki, M.I.; Dimitrakopoulos, P.G. Is annual or perennial harvesting more efficient in Ni phytoextraction? Plant Soil 2017, 418, 205–218. [Google Scholar] [CrossRef]
- Płociniczak, T.; Sinkkonen, A.; Romantschuk, M.; Sułowicz, S.; Piotrowska-Seget, Z. Rhizospheric Bacterial Strain Brevibacterium casei MH8a Colonizes Plant Tissues and Enhances Cd, Zn, Cu Phytoextraction by White Mustard. Front. Plant Sci. 2016, 7, 101–122. [Google Scholar] [CrossRef] [PubMed]
- Gadd, G.M. Microbial influence on metal mobility and application for bioremediation. Geoderma 2004, 122, 109–119. [Google Scholar] [CrossRef]
- Jocic, S.; Miladinovic, D.; Kaya, Y. Breeding and Genetics of sunflower. Sunflower: Chemistry, Production, Processing and Utilization; AOCS Press: Urbana, IL, USA, 2015; pp. 187–226. [Google Scholar]
- Alaboudi, K.A.; Ahmeda, B.; Brodiec, G. Phytoremediation of Pb and Cd contaminated soils by using sunflower (Helianthus annuus) plant. Ann. Agric. Sci. 2018, 63, 123–127. [Google Scholar] [CrossRef]
- Govarthanana, M.; Mythilib, R.; Selvankumarb, T.; Kamala-Kannanc, S.; Kima, H. Myco-phytoremediation of arsenic- and lead-contaminated soils by Helianthus annuus and wood rot fungi, Trichoderma sp. isolated from decayed wood. Ecotoxicol. Environ. Saf. 2018, 151, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Ramzani, P.M.A.; Khan, W.D.; Iqbal, M.; Kausar, S.; Ali, S.; Rizwan, M.; Virk, Z.A. Effect of different amendments on rice (Oryza sativa L.) growth, yield, nutrient uptake and grain quality in Ni-contaminated soil. Environ. Sci. Pollut. Res. Int. 2016, 23, 18585–18595. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S.A.; Ashraf, M.; Hussain, M. Phytotoxic effects of nickel on yield and concentration of macro- and micro-nutrients in sunflower (Helianthus annuus L.) achenes. J. Hazard Mater. 2011, 185, 1295–1303. [Google Scholar] [CrossRef]
- Saran, A.; Fernandez, L.; Cora, F.; Savio, M.; Thijs, S.; Vangronsveld, J.; Merini, L.J. Phytostabilization of Pb and Cd polluted soils using Helianthus petiolaris as pioneer aromatic plant species. Int. J. Phytoremediat. 2019, 11, 1–9. [Google Scholar] [CrossRef]
- Shin, S.H.; Lim, Y.; Lee, S.E.; Yang, N.W.; Rhee, J.H. CAS agar diffusion assay for the measurement of siderophores in biological fluids. J. Microbiol. Meth. 2001, 44, 89–95. [Google Scholar] [CrossRef]
- Nguyen, C.; Yan, W.; Le Tacon, F.; Lapayrie, F. Genetic variability of phosphate solubilising activity by monocaryotic and dicaryotic mycelia of the ectomycorrhizal fungus Laccaria bicolor (Maire) P.D. Orton. Plant Soil 1992, 143, 193–199. [Google Scholar] [CrossRef]
- Cunningham, J.E.; Kuiack, C. Production of citric and oxalic acids and solubilization of calcium-phosphate by Penicillium bilaii. Appl. Environ. Microbiol. 1992, 58, 1451–1458. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indole acetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romick, T.L.; Fleming, H.P. Acetoin production as an indicator of growth and metabolic inhibition of Listeria monocytogenes. J. Appl. Microbiol. 1998, 84, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belimov, A.A.; Hontzeas, N.; Safronova, V.I.; Demchinskaya, S.V.; Piluzza, G.; Bullitta, S.; Glick, B.R. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 2005, 37, 241–250. [Google Scholar] [CrossRef]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as a microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.F.; Xia, J.J; Jiang, C.Y.; He, L.Y.; Qian, M. Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ. Pollut. 2008, 156, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.H.H.; Abdelhafez, A.A. Role of EDTA in arsenic mobilization and its uptake by maize grown on an As-polluted soil. Chemosphere 2013, 90, 588–594. [Google Scholar] [CrossRef]
- Cardinale, M.; Brusetti, L.; Quatrini, P.; Borin, S.; Puglia, A.M.; Rizzi, A.; Zanardini, E.; Sorlini, C.; Corselli, C.; Daffonchio, D. Comparison of Different Primer Sets for Use in Automated Ribosomal Intergenic Spacer Analysis of Complex Bacterial Communities. Appl. Environ. Microb. 2004, 70, 6147–6156. [Google Scholar] [CrossRef] [Green Version]
- Michelland, R.J.; Dejean, S.; Combes, S.; Fortun-Lamothe, L.; Cauquil, L. StatFingerprints: A friendly graphical interface program for processing and analysis of microbial fingerprint profiles. Mol. Ecol. Resour. 2009, 9, 1359–1363. [Google Scholar] [CrossRef]
- Truyens, S.; Jambon, I.; Croes, S.; Janssen, J.; Weyens, N.; Mench, M.; Carleer, R.; Cuypers, A.; Vangronsveld, J. The effect of long-term Cd and Ni exposure on seed endophytes of Agrostis capillaris and their potential application in phytoremediation of metal-contaminated soils. Int. J. Phytoremediat. 2014, 16, 643–659. [Google Scholar] [CrossRef]
- Croes, S.; Weyens, N.; Janssen, J.; Vercampt, H.; Colpaert, J.V.; Carleer, R.; Vangronsveld, J. Bacterial communities associated with Brassica napus L. grown on trace element-contaminated and non-contaminated fields: A genotypic and phenotypic comparison. Microb. Biotechnol. 2013, 6, 371–384. [Google Scholar] [CrossRef]
- Montalban, B.; Croes, S.; Weyens, N.; Lobo, M.; Perez-Sanz, A.; Vangronsveld, J. Characterization of bacterial communities associated with Brassica napus L. growing on a Zn-contaminated soil and their effects on root growth. Int. J. Phytoremediat. 2016, 18, 985–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodewyckx, C.; Mergeay, M.; Vangronsveld, J.; Clijsters, H.; van der Lelie, D. Isolation, characterization, and identification of bacteria associated to the zinc hyperaccumulator Thlaspi caerulescens subsp. calaminaria. Int. J. Phytoremediat. 2002, 4, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Mastretta, C.; Taghavi, S.; van der Lelie, D.; Mengoni, A.; Galardi, F.; Gonnelli, C.; Barac, T.; Boulet, J.; Weyens, N.; Vangronsveld, J. Endophytic bacteria from seeds of Nicotiana tabacum can reduce cadmium phytotoxicity. Int. J. Phytoremediat. 2009, 11, 251–267. [Google Scholar] [CrossRef]
- Srikakolapu, M.S. Heavy metal tolerance of Bacillus spp. Soc. Sci. Dev. Agric. Tech. 2014, 8, 315–318. [Google Scholar]
- Idris, R.; Kuffner, M.; Bodrossy, L. Characterization of Ni-tolerant methylobacteria associated with the hyperaccumulating plant Thlaspi goesingense and description of Methylobacterium goesingense sp. nov. Syst. Appl. Microbiol. 2006, 29, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Fomina, M.; Gadd, G.M. Biosorption: Current perspectives on concept, definition and application. Bioresour. Technol. 2014, 160, 3–14. [Google Scholar] [CrossRef]
- Sun, Z.; Lv, Y.; Liu, Y.; Ren, R. Removal of nitrogen by heterotrophic nitrificationaerobic denitrification of a novel metal resistant bacterium Cupriavidus sp. S1. Bioresour. Technol. 2016, 220, 142–150. [Google Scholar] [CrossRef]
- Cardoso, P.; Corticeiro, S.; Freitas, R.; Figueira, E. Different efficiencies of the same mechanisms result in distinct Cd tolerance within Rhizobium. Ecotoxicol. Environ. Saf. 2018, 150, 260–269. [Google Scholar] [CrossRef]
- Remans, T.; Nacry, P.; Pervent, M.; Girin, T.; Tillard, P.; Lepetit, M.; Gojon, A. A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation in Arabidopsis. Plant Physiol. 2006, 140, 909–921. [Google Scholar] [CrossRef] [Green Version]
- Fisher, P.J.; Petrini, O.; Scott, H.M.L. The distribution of some fungal and bacterial endophytes in maize. New Phytol. 1992, 122, 299–305. [Google Scholar] [CrossRef]
- Benizri, E.; Baudoin, E.; Guckert, A. Root colonization by inoculated plant growth rhizobacteria. Biocontrol Sci. Technol. 2001, 11, 557–574. [Google Scholar] [CrossRef]
- Mehta, S.; Nautiyal, C.S. An Efficient Method for Qualitative Screening of Phosphate-Solubilizing Bacteria. Curr. Microbiol. 2001, 43, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.N. Environmental applications for biosurfactants. Environ. Pollut. 2005, 133, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Montalban, B.; Thijs, S.; Lobo, MC.; Weyens, N.; Ameloot, M.; Vangronsveld, J.; Perez-Sanz, A. Cultivar and Metal-Specific Effects of Endophytic Bacteria in Helianthus tuberosus Exposed to Cd and Zn. Int. J. Mol. Sci. 2017, 18, 2026. [Google Scholar] [CrossRef]
- Das, N.; Basak, L.V.G.; Salam, J.A.; Abigail, E.A. Application of biofilms on remediationmof pollutants, an overview. J. Microbiol. Biotechnol. 2012, 2, 783–790. [Google Scholar]
- Sessitsch, A.; Kuffner, M.; Kidd, P.; Vangronsveld, J.; Wenzel, W.; Fallmann, K.; Puschenreiter, M. The role of plant-associated bacteria in the mobilization and phytoextraction of trace elements in contaminated soils. Soil Biol. Biochem. 2013, 60, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Alkan, H.; Gul-Guven, R.; Guven, K.; Erdogan, S.; Dogru, M. Biosorption of Cd, Cu, and Ni Ions by a Thermophilic Haloalkalitolerant Bacterial Strain (KG9) Immobilized on Amberlite XAD-4. Pol. J. Environ. Stud. 2015, 24, 1903–1910. [Google Scholar] [CrossRef]
- Oh, S.E.; Hassan, S.H.; Joo, J.H. Biosorption of heavy metals by lyophilized cells of Pseudomonas stutzeri. World J. Microb. Biot. 2009, 25, 1771–1778. [Google Scholar] [CrossRef]
- Prapagdee, B.; Chanprasert, M.; Mongkolsuk, S. Bioaugmentation with cadmium-resistant plant growth-promoting rhizobacteria to assist cadmium phytoextraction by Helianthus annuus. Chemosphere 2013, 92, 659–666. [Google Scholar] [CrossRef]
- Zhuang, L.; Zhang, J.; Xiang, X. Point mutations in the stem region and the fourth AAA domain of cytoplasmic dynein heavy chain partially suppress the phenotype of NUDF/LIS1 loss in Aspergillus nidulans. Genetic 2007, 175, 1185–1196. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Persello Cartieaux, F.; Nussaume, L.; Robaglia, C. Tales from the underground: Molecular plant-rhizobacteria interactions. Plant Cell Environ. 2003, 26, 189–199. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Wu, J.; Zhang, J. The effect of biotic and abiotic environmental factors on Pd(II) adsorption and reduction by Bacilius wiedmannii MSM. Ecotoxicol. Environ. Saf. 2018, 162, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.-Y.; Cho, K.-S. Characterization of a Heavy Metal-Resistant and Plant Growth-Promoting Rhizobacterium, Methylobacterium sp. SY-NiR1. Korean J. Microbiol. Biot. 2007, 35, 58–65. [Google Scholar]
- Madhaiyan, M.S.; Poonguzhali, S.; Sa, T. Metal tolerating methylotrophic bacteria reduces nickel and cadmium toxicity and promotes plant growth of tomato (Lycopersicon esculentum L.). Chemosphere 2007, 69, 220–228. [Google Scholar] [CrossRef]
- Karthik, C.; Barathi, S.; Pugazhendhi, A.; Ramkumar, S.R.; Dung Thi, N.B.; Arulselvi, P.I. Evaluation of Cr(VI) reduction mechanism and removal by Cellulosimicrobium funkei strain AR8, a novel haloalkaliphilic bacterium. J. Hazard. Mater. 2017, 333, 42–53. [Google Scholar] [CrossRef]
- Płociniczak, T.; Chod, M.; Pacwa-Płociniczak, M.; Piotrowska-Seget, Z. Metal-tolerant endophytic bacteria associated with Silene vulgaris support the Cd and Zn phytoextraction in non-host plants. Chemosphere 2016, 219, 250–260. [Google Scholar] [CrossRef]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Piotrowska-Seget, Z. Monitoring the changes in a bacterial community in petroleum-polluted soil bioaugmented with hydrocarbon-degrading strains. Appl. Soil Ecol. 2016, 105, 76–85. [Google Scholar] [CrossRef]
- Ghasemi, Z.; Ghaderian, S.M.; Rodríguez-Garrido, B.; Prieto-Fernández, A.; Kidd, P.S. Plant species-specificity and effects of bioinoculants and fertilization on plant performance for nickel phytomining. Plant Soil 2018, 425, 265–285. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical properties | |||
| Texture (%) | Conductivity (µS cm−1) | CEC (Meq 100 g−1 DW) | pH |
| sandy-loam | 340 ± 1.2 | 9.2 ± 0.2 | 6.2 ± 0.8 |
| Trace element concentration (mg kg−1) | |||
| Cd | Cu | Pb | Zn |
| 0.42 ± 1.35 | 1.02 ± 1.16 | 5.48 ± 0.89 | 12.43 ± 20.07 |
| Values are mean ± S.E. (n = 24). | |||
| Strain | ACCD | Acetoin | a OA | b P-Solub. | b SID | c IAA | d Biofilm | pH | e Element Conc. |
|---|---|---|---|---|---|---|---|---|---|
| Lead tolerant | |||||||||
| Bacillus sp. ST1 | + | + | + | 0.06 ± 0.01 * | 0.16 ± 0.01 ** | 0.00 ± 0.00 | 0.01 ± 0.00 | 4.27 ± 0.18 * | 5.91 ± 0.05 ** |
| Bacillus proteoliticus ST4 | + | + | + | 0.06 ± 0.01 * | 0.26 ± 0.02 ** | 43.61 ± 1.03 * | 0.03 ± 0.00 * | 4.23 ± 0.28 * | 5.67 ± 0.05 * |
| Bacillus subtilis ST7 | - | + | - | 0.23 ± 0.01 ** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.08 ± 0.02 * | 4.22 ± 0.08 * | 6.31 ± 0.09 ** |
| Bacillus paramycoides ST9 | + | + | + | 0.19 ± 0.01 ** | 0.16 ± 0.01 | 0.00 ± 0.00 | 0.02 ± 0.00 | 4.20 ± 0.15 * | 5.85 ± 0.07 ** |
| Bacillus cereus ST10 | + | + | + | 0.09 ± 0.01 * | 0.18 ± 0.01 ** | 0.00 ± 0.00 | 0.02 ± 0.00 | 4.16 ± 0.07 * | 6.19 ± 0.05 ** |
| Bacillus safensis ST11 | - | + | + | 0.08 ± 0.01 * | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 4.15 ± 0.07 * | 6.02 ± 0.04 ** |
| Bacillus tropicus ST22 | + | + | + | 0.06 ± 0.01 * | 0.20 ± 0.01 ** | 0.00 ± 0.00 | 0.01 ± 0.00 | 4.21 ± 0.18 * | 6.01 ± 0.07 ** |
| Bacillus aryabhattai ST25 | + | - | + | 0.34 ± 0.02 ** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.03 ± 0.00 | 4.21 ± 0.32 * | 6.03 ± 0.05 ** |
| Brevibacterium frigoritolerans ST30 | - | + | + | 0.37 ± 0.02 ** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.02 ± 0.00 | 4.26 ± 0.18 * | 5.20 ± 0.12 * |
| Bacillus wiedmannii ST29 | + | + | + | 0.05 ± 0.01 * | 0.53 ± 0.04 ** | 0.00 ± 0.00 | 0.01 ± 0.00 | 4.25 ± 0.04 * | 5.09 ± 0.08 * |
| Bacillus tequilensis ST34 | - | + | - | 0.04 ± 0.00 * | 0.14 ± 0.01 ** | 0.00 ± 0.00 | 0.03 ± 0.00 * | 4.254 ± 0.08 * | 5.30 ± 0.10 * |
| Bacillus dentrensis ST38 | - | + | - | 0.05 ± 0.01 * | 0.11 ± 0.01 * | 0.00 ± 0.00 | 0.02 ± 0.00 | 4.17 ± 0.07 * | 6.58 ± 0.22 ** |
| Bacillus simplex ST43 | + | - | + | 0.01 ± 0.00 | 0.00 ± 0.00 | 54.30 ± 2.08 * | 0.02 ± 0.00 | 4.24 ± 0.08* | 5.79 ± 0.53* |
| Gordonia terrae ST51 | + | - | + | 0.08 ± 0.01 * | 0.22 ± 0.02 ** | 0.00 ± 0.00 | 0.04 ± 0.00 * | 4.18 ± 0.07 * | 8.22 ± 0.432 ** |
| Cellulosimicrobium cellulans-ST54 | + | + | + | 0.10 ± 0.01 * | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.07 ± 0.02 * | 4.21 ± 0.27 * | 6.22 ± 0.12 ** |
| Negative control (GYPb) | - | - | - | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.00 | 6.08 ± 0.02 | 0.49 ± 0.08 |
| Cadmium tolerant | |||||||||
| Bacillus cereus ST60 | + | + | + | 0.01 ± 0.00 | 0.10 ± 0.01 * | 0.00 ± 0.00 | 0.29 ± 0.06 ** | 5.08 ± 0.35 * | 7.67 ± 0.33 * |
| Bacillus tropicus ST77 | + | + | + | 0.00 ± 0.00 | 0.38 ± 0.03 ** | 50.08 ± 2.00 ** | 0.03 ± 0.00 * | 4.97 ± 0.027 * | 7.89 ± 0.09 |
| Rhizobium pusense ST80 | - | - | + | 0.15 ± 0.01 ** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.05 ± 0.00 * | 6.83 ± 0.44 | 3.48 ± 0.05 ** |
| Methylobacterium ST85 | + | + | + | 0.20 ± 0.01 ** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.00 | 6.98 ± 0.42 | 5.00 ± 0.12 ** |
| Pseudobacter sp. ST86 | + | + | + | 0.26 ± 0.02 ** | 0.00 ± 0.00 | 30.25 ± 1.98 * | 0.03 ± 0.00 * | 6.05 ± 0.26 | 4.08 ± 0.13 ** |
| Bacillus sp. ST87 | - | + | + | 0.08 ± 0.01 * | 0.10 ± 0.01 * | 0.00 ± 0.00 | 0.04 ± 0.00 * | 4.70 ± 0.15 * | 7.98 ± 0.21 |
| Bacillus proteolyticus ST89 | + | + | + | 0.03 ± 0.00 | 0.38 ± 0.03 ** | 0.00 ± 0.00 | 0.13 ± 0.02 * | 4.99 ± 0.28 * | 7.74 ± 0.21 * |
| Bacillus cereus ST90 | + | + | + | 0.12 ± 0.01 * | 0.60 ± 0.04 ** | 0.00 ± 0.00 | 0.04 ± 0.00 * | 4.89 ± 0.019 * | 6.29 ± 0.14 * |
| Achomobacter sp. ST95 | - | + | + | 0.10 ± 0.01 * | 0.24 ± 0.02 ** | 0.00 ± 0.00 | 0.10 ± 0.02 * | 6.98 ± 0.38 | 4.87 ± 0.28 ** |
| Bacillus paramycoides ST98 | + | + | + | 0.02 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.05 ± 0.00 * | 4.97 ± 0.15 * | 7.87 ± 0.14 * |
| Cuprvidus plantarum ST102 | + | + | + | 0.09 ± 0.01 * | 0.31 ± 0.02 ** | 25.94 ± 1.02 * | 0.02 ± 0.00 | 4.86 ± 0.13 * | 7.87 ± 0.9 * |
| Klebsiella variicola ST106 | + | + | + | 0.36 ± 0.02 ** | 0.00 ± 0.00 | 81.47 ± 4.55 ** | 0.07 ± 0.02 * | 3.57 ± 0.17 ** | 7.95 ± 0.12 |
| Negative control (GYCd) | - | - | - | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 6.6 ± 0.03 | 8.27 ± 0.12 |
| Strain | Stem Length (cm) | Dry Weight Stem (g) | Pb (mg Kg−1) | Cd (mg Kg−1) | BAF | |||
|---|---|---|---|---|---|---|---|---|
| Root | Leaves | Root | Leaves | Pb | Cd | |||
| Bacillus paramycoides ST4 | 4.7 | 2.00 ± 1.03 | 21.40 ± 1.61 | 9.06 ± 1.60 | 0.31 ± 0.23 | 0.02 ± 0.04 | 3.14 c | 25.63 d |
| Bacillus wiedmanni ST29 | 5.7 | 1.14 ± 0.35 | 40.14 ± 5.88 | 12.97 ± 4.96 | <DL a | 0.24 ± 0.19 | 1.09 ab | 2.46 |
| Brevibacterium frigoritolerans ST30 | 7.3 | 2.21 ± 1.40 | 55.90 ± 18.93 | 4.95 ± 3.22 | 0.02 ± 0.31 | 0.22 ± 0.05 | 1.66 b | 4.83 b |
| Cellulosimicrobium cellulans ST54 | 6.3 | 1.86 ± 0.93 | 35.77 ± 6.74 | 3.62 ± 1.25 | 0.33 ± 0.48 | 0.17 ± 0.12 | 6.17 d | 61.28 e |
| Methylobacterium sp. ST85 | 5.3 | 1.66 ± 1.11 | 20.02 ± 1.51 | 3.51 ± 2.77 | 0.51 ± 0.17 | 0.14 ± 0.02 | 1.84 b | 18.69 c |
| Bacillus proteolyticus ST9 | 11.0 | 2.50 ± 0.51 | 15.60 ± 1.54 | 3.40 ± 1.03 | 0.37 ± 0.26 | 0.21 ± 0.08 | 0.73 a | 2.45 a |
| Control | 7.0 | 1.85 ± 0.43 | 28.89 ± 2.04 | 3.87 ± 1.25 | 0.43 ± 0.25 | 0.08 ± 0.01 | 0.92 a | 4.15 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saran, A.; Imperato, V.; Fernandez, L.; Gkorezis, P.; d’Haen, J.; Merini, L.J.; Vangronsveld, J.; Thijs, S. Phytostabilization of Polluted Military Soil Supported by Bioaugmentation with PGP-Trace Element Tolerant Bacteria Isolated from Helianthus petiolaris. Agronomy 2020, 10, 204. https://doi.org/10.3390/agronomy10020204
Saran A, Imperato V, Fernandez L, Gkorezis P, d’Haen J, Merini LJ, Vangronsveld J, Thijs S. Phytostabilization of Polluted Military Soil Supported by Bioaugmentation with PGP-Trace Element Tolerant Bacteria Isolated from Helianthus petiolaris. Agronomy. 2020; 10(2):204. https://doi.org/10.3390/agronomy10020204
Chicago/Turabian StyleSaran, Anabel, Valeria Imperato, Lucia Fernandez, Panos Gkorezis, Jan d’Haen, Luciano Jose Merini, Jaco Vangronsveld, and Sofie Thijs. 2020. "Phytostabilization of Polluted Military Soil Supported by Bioaugmentation with PGP-Trace Element Tolerant Bacteria Isolated from Helianthus petiolaris" Agronomy 10, no. 2: 204. https://doi.org/10.3390/agronomy10020204
APA StyleSaran, A., Imperato, V., Fernandez, L., Gkorezis, P., d’Haen, J., Merini, L. J., Vangronsveld, J., & Thijs, S. (2020). Phytostabilization of Polluted Military Soil Supported by Bioaugmentation with PGP-Trace Element Tolerant Bacteria Isolated from Helianthus petiolaris. Agronomy, 10(2), 204. https://doi.org/10.3390/agronomy10020204