Leaf Removal Affects Maize Morphology and Grain Yield


 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
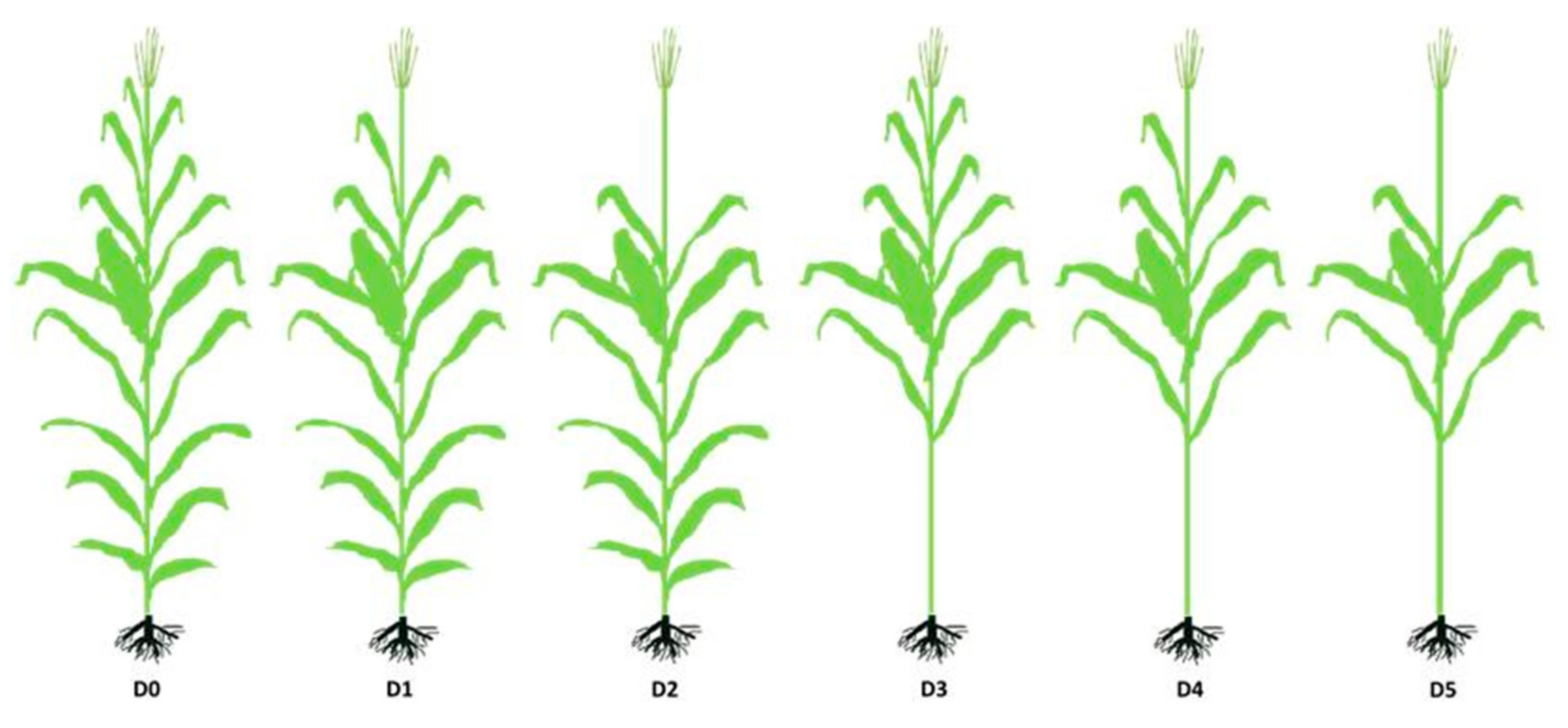
2.1. Site and Experiment Design
2.2. Sampling and Measurement
2.3. Statistical Analysis
3. Results
3.1. Grain Yield and Yield Components
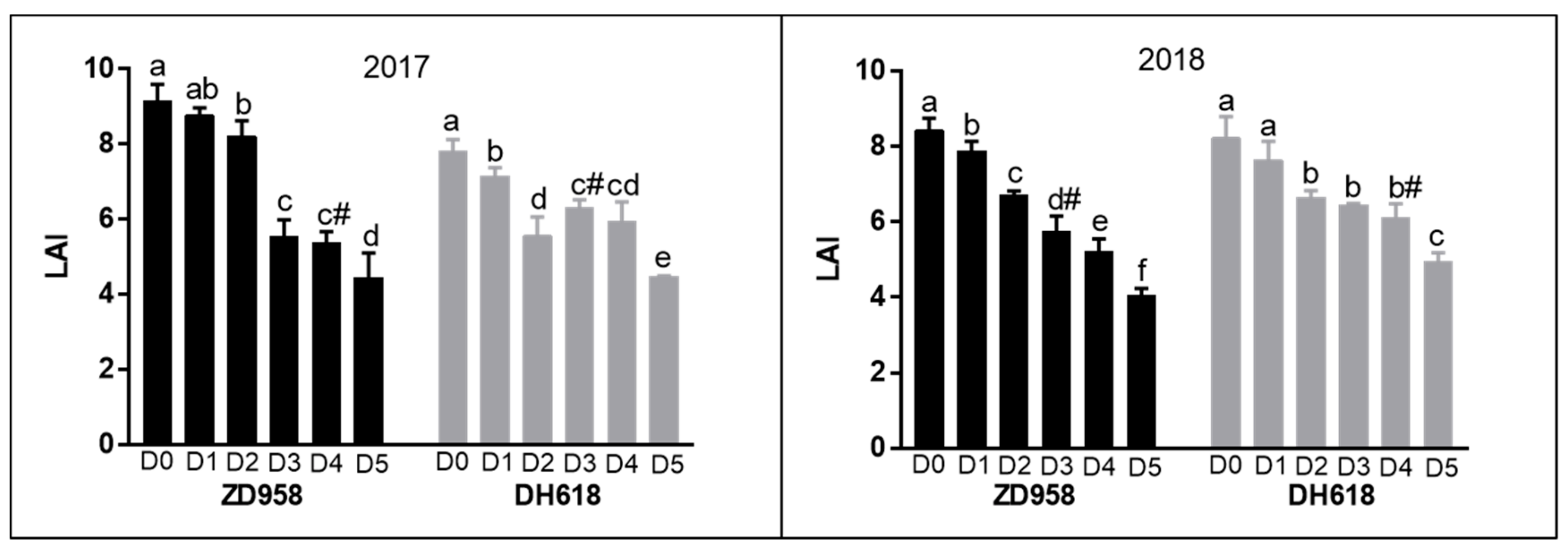
3.2. Maximum Leaf Area Index (LAI)
3.3. Dry Matter Accumulation and Post-Silking Source–Sink Ratio
3.4. Light Distribution
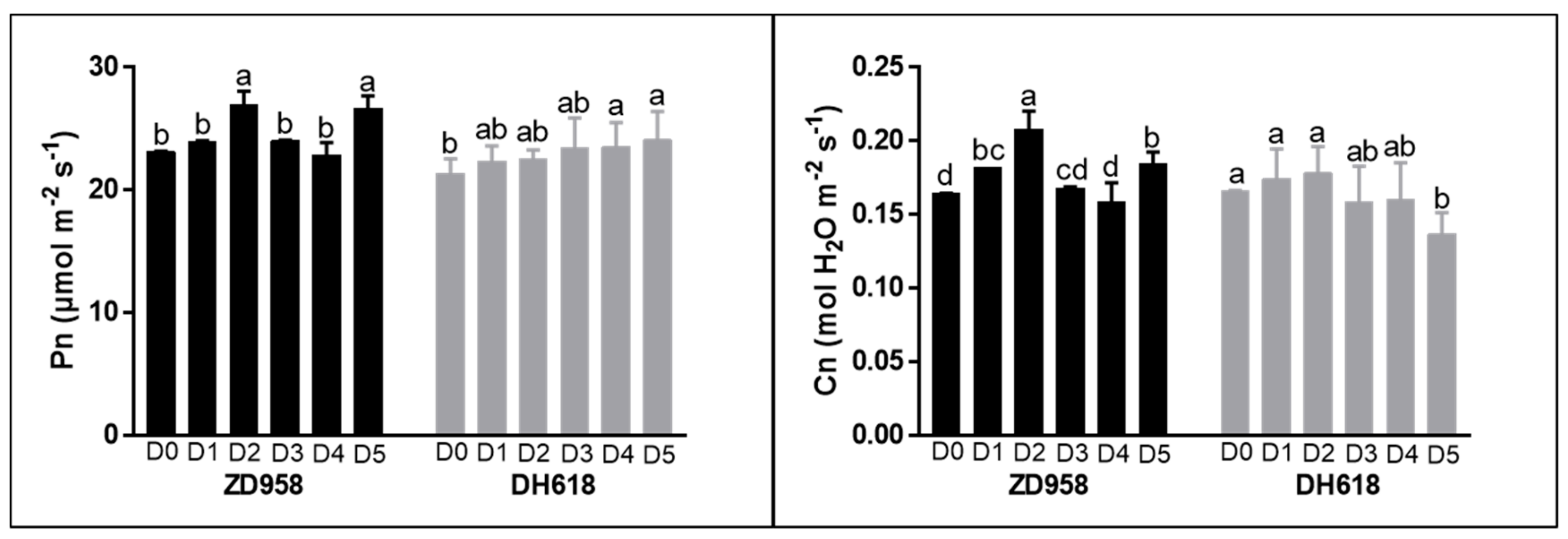
3.5. Photosynthetic Rate and Stomatal Conductance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Grassini, P.; Thorburn, J.; Burr, C.; Cassman, K.G. High-yield irrigated maize in the western US Corn Belt I. On-farm yield, yield potential, and impact of agronomic practices. Field Crop. Res. 2011, 120, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, V.P.; Zhou, Z. Food demand and the food security challenge with rapid economic growth in the emerging economies of India and China. Food Res. Int. 2014, 63, 108–124. [Google Scholar] [CrossRef]
- Long, S.P.; Zhu, X.G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef]
- Meng, Q.F.; Hou, P.; Wu, L.; Chen, X.P.; Cui, Z.L.; Zhang, F.S. Understanding production potentials and yield gaps in intensive maize production in China. Field Crop. Res. 2013, 143, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Tokatlidis, I.S.; Koutroubas, S.D. A review of maize hybrids’ dependence on high plant populations and its implications for crop yield stability. Field Crop. Res. 2004, 88, 103–114. [Google Scholar] [CrossRef]
- Liu, G.Z.; Hou, P.; Xie, R.Z.; Ming, B.; Wang, K.R.; Xu, W.J.; Liu, W.M.; Yang, Y.S.; Li, S.K. Canopy characteristics of high-yield maize with yield potential of 22.5 Mg ha−1. Field Crop. Res. 2017, 213, 221–230. [Google Scholar] [CrossRef]
- Xu, W.J.; Liu, C.W.; Wang, K.R.; Xie, R.Z.; Ming, B.; Wang, Y.H.; Zhang, G.Q.; Liu, G.Z.; Zhao, R.L.; Fan, P.P.; et al. Adjusting maize plant density to different climatic conditions across a large longitudinal distance in China. Field Crop. Res. 2017, 212, 126–134. [Google Scholar] [CrossRef]
- Xue, J.; Gou, L.; Shi, Z.G.; Zhao, Y.S.; Zhang, W.F. Effect of leaf removal on photosynthetically active radiation distribution in maize canopy and stalk strength. J. Integr. Agr. 2017, 16, 85–96. [Google Scholar] [CrossRef]
- Christensen, L.E.; Below, F.E.; Hageman, R.H. The effect of ear removal on senescence and metabolism of maize. Plant physiol. 1981, 68, 1180–1185. [Google Scholar] [CrossRef] [Green Version]
- Maddonni, G.A.; Otegui, M.E. Intra-specifc competition in maize: Early establishment of hierarchies among plants affects final kernel set. Field Crop. Res. 2004, 85, 1–13. [Google Scholar] [CrossRef]
- Huang, S.B.; Gao, Y.B.; Li, Y.B.; Tao, H.B.; Wang, P. Influence of plant architecture on maize physiology and yield in the Heilonggang River valley. Crop J. 2017, 5, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Li, R.F.; Liu, P.; Yang, Q.L.; Ren, H.; Dong, S.T.; Zhang, J.W.; Zhao, B. Effects of lower leaf senescence on carbon and nitrogen distribution and yield formation in maize (Zea mays L.) with high planting density. Acta Agron. Sin. 2018, 44, 1032–1042. [Google Scholar] [CrossRef]
- Boomsma, C.R.; Santini, J.B.; Tollenaar, M.; Vyn, T.J. Maize morphophysiological responses to intense crowding and low nitrogen availability: an analysis and review. Agron. J. 2009, 101, 1426–1452. [Google Scholar] [CrossRef] [Green Version]
- Timlin, D.J.; Fleisher, D.H. Plant density and leaf area index effects on the distribution of light transmittance to the soil surface in maize. Agron. J. 2015, 106, 1828–1837. [Google Scholar] [CrossRef]
- Liu, T.N.; Gu, L.M.; Dong, S.T.; Zhang, J.W.; Liu, P.; Zhao, B. Optimum leaf removal increases canopy apparent photosynthesis, 13C-photosynthate distribution and grain yield of maize crops grown at high density. Field Crop. Res. 2015, 170, 32–39. [Google Scholar] [CrossRef]
- Srinivasan, V.; Kumar, P.; Long, S.P. Decreasing, not increasing, leaf area will raise crop yields under global atmospheric change. Global Change Biol. 2017, 23, 1626–1635. [Google Scholar] [CrossRef] [Green Version]
- Li, D.H. Review and prospect in compact plant type breeding in maize. Crops 2000, 1–5. [Google Scholar]
- Kiniry, J.R.; Bean, B.; Xie, Y.; Chen, P.Y. Maize yield potential: critical processes and simulation modeling in a high-yielding environment. Agr. Syst. 2004, 82, 45–56. [Google Scholar] [CrossRef]
- Wang, X.P.; Li, B.G.; Guo, Y.; Zhai, Z.X. Measurement and analysis of the 3D spatial distribution of photosynthetically active radiation in maize canopy. Acta Agron. Sin. 2004, 30, 568–576. [Google Scholar]
- Ma, Y.H.; Xue, J.Q.; Zhang, R.H.; Zhang, L.C.; Hao, Y.; Sun, J. Relationship between dry matter accumulation and distribution to yield of different maize cultivars. Guangdong Agr. Sci. 2010, 36–40. [Google Scholar]
- Roekel, V.R.J.; Couter, J.A. Agronomic responses of corn to planting date and plant density. Agron. J. 2011, 103, 1414–1422. [Google Scholar] [CrossRef]
- Yang, D.G.; Zhao, W.; Qin, D.L.; Liu, F.Y.; Zhang, Q.; Guan, Y.; Yang, K.J. Yield and canopy structure of maize under the condition of high yield cultivation. J. Maize Sci. 2016, 24, 129–135. [Google Scholar]
- Sheng, C.F. Growth tedionsness as an explanation of over compensation of crops for insect feeding. J. Appl. Eco. 1990, 1, 26–30. [Google Scholar]
- Zhu, G.X.; Midmore, D.J.; Radford, B.J.; Yule, D.F. Effect of timing of defoliation on wheat (Triticum aestivum) central Queensland. 1. Crop response and yield. Field Crop. Res. 2004, 88, 211–226. [Google Scholar] [CrossRef]
- Chen, Y.Z.; Kong, X.Q.; Dong, H.Z. Removal of early fruiting branches impacts leaf senescence and yield by altering the sink/source ratio of field-grown cotton. Field Crop. Res. 2018, 216, 10–21. [Google Scholar] [CrossRef]
- Tollenaar, M.; Daynard, T.B. Effect of defoliation on kernel development in maize. Can. J. Plant Sci. 1978, 58, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.N.; Huang, R.D.; Cai, T.; Han, Q.F.; Dong, S.T. Optimum leaf removal increases nitrogen accumulation in kernels of maize grown at high density. Sci. Rep. 2017, 7, 39601. [Google Scholar] [CrossRef]
- Tollenaar, M. Sink-source relationships during reproductive development in maize. A review. Maydica 1977, 22, 49–75. [Google Scholar]
- Uhart, S.A.; Andrade, F.H. Nitrogen and carbon accumulation and remobilization during grain filling in maize under different source/sink ratios. Crop Sci. 1995, 35, 183–190. [Google Scholar] [CrossRef]
- Hao, M.B.; Wang, K.J.; Dong, S.T.; Zhang, J.W.; Li, D.H.; Liu, P.; Yang, J.S.; Liu, J.G. Leaf redundancy of high-yielding maize (Zea may L.) and its effects on maize yield and photosynthesis. Chinese J. Appl. Ecol. 2010, 21, 344–350. [Google Scholar]
- Liu, Z.Q.; Xu, X.M.; Shen, H.B.; Sun, J.H.; Shen, F.Y.; Che, S.P.; Wang, S.Y.; Lu, L.P. Effects of leaf cutting treatment to maize yield under the high plant population cultivation condition. J. Maize Sci. 2009, 17, 74–75. [Google Scholar] [CrossRef]
- Maddonni, G.A.; Otegui, M.E. Leaf area, light interception, and crop development in maize. Field Crop. Res. 1996, 48, 81–87. [Google Scholar] [CrossRef]
- Stewart, D.; Costa, C.; Dwyer, L.; Smith, D.; Hamilton, R.; Ma, B. Canopy structure, light interception, and photosynthesis in maize. Agron. J. 2003, 95, 1465–1474. [Google Scholar] [CrossRef]
- Subedi, K.D.; Ma, B.L. Ear position, leaf area and contribution of individual leaves to grain yield in conventional and leafy maize hybrids. Crop Sci. 2005, 45, 2246–2257. [Google Scholar] [CrossRef]
- Wei, S.S.; Wang, X.Y.; Jiang, D.; Dong, S.T. Physiological and proteome studies of maize (Zea mays L.) in response to leaf removal under high plant density. BMC Plant Biol. 2018, 18, 378. [Google Scholar] [CrossRef]
- Liu, G.Z.; Hou, P.; Xie, R.Z.; Ming, B.; Wang, K.R.; Liu, W.M.; Yang, Y.S.; Xu, W.J.; Chen, J.L.; Li, S.K. Nitrogen uptake and response to radiation distribution in the canopy of high-yield maize. Crop Sci. 2019, 59, 1236–1247. [Google Scholar] [CrossRef]
- Mu, X.H.; Chen, Q.W.; Chen, F.J.; Yuan, L.X.; Mi, G.H. Within-leaf nitrogen allocation in allocation in adaptation to low nitrogen supply in maize during grain-filling stage. Front. Plant Sci. 2016, 7, 699. [Google Scholar] [CrossRef]
- Ma, D.L.; Xie, R.Z.; Niu, X.K.; Li, S.K.; Long, H.L.; Liu, Y.E. Changes in the morphological traits of maize genotypes in China between the 1950 and 2000. Eur. J. Agron. 2014, 58, 1–10. [Google Scholar] [CrossRef]
- Hemingway, J.S. Effects of population density on yield of maize. East Afric. Agri. J. 1957, 22, 199–202. [Google Scholar] [CrossRef]
- Milander, L.; Jukic, Z.; Mason, S.; Glausha, T.; Kmail, Z. Plant population influence on maize yield components in Croatia and Nebraska. Crop Sci. 2016, 56, 2742–2750. [Google Scholar] [CrossRef]
- Zhou, B.Y.; Yue, Y.; Sun, X.F.; Wang, X.B.; Wang, Z.M.; Ma, W.; Zhao, M. Maize grain yield and dry matter production responses to variation in weather conditions. Agron. J. 2016, 108, 196–204. [Google Scholar] [CrossRef]
- Rivera-Amado, C.; Trujillo-Negrellos, E.; Molero, G.; Reynolds, M.P.; Sylvester-Bradley, R.; Foulkes, M.J. Optimizing dry-matter partitioning for increased spike growth, grain number and harvest index in spring wheat. Field Crop. Res. 2019, 240, 154–167. [Google Scholar] [CrossRef]
- Zhao, K.F. Effect of the leaves of different positions in maize on the corn yield and the photosynthetic properties of those leaves after the growing out of the female flowers. Acta Agron. Sin. 1981, 7, 259–266. [Google Scholar]
- Stirling, C.M.; Aguilera, C.; Baker, N.R.; Long, S.P. Changes in the photosynthetic light response curve during leaf development of field grown maize with implications for modelling canopy photosynthesis. Photosynth. Res. 1994, 42, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Austin, R.B.; Ford, M.A.; Edrich, J.A.; Hooper, B.E. Some effects of leaf posture on photosynthesis and yield in wheat. Ann. Appl. Biol. 1976, 83, 425–446. [Google Scholar] [CrossRef]
- Lambert, R.J.; Mansfield, B.D.; Mumm, R.H. Effect of leaf area on maize productivity. Maydica 2014, 59, 58–64. [Google Scholar]
- Liu, T.N.; Xu, C.L.; Gu, L.M.; Dong, S.T. Effects of leaf removal on canopy apparent photosynthesis and individual leaf photosynthetic characteristics in summer maize under high plant density. Acta Agron. Sin. 2014, 40, 143–153. [Google Scholar] [CrossRef]
- Wang+, Z.; Wu, Z.H.; Xu, K.Z. Relationship between leaf area index and photosynthetic rate of maize canopies. J. Jilin Agri. Univer. 2001, 23, 9–12. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Year | Cultivar | Treatment | Yield (Mg ha−1) | Ear Number (104 ears ha−1) | KNP | TKW (g) |
|---|---|---|---|---|---|---|
| 2017 | ZD958 | D0 | 19.4 ± 0.7b | 11.3 ± 0.0a | 469 ± 15ab | 382.5 ± 16.9a |
| D1 | 20.0 ± 1.4ab | 11.7 ± 1.2a | 530 ± 80a | 358.6 ± 13.4a | ||
| D2 | 18.3 ± 0.4b | 12.1 ± 0.5a | 450 ± 5b | 369.3 ± 6.0a | ||
| D3 | 19.0 ± 1.7b | 11.6 ± 0.6a | 475 ± 20ab | 364.4 ± 13.6a | ||
| D4 | 21.5 ± 0.7a | 12.1 ± 0.6a | 445 ± 18b | 371.8 ± 10.6a | ||
| D5 | 19.0 ± 1.2b | 12.4 ± 0.3a | 409 ± 18b | 375.2 ± 11.7a | ||
| DH618 | D0 | 19.8 ± 0.6b | 12.8 ± 0.2a | 424 ± 7a | 401.3 ± 6.9c | |
| D1 | 20.5 ± 0.6ab | 13.2 ± 0.5a | 356 ± 30bc | 457.3 ± 14.4a | ||
| D2 | 19.7 ± 2.4b | 13.1 ± 1.5a | 355 ± 17bc | 453.9 ± 15.9ab | ||
| D3 | 22.1 ± 0.1a | 13.3 ± 0.9a | 394 ± 18ab | 426.3 ± 16.5bc | ||
| D4 | 20.9 ± 0.7ab | 13.4 ± 0.3a | 357 ± 19bc | 441.8 ± 5.1ab | ||
| D5 | 18.9 ± 0.7b | 13.2 ± 0.3a | 340 ± 32c | 442.1 ± 22.2ab | ||
| 2018 | ZD958 | D0 | 17.6 ± 2.8ab | 9.6 ± 1.3abc | 493 ± 42a | 368.4 ± 18.7a |
| D1 | 18.6 ± 1.7a | 8.1 ± 0.1c | 535 ± 10a | 390.4 ± 31.3a | ||
| D2 | 17.8 ± 2.1a | 9.5 ± 1.0abc | 501 ± 13a | 359.8 ± 12.6a | ||
| D3 | 19.3 ± 1.9a | 8.8 ± 0.5bc | 502 ± 23a | 365.1 ± 12.6a | ||
| D4 | 17.8 ± 1.0a | 10.3 ± 0.7ab | 501 ± 47a | 332.3 ± 1.0a | ||
| D5 | 15.1 ± 0.5b | 10.7 ± 0.6a | 496 ± 32a | 363.0 ± 68.1a | ||
| DH618 | D0 | 19.4 ± 2.1a | 10.7 ± 0.2ab | 453 ± 56a | 405.4 ± 5.0b | |
| D1 | 18.7 ± 1.4a | 11.6 ± 0.7ab | 404 ± 44ab | 413.7 ± 15.2b | ||
| D2 | 17.5 ± 1.3a | 12.8 ± 0.1a | 385 ± 3ab | 400.8 ± 19.4b | ||
| D3 | 18.9 ± 0.9a | 12.4 ± 0.9a | 394 ± 9ab | 401.6 ± 15.1b | ||
| D4 | 19.8 ± 1.2a | 10.3 ± 0.3b | 449 ± 21ab | 403.8 ± 10.2b | ||
| D5 | 17.8 ± 0.9a | 12.7 ± 2.3a | 369 ± 68b | 488.9 ± 60.9a | ||
| Source of variation | ||||||
| Year (Y) | ** | ** | ** | * | ||
| Cultivar (C) | ** | ** | ** | ** | ||
| Leaf removal (L) | ** | ** | ** | * | ||
| Y × C | ns | * | ns | ns | ||
| Y × L | ns | ns | ns | ns | ||
| C × L | ns | * | ** | * | ||
| Year | Cultivar | Treatment | Dry Matter (Mg ha−1) | HI | Source–Sink Ratio (g/kernel) | ||
|---|---|---|---|---|---|---|---|
| Silking | Post-Silking | Maturity | |||||
| 2017 | ZD958 | D0 | 19.8 ± 1.5a | 15.0 ± 2.8cd | 34.7 ± 1.5a | 0.47 ± 0.02b | 0.27 ± 0.01d |
| D1 | 19.2 ± 2.3a | 13.3 ± 2.3d | 32.5 ± 0.0b | 0.53 ± 0.01a | 0.21 ± 0.03e | ||
| D2 | 16.9 ± 1.5a | 17.4 ± 1.5bc | 34.3 ± 0.1a | 0.53 ± 0.01a | 0.32 ± 0.00c | ||
| D3 | 16.9 ± 1.7a | 18.5 ± 1.3b | 35.4 ± 1.2a | 0.56 ± 0.01a | 0.32 ± 0.01c | ||
| D4 | 13.9 ± 0.8b | 21.9 ± 0.9a | 35.8 ± 0.4a | 0.55 ± 0.04a | 0.41 ± 0.02a | ||
| D5 | 13.6 ± 0.6b | 17.9 ± 0.5bc | 31.5 ± 0.5b | 0.56 ± 0.01a | 0.36 ± 0.02b | ||
| DH618 | D0 | 17.9 ± 0.6a | 20.5 ± 0.5bc | 38.3 ± 0.5a | 0.53 ± 0.01a | 0.36 ± 0.01c | |
| D1 | 16.0 ± 0.9b | 19.9 ± 2.7bc | 35.9 ± 3.6a | 0.54 ± 0.06a | 0.41 ± 0.04b | ||
| D2 | 15.6 ± 1.3b | 23.5 ± 1.2a | 39.2 ± 0.2a | 0.54 ± 0.01a | 0.49 ± 0.02a | ||
| D3 | 15.3 ± 0.2b | 22.4 ± 0.5ab | 37.7 ± 0.3a | 0.55 ± 0.01a | 0.42 ± 0.02b | ||
| D4 | 13.4 ± 0.3c | 24.0 ± 1.0a | 37.4 ± 1.2a | 0.52 ± 0.02a | 0.50 ± 0.03a | ||
| D5 | 13.3 ± 0.8c | 18.5 ± 2.1c | 31.8 ± 1.7b | 0.53 ± 0.01a | 0.40 ± 0.04bc | ||
| 2018 | ZD958 | D0 | 14.3 ± 1.1a | 23.8 ± 1.3a | 38.0 ± 2.2a | 0.56 ± 0.00a | 0.40 ± 0.04a |
| D1 | 14.0 ± 1.0a | 20.3 ± 0.7ab | 34.3 ± 5.1ab | 0.56 ± 0.02a | 0.32 ± 0.01bc | ||
| D2 | 13.4 ± 1.0ab | 17.6 ± 2.6bc | 31.0 ± 1.9b | 0.56 ± 0.02a | 0.29 ± 0.01c | ||
| D3 | 12.9 ± 0.9ab | 25.6 ± 3.9a | 38.4 ± 3.1a | 0.59 ± 0.02a | 0.42 ± 0.02a | ||
| D4 | 12.6 ± 0.9ab | 20.7 ± 1.7abc | 33.3 ± 0.9ab | 0.57 ± 0.02a | 0.34 ± 0.03b | ||
| D5 | 12.0 ± 0.9b | 16.6 ± 2.8c | 28.6 ± 2.0b | 0.57 ± 0.04a | 0.28 ± 0.02c | ||
| DH618 | D0 | 15.0 ± 1.1a | 29.6 ± 2.0a | 44.6 ± 2.5ab | 0.54 ± 0.00b | 0.48 ± 0.06ab | |
| D1 | 14.7 ± 1.1a | 25.5 ± 0.9b | 40.2 ± 0.1b | 0.54 ± 0.01b | 0.47 ± 0.05ab | ||
| D2 | 13.9 ± 1.0a | 16.2 ± 2.2d | 30.1 ± 3.5d | 0.51 ± 0.00c | 0.31 ± 0.00c | ||
| D3 | 14.2 ± 1.1a | 25.3 ± 1.7b | 39.5 ± 2.5b | 0.56 ± 0.01ab | 0.48 ± 0.01ab | ||
| D4 | 13.8 ± 1.0a | 32.0 ± 1.2a | 45.8 ± 0.3a | 0.55 ± 0.01b | 0.53 ± 0.02a | ||
| D5 | 13.1 ± 0.9a | 20.7 ± 1.5c | 33.8 ± 1.4c | 0.57 ± 0.00a | 0.42 ± 0.03b | ||
| Year | 2017 | 2018 | |||
|---|---|---|---|---|---|
| Transmission (%) | Bottom | Ear | Bottom | Ear | |
| ZD958 | D0 | 0.6 ± 0.2b | 3.6 ± 0.2b | 1.1 ± 0.4c | 9.4 ± 3.9b |
| D1 | 0.2 ± 0.0b | 8.5 ± 2.6ab | 3.2 ± 0.1b | 16.4 ± 2.3a | |
| D2 | 0.7 ± 0.0b | 8.9 ± 1.9ab | 3.6 ± 1.4b | 21.4 ± 8.0a | |
| D3 | 5.3 ± 1.7a | 9.3 ± 1.4ab | 7.1 ± 2.9b | 13.9 ± 3.7a | |
| D4 | 8.3 ± 0.9a | 13.3 ± 4.2ab | 7.2 ± 3.4b | 16.0 ± 1.0a | |
| D5 | 7.9 ± 2.1a | 14.1 ± 3.8a | 20.0 ± 2.5a | 21.1 ± 1.8a | |
| DH618 | D0 | 0.5 ± 0.2b | 2.7 ± 0.6b | 1.1 ± 0.6b | 14.3 ± 2.5a |
| D1 | 0.6 ± 0.2b | 6.3 ± 1.7ab | 0.8 ± 0.3b | 17.7 ± 4.4a | |
| D2 | 2.5 ± 0.8b | 10.8 ± 1.4a | 0.7 ± 0.3b | 21.3 ± 2.7a | |
| D3 | 1.7 ± 0.3b | 3.6 ± 1.0ab | 6.2 ± 2.2a | 14.8 ± 0.9a | |
| D4 | 2.8 ± 0.9ab | 8.2 ± 1.4ab | 8.4 ± 3.2a | 18.7 ± 3.0a | |
| D5 | 4.7 ± 2.1a | 9.3 ± 3.9ab | 10.8 ± 6.5a | 22.6 ± 8.3a | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Yang, Y.; Liu, W.; Guo, X.; Xue, J.; Xie, R.; Ming, B.; Wang, K.; Hou, P.; Li, S. Leaf Removal Affects Maize Morphology and Grain Yield. Agronomy 2020, 10, 269. https://doi.org/10.3390/agronomy10020269
Liu G, Yang Y, Liu W, Guo X, Xue J, Xie R, Ming B, Wang K, Hou P, Li S. Leaf Removal Affects Maize Morphology and Grain Yield. Agronomy. 2020; 10(2):269. https://doi.org/10.3390/agronomy10020269
Chicago/Turabian StyleLiu, Guangzhou, Yunshan Yang, Wanmao Liu, Xiaoxia Guo, Jun Xue, Ruizhi Xie, Bo Ming, Keru Wang, Peng Hou, and Shaokun Li. 2020. "Leaf Removal Affects Maize Morphology and Grain Yield" Agronomy 10, no. 2: 269. https://doi.org/10.3390/agronomy10020269
APA StyleLiu, G., Yang, Y., Liu, W., Guo, X., Xue, J., Xie, R., Ming, B., Wang, K., Hou, P., & Li, S. (2020). Leaf Removal Affects Maize Morphology and Grain Yield. Agronomy, 10(2), 269. https://doi.org/10.3390/agronomy10020269